直链淀粉聚合度对无籽面包果淀粉三元复合物体外一阶消化动力学的影响
2023-11-07张彦军黄崇杏
李 博,张彦军,赵 媛,黄崇杏,*
(1.广西大学轻工与食品工程学院,广西 南宁 530004;2.中国热带农业科学院香料饮料研究所,海南 万宁 571533)
无籽面包果树(Artocarpus artilis(Parkinson)Fosberg)是重要的桑科木本粮食作物,原产北美和非洲,于1992年引进中国[1]。近年来,我国香料饮料研究所培育出多种具有高直链淀粉的无籽面包果新资源,这些面包果形状为椭圆形,表皮为墨绿色,生果肉中含有近76.7%的淀粉[2]。但成熟果实裂解率高、货架期短,因此将这些面包果转化为淀粉可以最大限度地发挥其价值[3]。本课题组前期对香料饮料研究所1号面包果进行研究,结果表明,相比于伊巴丹和马达加斯加品种,其具有较高的淀粉纯度、凝胶强度、分子均一度、凝胶化温度,较低的回转半径(Rg)、表面粗糙度[2]。研究发现面包果淀粉在改性过程中会产生少量淀粉-脂质-蛋白质三元复合物,从而改善其消化性[3],但对中国高直链无籽面包果淀粉三元复合物的研究目前尚鲜有报道。
Zhang Yanjun等[4]报道淀粉超聚结构中直链淀粉链的葡萄糖单元聚集指数能显著影响淀粉多尺度结构,并引起斜率对数(log of slope,LOS)(多层级线性导数拟合)消化动力学的显著变化[5]。因此,适度调节淀粉直链淀粉的聚合度可以降低与饮食相关疾病的风险[6]。但工业化提取淀粉的纯度较低(<80%),通常在淀粉颗粒表面吸附蛋白质和脂肪,使直链淀粉分子、脂质和蛋白质在热处理后形成三元复合物,从而改变淀粉基食品的质地特性、冻融稳定性、消化性。因此,直链淀粉结构对三元复合体消化机理的影响逐渐成为研究热点[7]。低不饱和度的短链脂肪酸和高等电点的蛋白质(如月桂酸(lauric acid,LA)和β-乳球蛋白(β-lactoglobulin,β-LG)等)有利于淀粉三元复合体的形成[8]。Zhang Yanjun等[4]将不同类型直链淀粉与同一类型支链淀粉按质量比1∶1混合,制备出具有不同直链淀粉聚合度的重组淀粉。由于天然淀粉存在紧密有序的直链淀粉链、支链淀粉的高密度侧链螺旋构象、支链淀粉的高度分支位阻,脂质、蛋白质在复合过程中较难接触直链淀粉螺旋空腔,因此复合效率较低[9]。制备重组淀粉过程中淀粉首先需过度糊化,高聚集态的半晶超分子结构几乎完全被破坏,直链淀粉链析出且多数转变为卷曲态,支链淀粉双螺旋结构解旋,其分支点糖苷键旋角发生不可逆性变大[4]。学者研究发现,收集析出、解离的不同种类直链淀粉和同一类型的支链淀粉,在二者重组过程中,不同来源的直、支链淀粉会进一步减少直链-支链淀粉交联网络的形成,多数天然淀粉中直链淀粉的线性结构在水相中转变为具有疏水空腔的可包合螺旋结构,且相比天然淀粉,重组淀粉的直链淀粉卷曲并伸长导致螺旋中葡萄糖残基每转高度增加,能够容纳更多的配体,重组后的支链淀粉相比天然淀粉分支点旋角增大、支链伸长导致复合配体进入淀粉分子的位阻减小以及复合物形成结晶的时间缩短[7,10]。然而,直链淀粉聚合度对重组淀粉-LA-β-LG复合物消化机制的影响鲜有报道。
因此,本研究从5 种无籽面包果中提取具有最高聚合度的支链淀粉和不同聚合度的直链淀粉并制备重组淀粉,将其与LA和β-LG络合,制备重组淀粉-LA-β-LG三元复合物,研究直链淀粉聚合度对重组淀粉三元复合物LOS模式消化动力学的影响,旨在为淀粉基多组分食品的工业化生产提供新的理论指导。
1 材料与方法
1.1 材料与试剂
5 种无籽面包果(高直链变种香料饮料研究所1号软果品种(GJS)、香料饮料研究所1号嫁接菠萝蜜果树品种(FSS)、石梅山庄品种(SMS)、科摩罗品种(SS)和萨摩亚品种(CS))均选自中国热带农业科学院香料饮料研究所。
β-LG冻干粉(含β-LGA和β-LGB,分析纯,纯度≥90%,提取自牛乳)、LA冻干粉(分析纯,纯度≥98%,提取自天然椰子油)德国Sigma-Aldrich公司;抗性淀粉检测试剂盒(含淀粉葡萄糖苷酶液体、酶活力3300 U/mL,提取自黑曲霉)、葡萄糖氧化酶-过氧化物酶冻干粉(混有4-氨基安替比林,酶活力1000 U/mg,提取自黑曲霉)、α-淀粉酶固体冻干粉(酶活力100000 U/g,提取自猪胰腺)爱尔兰Megazyme有限公司。
1.2 仪器与设备
高效体积排阻色谱-多角度激光光散射检测器-示差折光检测器(high performance size-exclusion chromatography-multi-angle laser light-scattering detector refractive index detector,HPSEC-MALLS-RI)系统美国Waters公司;Scientz-18ND冷冻干燥机 宁波新芝生物科技有限公司;80胶体磨 上海科劳机械厂;快速糊化黏性分析仪(rapid visco analyzer,RVA)瑞典波通仪器公司;UV2700紫外分光光度计 日本岛津公司;LIJXN-26离心机 美国Beckman Coulter公司;Nicolet 6700傅里叶变换中红外光谱仪(Fourier transform infrared spectroscopy,FTIR)美国赛默飞世尔公司;TA25差示扫描量热仪(differential scanning calorimetry,DSC)美国TA公司;D8 X射线衍射仪(X-ray diffractometer,XRD)、NanoSTAR小角X射线散射(small angle X-ray scattering,SAXS)系统 德国布鲁克公司;5100N原子力显微镜(atomic force microscope,AFM)日本日立公司。
1.3 方法
1.3.1 淀粉提取
将面包果去皮,50 ℃干燥40 h,研磨成粉,将其与蒸馏水按质量比1∶3混合,然后在胶体磨中细化10 min,浆液过200 目滤布,滤液离心(3000×g、15 min),收集沉淀物。沉淀物按质量比1∶3加入0.5 mol/L Na2S2O3溶液反应36 h,然后离心(3000×g、15min),收集沉淀物并刮去褐色表皮,用体积分数50%乙醇溶液洗3 次,在50 ℃真空干燥直至水分质量分数小于8%,过200 目筛,得纯淀粉[11]。
1.3.2 直链与支链淀粉分离
将淀粉用蒸馏水配制为质量分数3%的淀粉悬浮液,并于140 ℃油浴1 h制备淀粉糊化液,将该解离液于5000×g离心20 min,收集上清液,然后按体积比1∶1加入无水乙醇静置12 h,并于6000×g离心20 min,收集沉淀,即为提取的直链淀粉。
将上述沉淀物与甲醇完全混合,制备80 g/100 mL溶液,通过离心(6000×g,20 min)收集沉淀得支链淀粉,干燥后保存待用[4]。
1.3.3 聚合度及分子质量测定
将0.05 g/100 mL淀粉溶液(二甲基亚砜为溶剂)进样于HPSEC-MALLS-RI系统,柱温60 ℃,进样速率0.3 mL/min。用ASTRA软件处理数据得重均分子质量(mw)、数均分子质量(mn)、分子分散度(polydispersity index,PI,PI=mw/mn)、Rg、分子密度指数(ρ)。聚合度采用mw除以162计算获得。分子构型指数(vf)通过动态分子质量和动态均方Rg曲线的斜率对数拟合得出[12]。
1.3.4 不同直链淀粉聚合度重组淀粉制备
根据1.3.3 节支链淀粉聚合度和血糖生成指数(glycemic index,GI)结果,GJS样品提取的支链淀粉具有最高的聚合度(389624.79)和最低GI(88.95)。因此,将GJS的支链淀粉分别与GJS、CS、SS、SMS、FSS的直链淀粉分别按质量比1∶1混合,将各混合淀粉均按质量比1∶1添加蒸馏水,并于180 ℃充分糊化20 min,然后于4 ℃凝沉72 h制备重组淀粉,重组淀粉于60 ℃干燥24 h。不同直链淀粉聚合度的重组淀粉分别命名为GJS’、FSS’、SMS’、SS’、CS’[13]。
1.3.5 三元复合物的制备及黏度特性分析
在RVA铝桶中加入2.0 g重组淀粉样品、100 mg LA、200 mgβ-LG、20.0 g蒸馏水,利用0.3 mol/L NaOH溶液调节pH值至8.0,定容使总质量达28.0 g。利用RVA的Stander 1程序进行络合(95 ℃、5.0 min、160 r/min),络合过程如图1所示[5]:首先,样品在RVA内部达到淀粉的成糊温度,淀粉颗粒吸水溶胀,其超聚结构裂解,直链淀粉和支链淀粉析出并产生热凝胶化效应,使黏度逐渐增加至临界胶束浓度(即最大黏度);在最大黏度峰和最小黏度峰的区间内,解离的直链淀粉非极性疏水空腔与蛋白质和脂肪酸通过分子间作用力相互吸引;在最小黏度峰和最终黏度的温度区间内,首先直链淀粉与蛋白质和脂肪酸通过结合位点发生复合形成I型和II型复合物;最后,直链淀粉复合物、未复合的游离直链淀粉分子与支链淀粉分子进行半结晶重组从而形成完整的三元复合物颗粒[9]。利用体积分数50%的乙醇溶液洗涤三元复合物,使用液氮将复合物快速凝沉,于60 ℃干燥至水分质量分数小于8%,将不同直链淀粉聚合度的复合物样品命名为GJS’-β-LG-LA、FSS’-β-LG-LA、SMS’-β-LGLA、SS’-β-LG-LA、CS’-β-LG-LA。
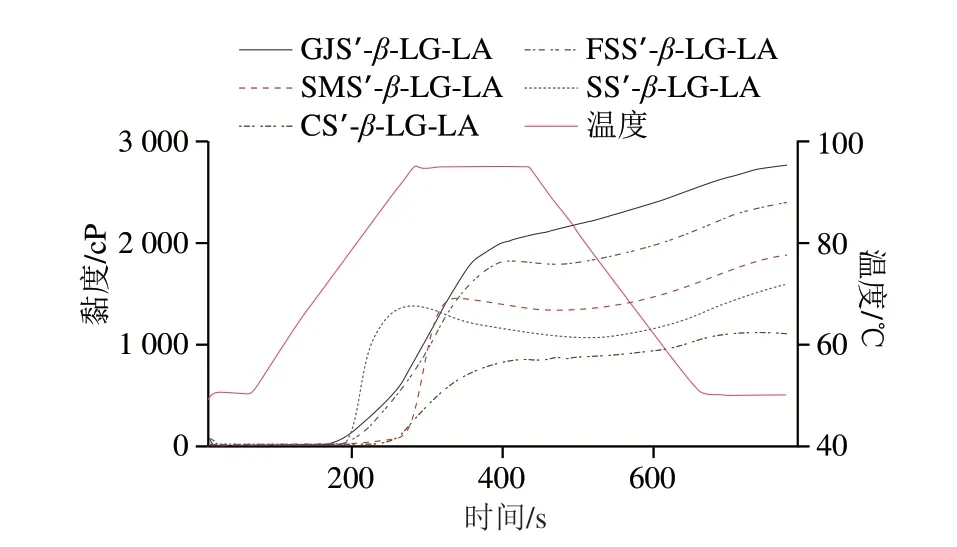
图1 重组淀粉样品的复合过程Fig.1 Changes in viscosity and temperature during complexation of recombinant starch samples
1.3.6 复合指数(complexing index,CI)分析
向100 mg淀粉(干基)中加入1 mL体积分数10%乙醇溶液与9 mL 0.9 mol/L NaOH溶液。将该分散液沸水浴10 min,冷却后稀释至100 mL,取2.5 mL此稀释液继续稀释至50 mL,将其与1 mL I2-KI进行显色反应20 min,利用紫外分光光度计于620 nm波长处测定吸光度,以马铃薯直链淀粉为标准品,按照标准曲线方程y=2.4607x+0.0319(R²=0.9992)计算直链淀粉含量[14]。CI按式(1)计算。
1.3.7 淀粉的LOS模式一级消化动力学分析
根据Zhen Yiyuan等[5]的方法,将200 mg淀粉(干基)和15 mL 0.2 mol/L醋酸钠溶液(pH 5.2)混匀。将10 mL含有290 U/mL胰腺α-淀粉酶和15 U/mL淀粉葡萄糖苷酶的酶混液加入上述淀粉悬液,于37 ℃、150 r/min水浴振荡。在酶解0、10、20、30、40、50、60、90、120、180、360、540 min时取样,收集上清液0.5 mL,加入4.5 mL无水乙醇灭酶,加入葡萄糖氧化酶-过氧化物酶进行显色反应,测定对应时间点葡萄糖浓度。酶解率按式(2)计算。淀粉样品酶解曲线遵循一级动力学方程,如式(3)、(4)所示。
式中:Gt为淀粉被酶解至t时间后转化的葡萄糖质量/mg;C为在t时刻时消化酶酶解淀粉后所产生葡萄糖的浓度/%;Cf∞为最终平衡浓度/%;k为一级动力学速率常数/(10-2min-1)。
将基于上式的LOS消化动力学转化为二阶有限差分方程[5],如式(5)所示。
式中:ki和Ci∞(i=1,2,3,…,f)分别表示对应ti(i=1,2,3,…,f)时间拐点内的多层级动力学速率常数和平衡浓度,f为水解最终阶段。
利用二阶拆分法获取LOS拟合线段[ln[(C3-C1)/(t3-t1)]、ln[(C3-C1)/(t3-t1)]…],对应时间间隔拆分为[(t3+t1)/2,(t4+t2)/2…]。
分别按式(6)、(7)计算曲线下面积(area under curve,AUC)和酶解指数(hydrolysis index,HI)。GI是测试样品基于对比参考食物白面包的餐后血糖应答等级[15],按式(8)计算。
1.3.8 半晶结构表征
利用FTIR的智能ATR附件于4000~400 cm-1范围内进行扫描全波长;对1200~800 cm-1处透光度进行卷积,计算I1047cm-1/I1022cm-1用于分析短程分子有序性。
1.3.9 晶体结构分析,
使用XRD扫描样品得晶体特征曲线,扫描范围为4°~40°,步长2°、扫描速率为4°/min。用JADE 6.5软件计算相对结晶度(relative crystallinity,Rc)。
1.3.10 热力学特性探究
将2 mg样品(干基)与6 μL蒸馏水在坩埚中密封静置12 h,用DSC测定样品的起始温度(To)、峰值温度(Tp)、凝胶化温度(Tc)、凝胶化温度范围(R,R为Tc与To之间的差值)和糊化焓(ΔHg)。仪器参数:温度范围10~120 ℃,加热速率10 ℃/min。
1.3.11 半晶片层结构解析
利用SAXS表征样品片层结构[16],其二维散射光斑数据用于计算0.015<q<0.2 Å-1范围内的一维散射曲线,高斯函数公式如式(9)所示。
式中:r为空间距离/Å;q为散射矢量/Å-1;I(q)为散射强度;d为片层结构厚度/nm。
结合式(9)计算得出强散射峰位置(q1)、半晶片层厚度(d)、非晶相片层厚度(da)和结晶相片层厚度(dc)。
此外,样品分子内部质量分形维数(mass fractal dimension,Dm)可按式(10)计算。
式中:α为分形几何指数,可由I(q)计算得出。
非晶相片层与结晶相片层之间存在可定量的特征间距(ξ),ξ和结晶相片层内微晶单元的平均特征长度(ξc)分别按式(11)、(12)计算。
1.3.12 纳米分形形貌解析
利用AFM轻敲模式扫描2 μm×2 μm样品得纳米微结构均方根粗糙度(Rq)和纳米表面平均粗糙度(Ra)以及三维扫描图[17]。
1.4 数据处理与分析
使用SPSS 22.0软件进行数据分析,采用方差分析进行差异显著性分析。采用Origin Pro 2023软件进行数据拟合、图表绘制和图像处理。
2 结果与分析
2.1 5 种无籽面包果直链淀粉和支链淀粉的聚合度、GI
根据mw除以162计算得出,CS、SS、SMS、FSS、GJS直链淀粉聚合度分别为13450.22、20017.39、25290.01、36017.07、40242.27,CS、SS、SMS、FSS的支链淀粉聚合度分别为125512.65、258646.17、319198.29、339171.66。CS、SS、SMS、FSS的支链淀粉GI分别为91.77、93.85、98.85、99.69。与CS、SS、SMS、FSS样品相比,GJS样品提取的支链淀粉具有最高的聚合度(389624.79)和最低GI(88.95)。因此,将GJS的支链淀粉分别与GJS、CS、SS、SMS、FSS的直链淀粉混合的重组淀粉,用于制备三元复合物。
2.2 三元复合物的一阶消化动力学
图2为三元复合物样品的一阶消化动力学拟合曲线。根据图2计算二阶差分LOS线段,每条线段代表一个消化动力学阶段,如图3所示,每个样品均具有两条斜率与截距完全不同的LOS线,表明每个样品的消化过程中均经历了两个完全不同的酶解动力学阶段,存在两种酶解位点,因此均具有两个C∞与k值,分别对应C1∞、C2∞、k1、k2[18]。根据Kim等[19]的报道,这两种酶解位点可能分别位于无定形区以及结晶区与无定性区之间的缺陷晶层。Zhen Yiyuan等[5]研究大米淀粉三元复合物的LOS动力学得到相似的结果。

图2 三元复合物样品的一阶消化动力学拟合曲线Fig.2 First-order kinetic fitting curves of digestion of complexes
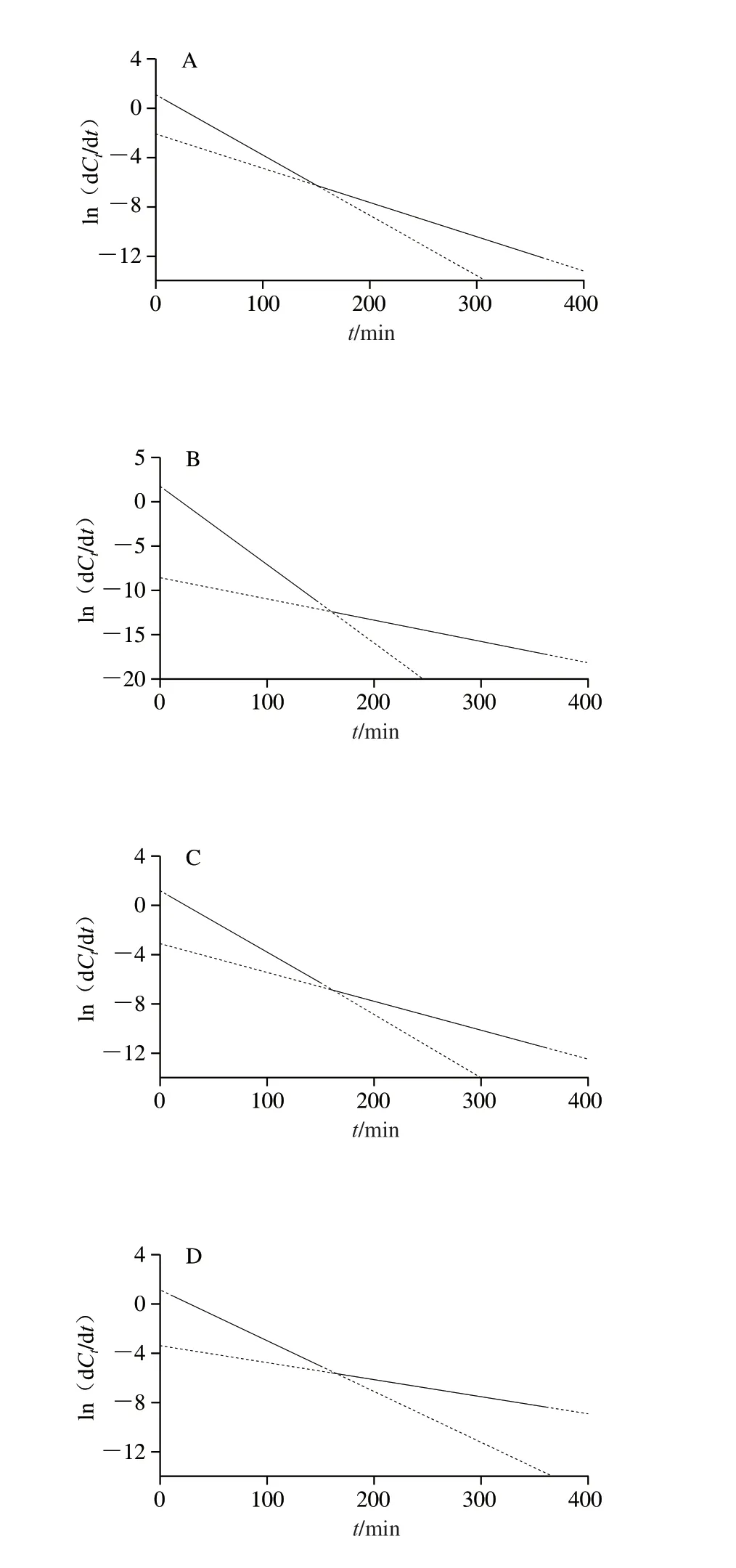
图3 样品的LOS一阶消化动力学模型Fig.3 LOS plots of first-order digestion kinetic models of samples
如表1所示,随着复合物直链淀粉聚合度的增加,样品的C1∞、C2∞分别由69.02%、70.97%降低至60.33%、61.97%;k2从1.00×10-2min-1升高至2.92×10-2min-1,k1则无明显变化规律。根据Kim等[19]的报道,随着直链淀粉聚合度的增加,在复合过程中直链淀粉对游离配体的缔合能力和直链淀粉的重结晶能力逐渐增强,导致抗酶解能力增强,因此复合物直链淀粉聚合度与C1∞、C2∞呈负相关。Rahaman等[20]报道,随着复合物直链淀粉聚合度的增加,缺陷微晶基质内消化酶对相应支链淀粉侧链α-糖苷键的可及性被加强,使淀粉颗粒更迅速地从C1∞酶解至C2∞,这与本研究中复合物直链淀粉聚合度与k2呈正相关的结果一致。此外,三元复合物样品的t1(150 min)和t2(540 min)均相同,且k1(4.02×10-2~8.27×10-2min-1)与直链淀粉聚合度呈弱相关,此结果可能与样品无定形区非线性直链淀粉链的异质性较大有关[21]。
如表1所示,5 种三元复合物的HI为69.73~82.47、GI为78.83~85.71。根据Goñi等[22]的分类,本研究5 种样品均属于高GI食物(GI>70),且随着直链淀粉聚合度的增加,三元复合物的HI和GI降低。这可能归因于复合物直链淀粉聚合度的升高会增加直链-支链分子交联网络的紧密度、表面微结构的致密性,导致形成更强的物理屏障,从而降低了消化酶对底物的可及性,延缓了血糖代谢使C∞、HI和GI降低[20]。Cai Canxin等[15]报道大米淀粉复合物的HI和GI为88.6~93.9和88.4~91.2,Zhen Yiyuan 等[5]报道大米淀粉复合物的k1和k2为6.75×10-2~6.98×10-2min-1和1.68×10-2~1.88×10-2min-1,与本研究结果相似。
2.3 三元复合物的半晶结构特征
FTIR和XRD图谱可以用于评估重组淀粉复合体的半晶结构。如图4所示,5 种复合物样品的在FTIR全谱均在1540 cm-1和2846 cm-1处观察到振动峰,这些峰与脂肪酸亚甲基上的羰基和C—H键的不对称伸缩振动以及蛋白质内部氨基酸酰胺II带的形变振动相关,表明重组淀粉与脂肪酸和蛋白质成功复合。如图5所示,支链淀粉链的支化方式、直链-支链淀粉微晶单元的堆积方式为偏序型,使复合物最终呈现V型结晶反射,均在2θ为13°、20°处存在衍射峰。如表2和图6所示,随着直链淀粉聚合度的增加,1540 cm-1和2846 cm-1处峰强度、CI(68.81%~83.55%)、短程有序性(I1047cm-1/I1022cm-10.44~0.63)、Rc(21.10%~28.35%)均增加。结合三元复合物CI和消化动力学结果分析发现,随着直链淀粉聚合度的升高,三元复合物CI和GI同时增加,这可能是由于三元复合物直链淀粉聚合度的升高会增加线性直链葡聚糖最佳络合位点的数量,使直链淀粉螺旋空腔中的(4→1)-α、(1→4)-α-CH2O桥基结合脂肪酸疏水尾部和蛋白质多肽极性侧链的能力提高,增加了晶核成核速度,促进了更多V型结晶复合物的形成[23]。此现象会进一步导致三元复合物晶体结构中的羟基体积增大、支链淀粉簇的纳米结节致密度增加,因而增加了淀粉精细结构的空间位阻密集度,导致酶分子的结合率降低,从而降低GI[24]。综上所述,随着复合物直链淀粉聚合度增加,样品的CI、颗粒内部有序结构数量增加,导致了C2∞、GI的降低。Kawai等[14]报道马铃薯淀粉复合物的CI为68%~85%;Cai Jingjing等[25]报道了马铃薯、玉米、小麦淀粉复合物的Rc为30.3%~33.2%;Lin Li等[23]发现大米淀粉复合物的短程有序性为0.542~0.544,上述报道与本研究结果相似。
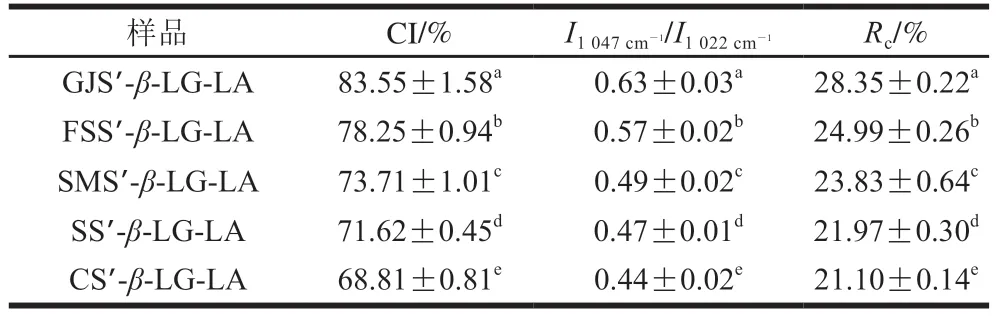
表2 复合物样品的CI、I1047cm-1/I1022cm-1和RcTable 2 CI,I1047cm-1/I1022cm-1 and Rc of complex samples
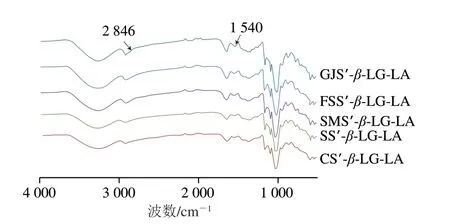
图4 样品短程结构的FTIR图谱Fig.4 FTIR spectra of short-range order structures in starch samples

图5 复合物样品的XRD图谱Fig.5 XRD patterns of complex samples
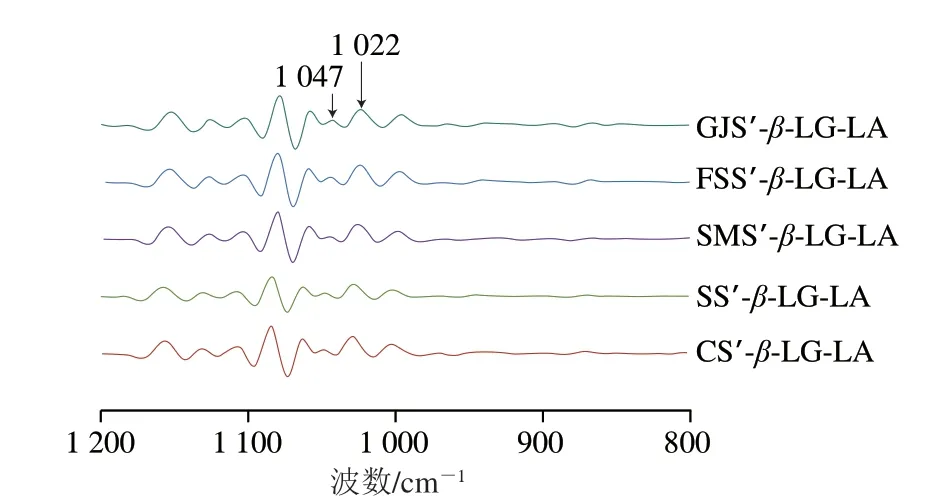
图6 样品短程结构的FTIR去卷积光谱Fig.6 Deconvoluted FTIR spectra of short-range order structures in starch samples
2.4 三元复合物的热力学特性
如图7所示,5 种复合物样品均具有3 个热力学特征峰,峰1为游离脂肪酸熔融峰(39.78~42.05 ℃)、峰2为I型复合物的凝胶化峰(56.68~60.11 ℃)、峰3为IIb型复合物(98.52~103.71 ℃)。图中并未显示IIa型复合物峰,这可能是由于IIa型复合物与未复合的凝沉淀粉有相似的凝胶化温度从而汇聚成一条宽截面曲线[26]。如表3所示,随着复合物直链淀粉聚合度的增加,样品的凝胶化温度范围、Tp、ΔHg分别从19.42 ℃、98.52 ℃、13.95 J/g增加到26.69 ℃、103.71 ℃、18.73 J/g。结果表明,复合物直链淀粉聚合度的增加可以增加晶格单元在无形性介质中交联的有序性、晶胞内V6/7/8型双螺旋(一个螺旋单元有6/7/8 个脱水葡萄糖单元)单元的有序性、晶胞内双螺旋单元的数量[21]。高直链淀粉复合物直链淀粉聚合度的升高可使复合物具有较高的CI、短程有序性和Rc,从而引起更多更有序的晶胞交联结构聚集,导致复合物更难熔融以及被酶解。Cai Jingjing等[25]报道玉米、马铃薯、小麦淀粉复合物的Tp为98.7~100.5 ℃。此外,直链淀粉聚合度与I型复合物的凝胶化温度范围、Tp、ΔHg呈现较弱的相关性。
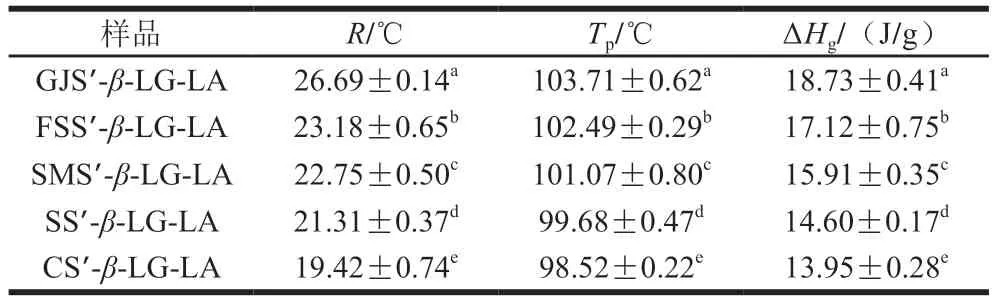
表3 复合物样品的凝胶化温度范围(R)、Tp和ΔHg Table 3 Ranges of gelation temperature (R),Tp and ΔHg of complex samples
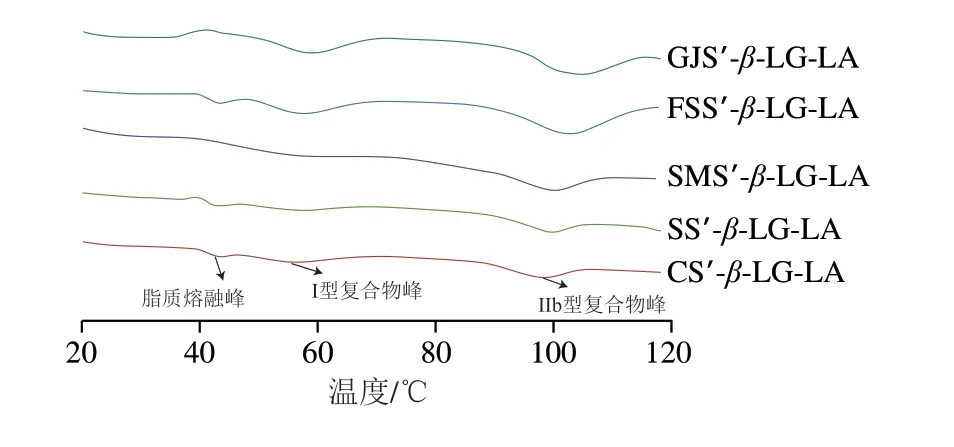
图7 复合物样品的DSC曲线Fig.7 DSC curves of complex samples
2.5 三元复合物的多尺度超分子结构
2.5.1 片层结构特征
淀粉的多尺度超分子结构常用SAXS、动态分子质量以及AFM等手段评估[27]。如图8所示,所有样品中均在约0.5 nm-1处显示出不明显的宽峰,可能与周期长度约13 nm的未复合质量分形结构相关,而样品的强峰q1位于3.71~3.72 nm-1处,表明所有络合物样品都有V 100-B型螺旋间反射[16]。然而,XRD分析并未检测到V+B型峰,这可能是因为V 100-B型螺旋晶胞间距较小、布拉格间距过大导致[28]。2D散射图(图9)表明出复合物样品均具有不均匀的环状近晶相非周期结构,这种分形聚集体又被称为“两相非颗粒体系分子结构”[16],此结果与上述复合物样品具有两种酶解位点互相印证。
图8 三元复合物样品的SAXS图谱Fig.8 SAXS curves of complexes
如表4所示,随着复合物直链淀粉聚合度的增加,片层结构参数d(3.20~4.82 nm)、da(1.60~2.62 nm)、dc(1.60~2.20 n m)、Dm(1.28~1.81)、ξ(4.54~5.53 Å)、ξc(6.33~7.01 Å)均降低。此结果表明复合物直链淀粉聚合度的增加可以加强复合物球晶边缘之间的超共轭效应,进而增加分子间的静电吸附力,使整个半晶片层体系电子云密度和片层结构内容物的均匀性升高、微晶单元致密性增加。此外,Lu Xuanxuan等[16]认为,复合物直链淀粉聚合度的增加不仅会使片层结构更紧致,还会使复合物在熔融或酶解时羟基暴露的体积增大,从而使结合水/酶分子位点的活性降低,导致Tp、R、ΔHg、k2增加和C∞、GI值减小。Zhen Yiyuan等[5]报道大米淀粉三元复合物的da和dc分别为1.84~2.23 nm和1.75~2.02 nm,其与本实验结果相似。

表4 复合物样品半晶片层特征Table 4 Characteristics of lamellar structure of complex samples
2.5.2 分子构型参数
如图10和表5所示,折射率色谱图显示出明显的三元复合物峰(106~107Da),这与FTIR和DSC图谱的结果一致。此外,分子质量曲线导数拟合后发现,分子自旋态达到最终态时样品呈现的有效构象指数(νf)接近于1,表明复合物样品均为蓬松棒状分子构型,说明高摩尔质量的球晶核主要随机分布在两相非颗粒体系分子结构的外部[29],这与复合物直链淀粉聚合度对样品中I型复合物热力学参数以及k1影响较弱相关。
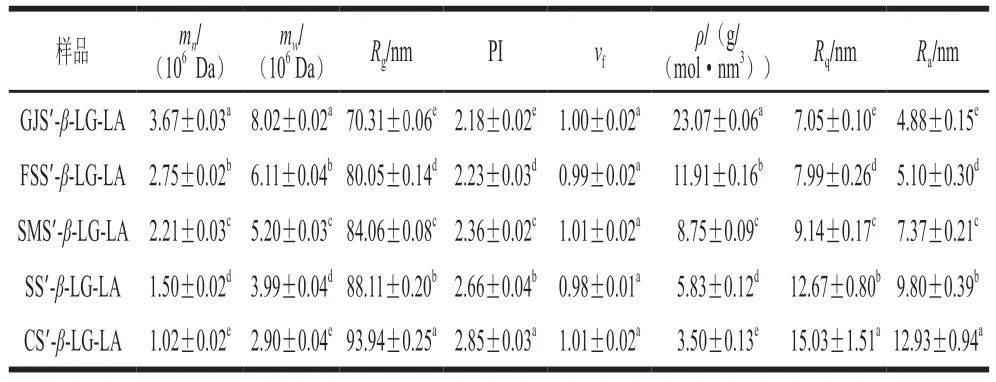
表5 分子构型特征参数与纳米微表层粗糙度Table 5 Characteristic parameters of molecular configuration and nanosurface roughness
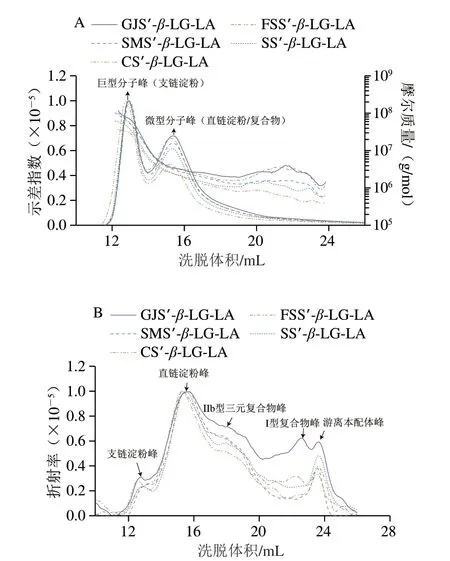
图10 样品的多角度激光光散射信号和分子质量分布(A)以及折射率色谱图(B)Fig.10 Multiangle laser light-scattering signals and molecular mass distribution (A) and chromatograms of refractive index of samples (B)
分子构型参数结果表明,随着复合物直链淀粉聚合度的增加,mn(1.02×106~3.67×106Da)、m w(2 .90 × 106~8 .02 × 106Da)、ρ(3 .50~23 .07 g/(mol·nm3))均增大,Rg(70.31~93.94 nm)、PI(2.18~2.85)则减小(表5)。上述结果可能归因于,在络合过程中随着直链淀粉聚合度的增加,支链淀粉糖苷键构象旋角的角度减小、V8直链淀粉单螺旋包合物含量增加,导致d、Dm、ξ、ξc减少,Rc、CI增加[12]。此外,Wang Shujun等[30]报道,复合物直链淀粉聚合度的增加可以增强支链淀粉游离侧链的空间位阻,增强其抗热解性,从而引起GI值减小。
2.5.3 颗粒表面纳米微结构
Rq表示形成于颗粒表面并连接至淀粉颗粒内部的“止水塞”突起结构尺寸[17]。如图11和表5所示,随着复合物直链淀粉聚合度的增加,样品Rq和Ra分别由15.03 nm和12.93 nm降低至7.05 nm和4.88 nm。基于此推测复合过程中直链淀粉聚合度的升高使直链淀粉链间相邻糖苷氧O6与O2/O3之间形成更多的氢键,导致“止水塞”突起结构尺寸降低[31]。这一变化还可能引起d、ξc、Rg减少、mw、CI、Rc、短程有序性增加,导致颗粒表面空腔数量、粗糙度、分形维数、海绵状结构减少(图11),使Tp增加和C∞降低。Chen Long等[17]报道马铃薯复合物的Rq和Ra分别为4.42~10.30 nm和3.58~8.16 nm,与本研究结果相似。
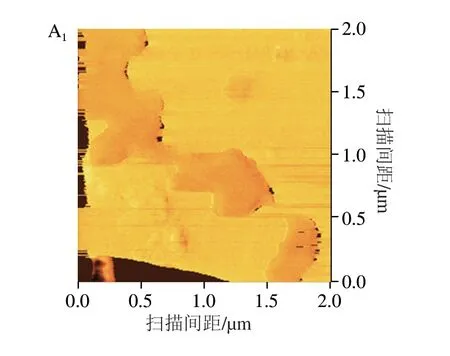
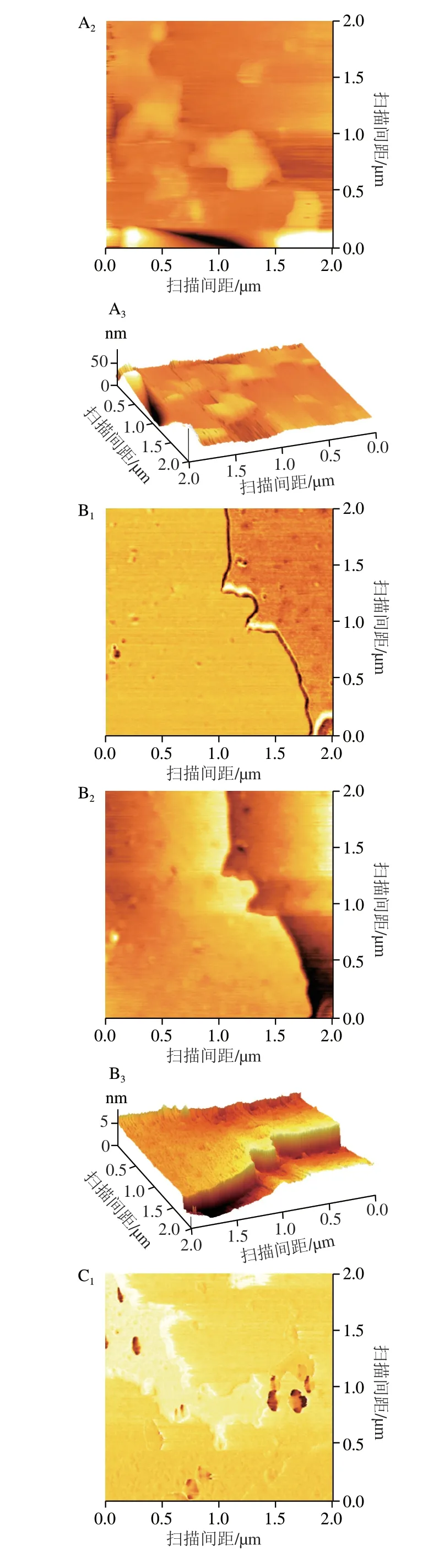
2.6 多元神经网络分析结果
为进一步筛选对体外消化动力学有极显著影响的结构因素(相关性系数(r)大于0.99或小于-0.99,P<0.01),本实验基于邻接矩阵和Fruchterman-Reingold算法进行神经网络分析。如图12所示,神经网络主要包含两个区域,内部区连接点包括C1∞、C2∞、k2、GI、CI、Rc、R、Tp、ΔHg、d、da、Dm、ξ、mw、Rg、PI、Rq,表明这些参数在P<0.01水平上极显著相关。Zhang Yutong等[32]同样报道了变性菠萝蜜淀粉的HI、GI、C∞呈极显著正相关(r>0.99、P<0.01)。而神经网络区由外部散点组成,包括k1、ρ、I1047cm-1/I1022cm-1、ξc、dc,表明这些参数之间在P<0.01水平上无显著相关性,但ρ、I1047cm-1/I1022cm-1、ξc、dc与C1∞、C2∞、k2、GI在P<0.05水平上呈显著相关,本研究发现复合物具有“两相非颗粒体系”以及多消化位点,说明ρ、I1047cm-1/I1022cm-1、ξc、dc在复合物直链淀粉聚合度对消化动力学的影响机制中只对“单相非颗粒体系”以及专一消化位点有显著影响,其他极显著结构因素则对复合物两种酶解位点均具有显著影响。综上所述,复合物直链淀粉聚合度的变化可以极显著地改变复合物消化性。
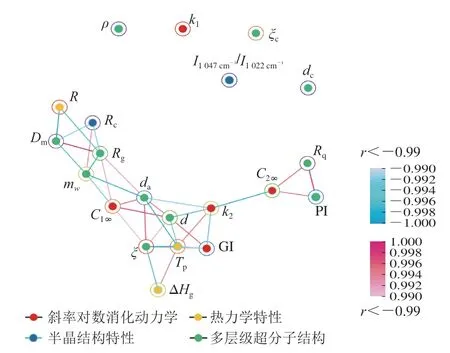
图12 复合物体外消化动力学影响因素的多元神经网路分析图Fig.12 Multivariate neural network analysis of factors influencing in vitro digestion kinetics of ternary complexes
3 结论
本实验探究了直链淀粉聚合度对重组无籽面包果淀粉-β-LG-LA复合物消化性的影响机制。研究发现,复合物样品均呈现V型结晶结构、棒状分子构型,均具有较高的三元复合率。随着直链淀粉聚合度的增加,样品的d、da、Dm、ξ、ξc、dc、Rq减少,CI、I1047cm-1/I1022cm-1、Rc、mw、PI、ρ、R、Tp、ΔHg增加,导致消化动力学参数C1∞、C2∞、GI降低,k2增加。结合神经网络分析发现,复合物颗粒内、外部超显微结构对消化性有极显著影响(r>0.99或r<-0.99、P<0.01)。进一步分析发现,复合物直链淀粉聚合度的增加可以增强在螺旋内、螺旋间、晶胞单元内、纳米片层结构内的分子间、分子内相互作用,导致多层级超结构有序度、致密性增加,使颗粒表层纳米“止水塞”突起尺寸、空腔数量、分形维数显著减少,导致颗粒热溶胀能力下降,从而使LOS消化动力学降低。本研究结论可为针对不同营养需求人群的多组分食品研发提供新的理论参考。
