东阿拉善不同发育阶段沙冬青种群的空间分布格局
2023-11-02刘晓婷王铁娟苏日古嘎李晓田刘雅洁
刘晓婷,王铁娟,苏日古嘎,张 丽,李晓田,刘雅洁
(内蒙古师范大学生命科学与技术学院;内蒙古自治区高等学校生物多样性保护与可持续利用重点实验室,内蒙古 呼和浩特 010022)
种群的空间分布及种内的关联性可以直观反映出种群的动态变化,并有助于理解种群空间结构及群落形成机制[1]。植物种群的空间分布格局即植株在特定的时间和空间中的分布情况,按照植株的分布情况可以分为均匀分布、聚集分布和随机分布[2-3]。均匀分布是由于植株之间对资源的激烈竞争所引起的分布类型;聚集分布是由于植株以母株为中心进行繁殖或资源配置不均匀所引起的;随机分布则是由于资源合理分配之后,个体之间没有明显的生态关系所致[4]。种群空间分布格局对空间尺度具有很强的依赖性,点格局分析[5]法可以在尺度连续增大的过程中,分析不同尺度下的种群格局和种间关系[6],因而被广泛应用于多尺度空间分布格局和不同龄级之间多尺度空间关联的研究中[7],从而在不同尺度上解释种群与环境的关系及生态学过程[8-10]。
沙冬青(Ammopiptanthusmongolicus)为豆科沙冬青属植物,强旱生常绿阔叶灌木。生长在荒漠区沙质和沙砾质地,分布于内蒙古的西部、贺兰山、宁夏北部、甘肃河西走廊北部以及蒙古国南部。沙冬青是古老的第三纪残遗种,为国家二级重点保护植物[11]。该种具有固沙与观赏作用,枝、叶可入药,用于治疗冻疮和慢性风湿关节病[11]。沙冬青属只有2个种,是亚洲中部荒漠区仅有的常绿阔叶灌木,因而可为亚洲中部荒漠植物区系的热带起源学说提供有力的证据[12],具有重要的科学价值。许多学者研究了沙冬青耐性相关的功能基因[13-15]、遗传多样性与分化[16-17]、繁殖特性[18-19]、地理分布[20-21]、生理生态[22-23]以及群落特征[24-25]等。对沙冬青种群分布格局研究也有一些研究报道,多数的研究采用空间指数法[26-28]或以多个指标综合评价[29-31],近些年,申景昕等[9,32]分别运用点格局分析法对不同衰退程度、不同放牧干扰下的沙冬青种群进行了研究。我们在对不同区域沙冬青群落调查过程中发现,地势平坦、沙冬青较大面积分布的区域中,沙冬青个体植株整体较大的群落内几乎不见沙冬青幼株,仅在群落边缘有幼株出现,而在距这样的群落不远的地方会出现以幼株为主稀疏分布的小斑块,周围还有以中等大小植株占优势的沙冬青群落。从种群角度讲,体现了不同发育阶段沙冬青种群的空间异质性。基于此,我们试图通过点格局分析法对处于三个发育阶段的沙冬青种群进行不同空间尺度上的分布格局与空间关联性研究,探讨种群更新的生态学过程及其规律,为沙冬青的保护和进一步的研究提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于巴彦淖尔市的磴口县,属于温带大陆性季风气候,年平均气温7.6℃,年平均降水145 mm,年平均蒸发量2 398 mm,气候干旱,昼夜温差大,降雨量小且多集中在夏季,风沙大。调查地点位于该县西北部的沙金套海苏木,地处草原化荒漠区,海拔为1 064~1 070 m,地势平坦,沙砾质地,生境条件较为一致。选择沙冬青为单优建群种的群落设置样地,样地1地表大面积裸露,沙冬青以小株占据优势,个体分布较为稀疏,为年轻的种群(发育阶段1);样地2中的沙冬青总体以中等大小的植株为主,其他大小的植株也可见,为成熟阶段的种群(发育阶段2);样地3中的沙冬青植株冠幅总体偏大,且大部分植株出现不同程度的枯枝,几乎不见沙冬青幼小植株,为衰退阶段的种群(发育阶段3)。
1.2 调查方法
于2022年在每个样地随机设置了1个50 m×50 m的样方,同时,用测绳、样方钉等工具将每一个样方划分为25个10 m×10 m小样方。以每一个小样方同方向上的顶点选为坐标原点,记录每株沙冬青的空间二维坐标、测量南北向冠幅(a)、东西向冠幅(b)和高度(H),并记录其生活状态。
1.3 大小级结构划分
对于灌木年龄结构的研究,常用大小级来代替年龄级。沙冬青是一种基部多分枝、无明显主干的灌木,因而本研究也采取大小级的划分方法。根据样地内沙冬青的个体大小比例与植株的枯枝率,结合何恒斌等[29]、靳虎甲[30]的研究,采用沙冬青植株体积(即植株高度×冠幅长×冠幅宽)的立方根(d,cm)将沙冬青个体的大小划分为3级,分别为:Ⅰ级,d≤60 cm;Ⅱ级,60 cm
1.4 点格局分析
采用配对相关函数g(r)(Pair correlation function) 对不同发育时期的沙冬青种群以及各大小级进行点格局分析,g(r)函数[33]是在Ripley's单变量K(r)函数[5]的基础上推演得到,是基于圆环内的点与点之间的距离,能够克服K(r)函数存在累积效应的问题,可以较容易的扩展到双变量形式[34]。相关公式为:
(1)
式中:r为空间尺度;A为样方面积;n为个体总数;uij为第i株沙冬青与第j株沙冬青之间的距离;Ir(uij)为指示函数,若uij≤r时,Ir(uij)=1,若uij>r时,Ir(uij)=0;Wij为权重值,用于校正边缘误差。
(2)
当g(r)值位于上包迹线上方时,种群为集群分布,位于下包迹线下方时,为均匀分布,位于上下包迹线之间时,为随机分布。
多元点格局分析是两个龄级(大小级)的点格局分析。本研究采用gab(r)双变量函数进行分析,该函数由Kab(r)函数推导而来。公式如下:
(3)
式中:na、nb表示不同大小级的个体数。
(4)
当gab(r)值落在上下包迹线之间,在r尺度下大小级之间关联不显著;当gab(r)值在上包迹线之上,在r尺度下呈显著正关联;当gab(r)值在下包迹线之下,在r尺度下呈显著负关联。
1.5 零模型选择
点格局分析采用K(r)函数时,易造成在大尺度分析中受到小尺度积累效应的影响。因此,通过采用K函数推导而来的g(r)函数来消除这一效应。但是,g(r)函数存在非聚集性,需要借助零模型来剔除环境异质性的影响[34-35]。本研究通过采用完全随机模型(CSR)和异质泊松模型(HP)分析沙冬青种群及其不同大小级之间的空间分布格局。
运用Origin 9软件做个体空间分布图,利用R 3.6.3软件进行点格局分析。采用199次Monte-Carlo随机模拟计算出由上下两条包迹线围成的95%置信区间。空间尺度r为0~25 m,步长为0.5 m。
2 结果与分析
2.1 沙冬青大小级结构分析
由表1可知,发育阶段1中有66 株植株,盖度为4.0%,密度为0.03株·m-2,个体大小平均值为84.21 cm,Ⅰ级(幼龄级)植株所占比例最大,为45.5%,而Ⅲ级(老龄级)个体很少(占12.1%),Ⅱ级(中龄级)植株也占据较大比例,为增长型种群,处于幼龄向成熟的过渡期,为年轻种群阶段。发育阶段2中有228株沙冬青,盖度为20.3%,密度为0.09株·m-2,个体大小平均值为101.02 cm,其中Ⅱ级植株所占比例最大,为80.7%,Ⅰ级植株数量略多于Ⅲ级植株,为稳定型种群,处于种群发展的成熟阶段。发育阶段3中共有沙冬青96株,盖度为19.3%,密度为0.04 株·m-2,个体大小均值较大,为142.45 cm,该样地中仅有3株Ⅰ级植株,Ⅲ级植株所占比例最大,为51.0%,为下降型种群,且大部分植株出现不同程度的枯枝,处于衰退阶段。
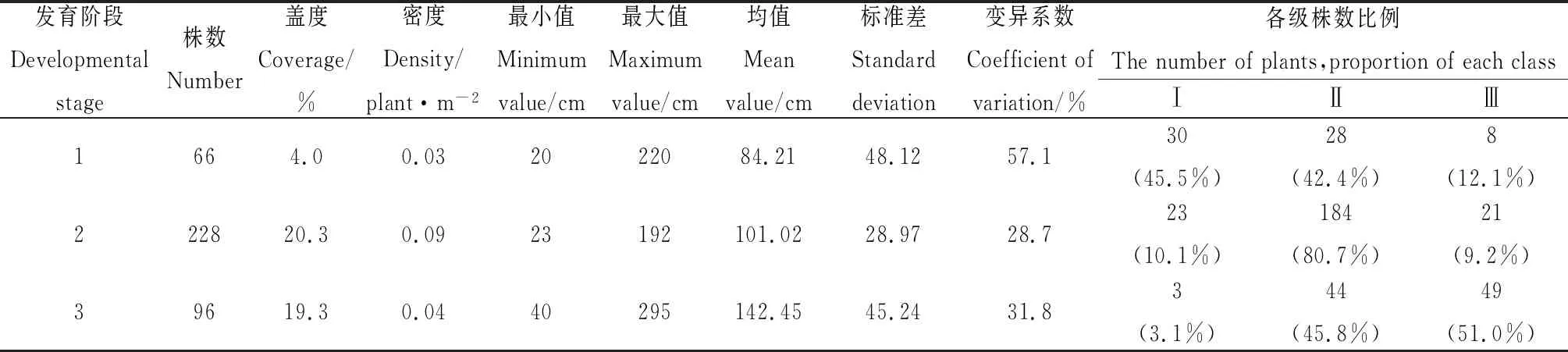
表1 沙冬青种群植株个体大小特征Table 1 Individual size characteristics of A. mongolica populations
2.2 沙冬青种群空间分布格局
由图1可知幼龄级为主的增长型种群(阶段1)个体数较少,中龄级为主的稳定型种群(阶段2)个体数多,且以Ⅱ级占明显优势,高龄级占优势的下降型种群(阶段3)中个体数减少,以Ⅲ级占优势。
采用两种零模型分析沙冬青不同发育阶段种群中个体的空间分布格局,结果略有差异。CSR模型结果显示(图2),阶段1基本呈现随机分布的特点。阶段2在极小尺度下(0~0.6 m)为均匀分布,其余尺度基本表现为随机分布;阶段3与稳定型种群类似,在0~0.6 m尺度下呈现均匀分布,而在3.6~4.7 m,9.1~10.3 m,19.5~20.5 m尺度下为聚集分布,其余尺度呈现随机分布。通过HP模型排除生境异质性的影响后(图2),阶段1种群仅在个别尺度上表现为略高于上包迹线,稍显聚集性;阶段2种群与CSR结果相比,在小尺度(1~5 m)的聚集性增强,中尺度的个别点也呈现聚集分布;阶段3种群与CSR结果基本保持一致。可见环境异质性对沙冬青种群的空间分布影响较小。
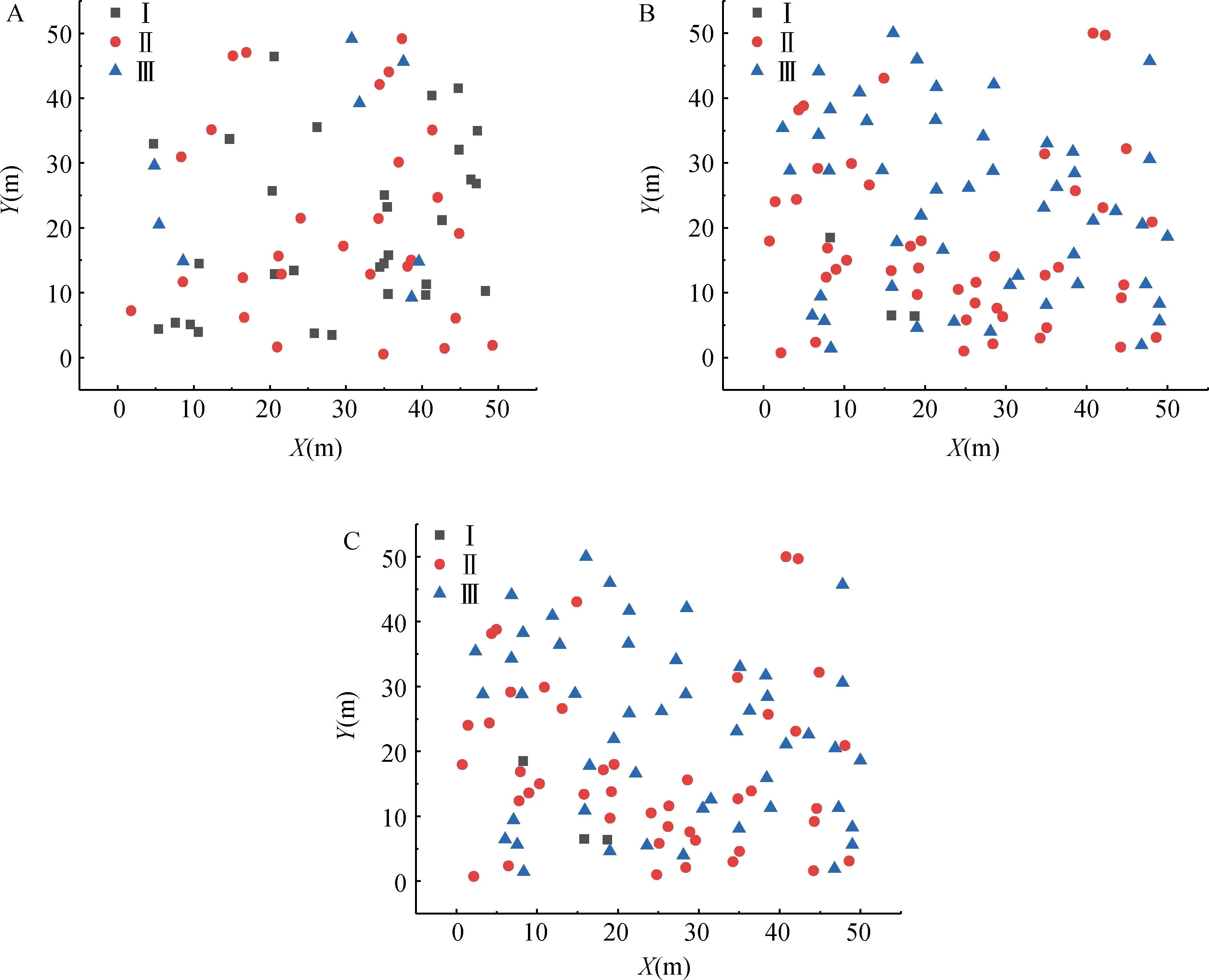
图1 沙冬青种群不同大小级个体的空间分布Fig.1 Spatial distribution pattern of A. mongolicus individuals in different size classes注:A,B,C分别表示发育阶段1,2,3;Ⅰ,d≤60 cm;Ⅱ,60 cm
如图3所示,因个体数少于10株的大小级做分布格局误差较大,因而阶段1仅显示Ⅰ级、Ⅱ级植株的结果,阶段3仅显示Ⅱ级、Ⅲ级植株的结果。CSR模型的结果显示,阶段1种群中Ⅰ级植株在小尺度0.3~1.8 m呈聚集分布,其余尺度为随机分布;Ⅱ级植株在较小尺度的小范围(2.9~4.2 m尺度下)呈均匀分布,其他尺度以随机分布为主。阶段2种群中Ⅰ级植株也以随机分布为主,仅在个别小范围呈现聚集分布(5.7~6.3 m)和均匀分布(11.5~11.9 m和21.0 ~12.5 m);Ⅱ级植株在极小尺度(0~0.6 m)为均匀分布,其他尺度以随机分布为主,但在多个小范围呈聚集分布,其中在1.1~3.8 m聚集性明显;Ⅲ级植株以随机分布为主,在大尺度中的个别小范围呈现均匀分布(17.9~18.6 m和22.8~24.2 m)。阶段3种群中Ⅱ级植株的分布格局在3.8~5.0 m和9.0~12.5 m为聚集分布,其他尺度为随机分布;Ⅲ级植株在小尺度下(0~2.2 m)为均匀分布,其余绝大部分尺度为随机分布。HP模型的结果显示,阶段1种群中Ⅰ级植株与CSR的结果基本一致;Ⅱ级植株的曲线波动性高于CSR的结果,其中在5.0~7.5 m和23.8~24.6 m尺度下聚集性较明显。阶段2种群中Ⅰ级、Ⅱ级、与CSR结果基本一致,Ⅲ级植株在小尺度范围内出现了均匀分布。阶段3种群中,与CSR结果相比,Ⅱ级植株在小尺度上0.8~2.1 m尺度上出现了聚集分布,其余尺度基本一致;Ⅲ级植株在小尺度上均匀分布(0~2.5 m)的范围加宽,中、大尺度的个别点(9.1~10 m和19.6~20.3 m尺度)聚集性增强。
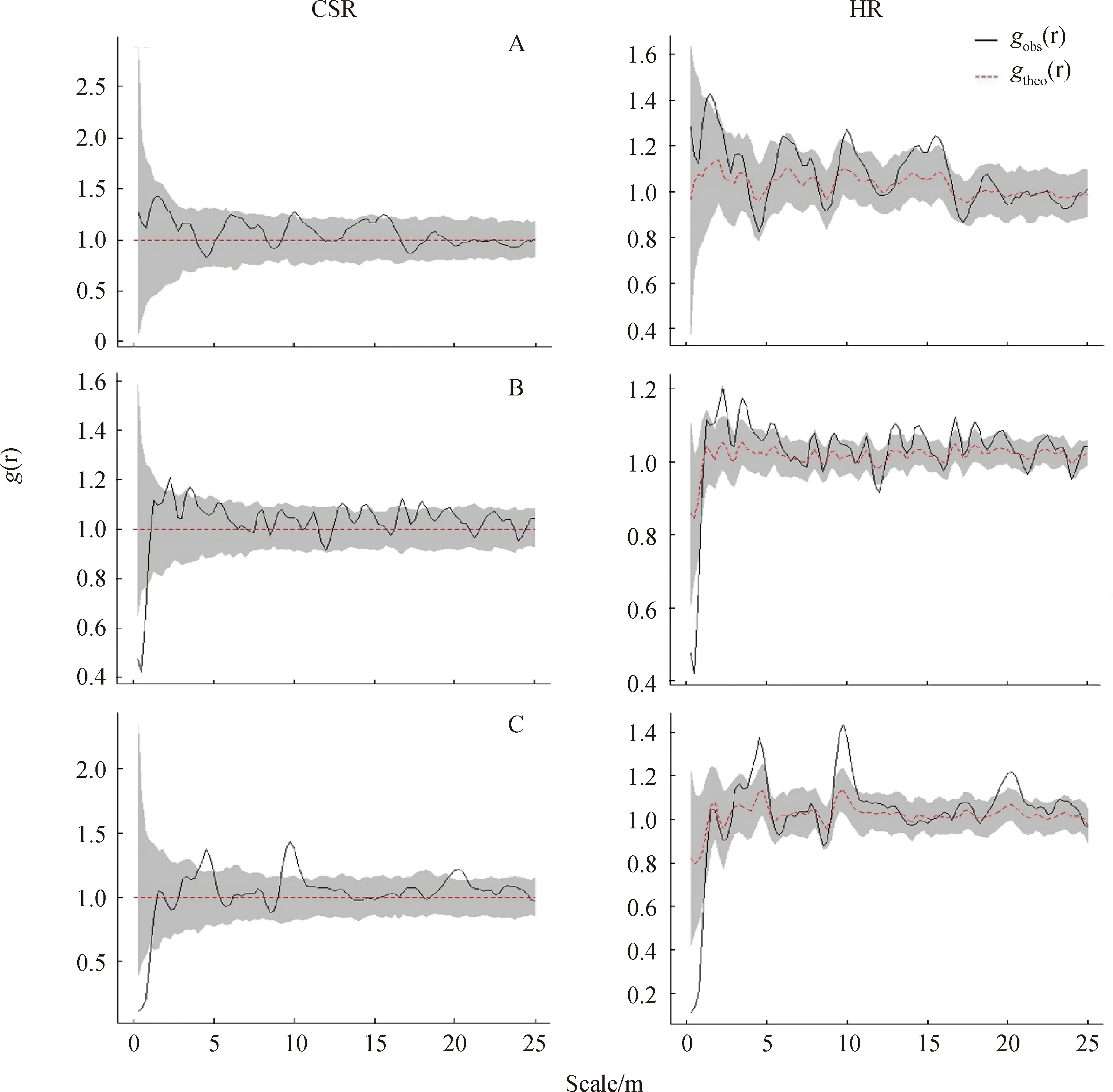
图2 沙冬青种群在完全随机模型(CSR)和异质泊松模型(HP)零模型下的空间分布点格局Fig.2 Spatial distribution pattern of A. mongolicus populations under complete spatial randomness (CSR,left panels) and heterogeneous Poisson (HP,right panels) models注:(r):g(r)的理论值;gobs(r) :g(r)的观测值;灰色阴影部分为95%的置信区间。下同Note:(r) is theoretical value of g(r);gobs(r) observed value of g(r);Gray shaded part was the 95% confidence interval.The same as below
2.3 沙冬青各大小级间关联性
三个发育阶段沙冬青种群不同大小级间的关联性见图4,各大小级间以无显著关联性为主,个别尺度具有一定的关联性,表现在:Ⅰ级与Ⅱ级植株在发育阶段1的中尺度(8.9~11.2 m)、发育阶段2的小尺度(0.4~3.5 m)表现出正关联;III级植株与另2个级别的植株具有一定负关联,这种负关联性表现在小尺度上,其中阶段2中Ⅲ级与Ⅰ级植株在0.7~2.7 m尺度、Ⅲ级与Ⅱ级植株在小于0.4 m为负关联,在阶段3中,Ⅲ级与Ⅱ级植株在0.4~1.5 m和2.3~3.1 m尺度内表现为负关联,不过在大尺度19.2~20.2 m和22.0~25.0 m尺度内表现为正关联。
3 讨论
3.1 沙冬青种群的年龄结构
种群年龄结构是指不同生长发育阶段内的个体在种群内的占比情况,可以反映种群的动态、发展趋势以及与环境的关系[36]。就沙冬青而言,在2010年及之前的大多数研究显示其种群呈下降型或稳定型[26,28-30,37],且种群中缺乏幼苗,更新困难。因沙冬青仅以种子进行繁殖,如果种群不能补充幼体,那么近些年沙冬青种群是否会因衰退而大量减少呢?我们于2021—2022年对内蒙古阿拉善盟、巴彦淖尔市、乌海市和鄂尔多斯市不同沙冬青群落的调查显示,沙冬青年龄结构以稳定型为主,增长型和下降型也可见到。在调查过程中我们发现,沙冬青呈较大面积分布的区域,相邻地点存在着不同龄级占优势的类型。以中龄级(Ⅱ级)占优势的种群中,幼龄级(Ⅰ级)会占据着一定的比例(如发育阶段2),而处于衰退期的沙冬青种群即使中龄级占据着一定的比例(如发育阶段3),幼龄级也极为少见。这可能是由于种群发展到后期,成熟个体间对水分、养分等资源的竞争激烈,对弱小竞争者愈加不利。以幼龄级为主的增长型种群中沙冬青盖度很小,大部分地面裸露,反映出是在空白地建立的年轻种群。每个物种都试图拓展其分布空间,逃避假说(Janzen-Cannell假说)认为种子离开母体的扩散且占据营养更丰富的生境,有助于种子萌发、幼苗存活[38-39],况且通过种子传播占据更广阔的生境也可以减少未来代际间的竞争,沙冬青正是通过种子扩散占据新生境,保持着种的延续。然而,即使是以幼龄级为主的沙冬青种群(增长型种群)中仍然缺乏实生苗,根据刘果厚[40]的研究,作为热带起源的沙冬青,种子适宜萌发温度为30℃左右,6月份地面温度较适,但缺乏降水,7-8月份降水增加,但地面温度过高,加之沙冬青种子的虫蛀率高以及动物的取食,造成了沙冬青幼苗极少。根据赵新艳[37]对沙冬青繁殖的生物气候响应实验,在 6月萌发的沙冬青生长较好,且能越冬。因而沙冬青的更新主要受水分的影响,与其他荒漠植物一样,水分是主要的限制因子,而荒漠区降水波动性很大,沙冬青增长型种群的存在,说明沙冬青遇雨水较好的年份(尤其是6月份),便会有幼苗的补充,只是为间歇性的。
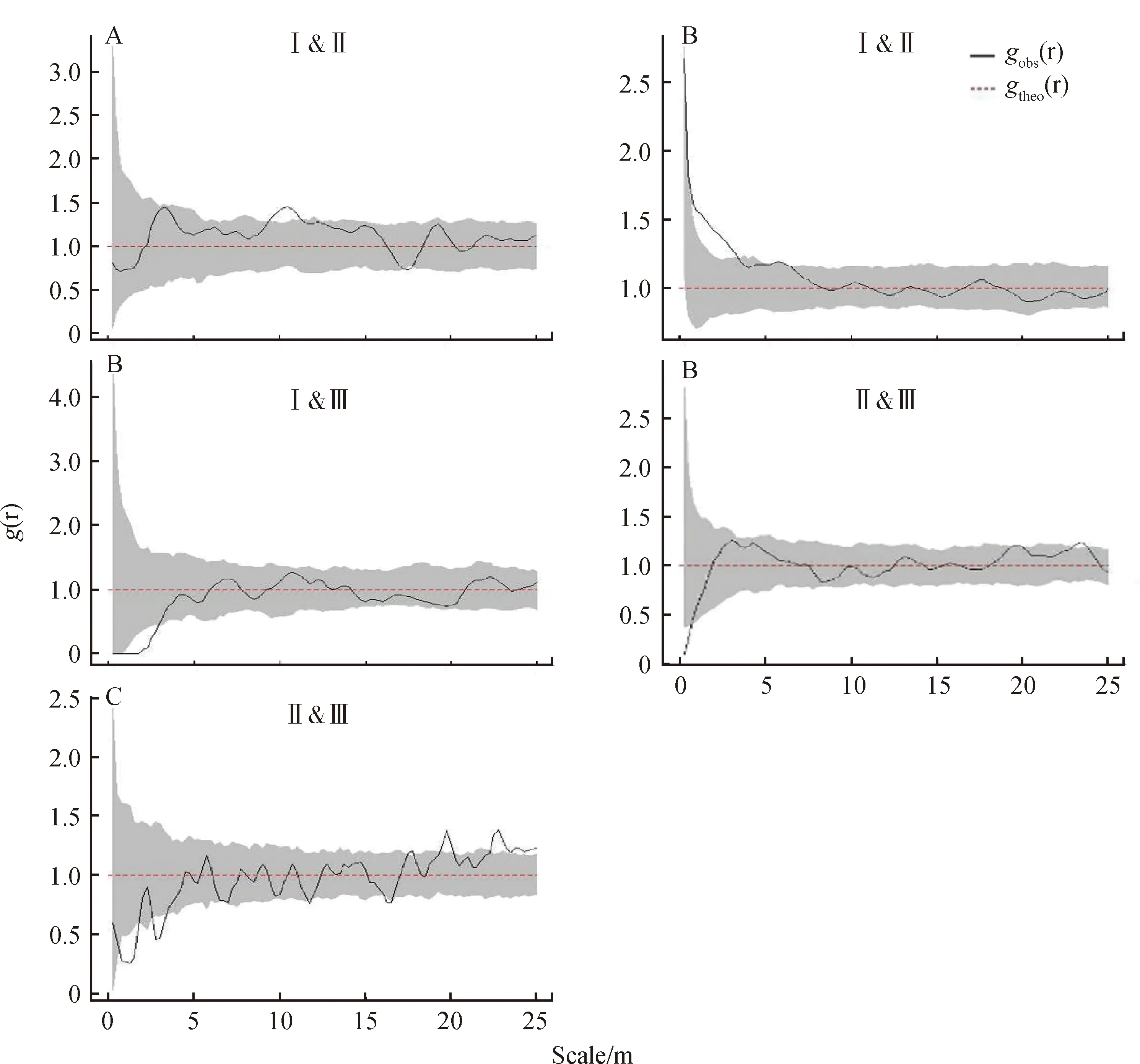
图4 沙冬青种群不同大小级间的空间关联性Fig.4 Spatial correlation between different size classes of A. mongolicus populations
3.2 沙冬青种群空间分布格局分析
植物种群的空间分布格局是种子扩散机制、密度依赖、环境异质性等生物与非生物因素共同作用的结果[9,41]。从沙冬青三个发育阶段总的分布格局来看(图2),从年轻的增长型种群到衰退的下降型种群,个别尺度的聚集性有所增强,但仅限于很小范围内,主要呈现随机分布。不过成熟阶段和衰退阶段的种群,在小尺度下呈现均匀分布,反映出这两个阶段的种群个体间的竞争加大。
由三个阶段各大小级的分布格局来看(图3),幼龄级个体(Ⅰ级)在年轻的增长型种群中(阶段1)的小尺度上呈现聚集分布,在稳定型种群(阶段2)中的中小尺度存在聚集分布,其他尺度基本为随机分布。一般来讲,小尺度的空间分布格局主要受自身生物学特性影响,尺度较大时,主要受种间的相互作用和环境因素的作用[8,42]。根据张荣[43]对沙冬青果实形态与扩散特征的研究结果,沙冬青的种子随果荚在风的作用下脱离母株,果荚具有3种形态,其中开裂—扭曲状的果荚与地面的摩擦面积小,传播距离最远,这种果实也是沙冬青在进化过程中形成适应风力传播的生态策略,并且预测扭曲状果实潜在最大扩散面积为3 017 m2。在一个地面裸露且较为匀质的新生境中,沙冬青荚果随风传播而至,种子从荚果中掉落具有一定的随机性,并且在大风的作用下会发出多次的随风扩散,加之可利用的资源较多,因而Ⅰ级植株呈现以随机分布为主的特点。不过在小尺度上存在聚集性,反映了幼小个体通过聚集生活和彼此庇护来提高生存机率[44]。种群内的中龄级(Ⅱ级)植株的分布格局状况是在幼龄级的基础上发展起来,对于研究区的沙冬青种群,在年轻型种群的基础上,随着种群的发育,植株长大,种群中伴随着种子的萌发不断补充幼龄个体,并且世代不断叠加,所以使分布格局变得复杂。Ⅱ级植株虽然也以随机分布为主,但在增长型种群和稳定型种群中的小尺度上出现均匀分布,反映了邻近个体间的竞争;在三个发育阶段的中或中小尺度上存在聚集分布,尤其是在稳定型种群中更为明显,与成熟植株对环境的适应能力增强,数量增加有关。与Ⅱ级植株相比,老龄级(Ⅲ级)植株的随机分布范围又变宽,在大部分尺度上表现为随机分布,其中在下降型种群中的Ⅲ级植株在小尺度上呈均匀分布,且强度大于稳定型种群的Ⅲ级植株,反映了从成熟期到衰退期高龄级个体间竞争作用加大。沙冬青分布于荒漠区,环境条件恶劣,资源有限,伴随着对水分等条件的竞争,处于弱势的个体被逐步淘汰,个体间的关系减弱,因而Ⅲ级植株的分布格局在更多的尺度范围呈现随机分布。何恒斌等[29]对磴口县三种生境下沙冬青种群分布格局的研究显示:幼龄和老龄植株均成随机分布,中龄植株呈聚集分布,其所研究的尺度范围内(仅做了5~20 m)呈现的规律与本研究中稳定型(发育阶段2)和下降型(发育阶段3)种群具有一致性。本研究中基于完全空间随机零模型(CSR)与基于异质泊松零模型(HP)的结果相比,后者的变化更为明显一些,但总体具有较高的一致性,与研究区生境状况相似,异质性较低有关。
3.3 沙冬青种群各大小级间的关联性
处于不同发育阶段的沙冬青种群,各大小级之间的关系有着一定的差异。在年轻的种群(增长型种群,阶段1)中,Ⅰ级(幼龄级)与Ⅱ级(中龄级)植株在中尺度上小范围内呈现显著正关联,在发育成熟的种群(稳定型种群,阶段2)中,两者在小尺度呈现正关联,表现出幼小植株获取资源的能力较低,对成年植株具有一定依赖性。对于稳定型沙冬青种群,大小级结构完整,Ⅱ级植株占明显优势,Ⅰ级和Ⅲ级也占有一定比例,其中Ⅲ级与Ⅰ级、Ⅱ级植株在小尺度呈现显著负关联,说明处于老龄的Ⅲ级个体对临近的另两个级的个体具有一定的竞争排斥效应。而到了衰退阶段的下降型种群中,幼龄(Ⅰ级)个体只占3.1%,大部分个体已进入到中、老龄,Ⅱ级与Ⅲ级植株在小尺度上仍反映出负关联,进一步反映出两个龄级间的竞争关系。不过,Ⅱ级与Ⅲ级在大尺度上出现一定的正关联,反映了成熟植株间利用资源的相似性,不同龄级之间相互协作,可增加种间竞争力,使种群在群落中处于优势地位[8,45]。不同发育阶段的沙冬青大小级间在大部分尺度范围内呈现无关联性,与年轻种群阶段资源较丰富且生境较一致,降低龄级间的相互作用,成熟与衰退种群阶段自疏效应导致个体间关联性降低有关。
4 结论
研究区沙冬青大面积分布,相邻地点存在不同发育阶段的种群,在空间上具有一定的关联性和异质性。处于老龄阶段的种群幼龄个体数量严重不足,呈现衰退趋势,年轻的增长型种群存在于有较大剩余空间的环境中。不同大小级的沙冬青植株随尺度变化分布格局具有一定的差异性:幼龄个体(Ⅰ级)以随机分布为主,小尺度呈现聚集分布;中龄个体(Ⅱ级)多在小尺度呈现均匀分布,中、大尺度出现聚集分布,且以稳定型种群最为明显,下降型种群次之;老龄个体(Ⅲ级)又以随机分布为主,在小尺度出现均匀分布,且在下降型种群中更为明显。这些变化特征与沙冬青种子传播方式、成熟个体间竞争以及对环境的适应能力密切相关。从年轻种群到成熟种群阶段,沙冬青Ⅰ级与Ⅱ级植株在中或小尺度具有一定的正关联性,反映出幼龄对成熟个体的依赖性。成熟种群到衰退种群阶段,Ⅱ级与Ⅲ级植株在大尺度上存在一定正关联,表现出成熟植株利用资源的相似性;Ⅲ级植株在小尺度上与另两个龄级存在负关联,表现出对相邻的两个龄级个体具有一定的竞争排斥效应。本研究基于沙冬青大面积分布的区域,对于小片分布的地点,其种群发展动态如何应定期监测,及时了解其生存状态,采取针对性的保护措施,其中保证沙冬青的拓展空间、提高幼龄个体存活率,对保持种群的更新尤显重要。
