KLHL7基因在胃癌中的表达及与胃癌临床病理特征的关系分析*
2023-11-01高夏青杨春婷王梓年王宏伟李海龙
安 霞,高夏青,杨春婷,王梓年,王宏伟,李海龙△
1.甘肃中医药大学附属医院检验科,甘肃兰州 730000;2.甘肃中医药大学第一临床医学院,甘肃兰州 730000;3.兰州市第一人民医院肿瘤科,甘肃兰州 730000
胃癌是全球高发的恶性肿瘤之一,也是全球常见癌症相关死亡原因排第二位的恶性肿瘤。目前胃癌的治疗手段主要是手术切除,但绝大多数胃癌在确诊时已属于进展期,已失去通过手术根治的机会。因此,研究早期诊断和预后标志物意义重大。KLHL7是Kelch样家族蛋白成员之一,主要编码BTB-Kelch相关蛋白,所编码的蛋白质可能参与蛋白质降解。有研究发现,KLHL7基因高表达于肺癌、卵巢癌和结直肠癌等肿瘤中[1],其异常表达参与了部分肿瘤的恶性表型形成,而关于该基因在胃癌中的表达及临床意义目前鲜见报道。本文拟借助数据库对KLHL7在胃癌中组织中的表达情况、预后参数及其相关信号通路等进行生物信息学分析。
1 资料与方法
1.1KLHL7基因在人胃癌组织及癌旁组织中的表达 选择在线仙桃学术数据库(https://www.xiantao.love/)检索分析KLHL7基因在人胃癌组织及癌旁组织中的差异表达情况。仙桃学术数据库所用数据是经癌症基因组学数据分析平台UCSC XENA (https://xenabrowser.net/datapages/)统一处理的肿瘤基因组图谱(TCGA)和基因型-组织表达数据库(GTEx)的TPM(transcripts per million reads)格式的高通量测序数据。本研究所分析的数据均提取自TCGA中的胃癌组织和GTEx中对应的癌旁组织或非配对正常组织数据。所有RNA测序(RNA-seq)数据均以TPM格式表示,并进行log2转化后进行样本间的KLHL7的差异表达比较。非配对组织样本数据选择仙桃学术数据库中来自TCGA数据库414例胃癌样本,210例非配对胃黏膜组织;胃癌表达数据来源于经UCSC XENA统一处理的TCGA和GTEx的FPKM(fragments per kilobase per million reads)格式的高通量测序数据。非配对正常胃黏膜组织提取自TCGA和GTEx中胃癌组织对应的正常组织数据。配对组织样本数据选择仙桃学术数据库中来自TCGA数据库的27例胃癌样本和癌旁组织样本,于TCGA数据库 (https://portal.gdc.cancer.gov/)下载并整理TCGA-胃腺癌(TCGA-STAD)项目STAR流程的RNA-seq数据,并提取TPM格式的数据。
1.2KLHL7基因在人胃癌组织异常表达的临床病理特征分析 利用仙桃学术数据库分析KLHL7基因在配对胃癌组织与癌旁组织中的表达情况、KLHL7基因异常表达的胃癌组织的TNM分期、组织学类型,并对KLHL7的异常表达开展免疫细胞浸润特征分析和受试者工作特征(ROC)曲线分析。
1.3与KLHL7基因具有相关性的共表达基因进行京都基因和基因组百科全书(KEGG)信号通路富集分析 使用cBioPortal在线软件 (https://www.cbioportal.org/)[2]检索基于TCGA数据库的412例胃癌与癌旁组织样本的数据集,检索其“co-expression”模块中与KLHL7基因具有相关性的共表达基因。剔除相关系数<0.30的基因,并分别将剩余的上调和下调的共表达基因全部输入Sanger学术平台(http://vip.sangerbox.com/index.html)[3],运用KEGG一键化富集分析工具进行KEGG信号通路富集分析。
1.4相关共表达基因的蛋白互作(PPI)网络分析和核心基因筛选 选择在线STRING(https://string-db.org/)数据库[4],将有93个与KLHL7具有相关性的肿瘤基因输入在线软件STRING进行PPI网络分析,采用K-means聚类算法,以n=3进行聚类分析。观察并分析节点后,将节点(node)和周边(edge)数据提取后导出文件。将PPI数据库构建作用靶点输入Cytoscape 3.9 软件构建网络,采用其插件CytoNCA计算参数,筛选关键核心基因。进一步使用在线仙桃学术数据库验证核心基因在胃癌与癌旁组织间的差异表达。
1.5统计学处理 如果组间数据满足正态分布和方差齐性,则选用t检验;如果满足正态分布,而不满足方差齐性,则选用Welcht′检验;如果组间数据不满足正态分布,则选用非参数检验Wilcoxon秩和检验,信号通路富集分析使用R软件包cluster Profiler(version 3.14.3)进行富集分析,KEGG信号通路富集分析中,选择Benjamini-Hochberg法错误发现率(FDR)<0.1及P<0.05的信号通路为筛选标准。以P<0.05为差异有统计学意义。
2 结 果
2.1KLHL7基因在人胃癌组织及癌旁组织中的表达 在线仙桃学术数据库检索KLHL7基因,分析结果表明,相对于癌旁组织,KLHL7基因在人胃癌组织中表达上调(P<0.001),见图1;在非配对组织中,KLHL7基因在人胃癌组织的表达也高于正常组织(P<0.05),见图2。其中,图中纵坐标是FPKM格式的高通量测序数据经对数转换后显示的基因表达量。
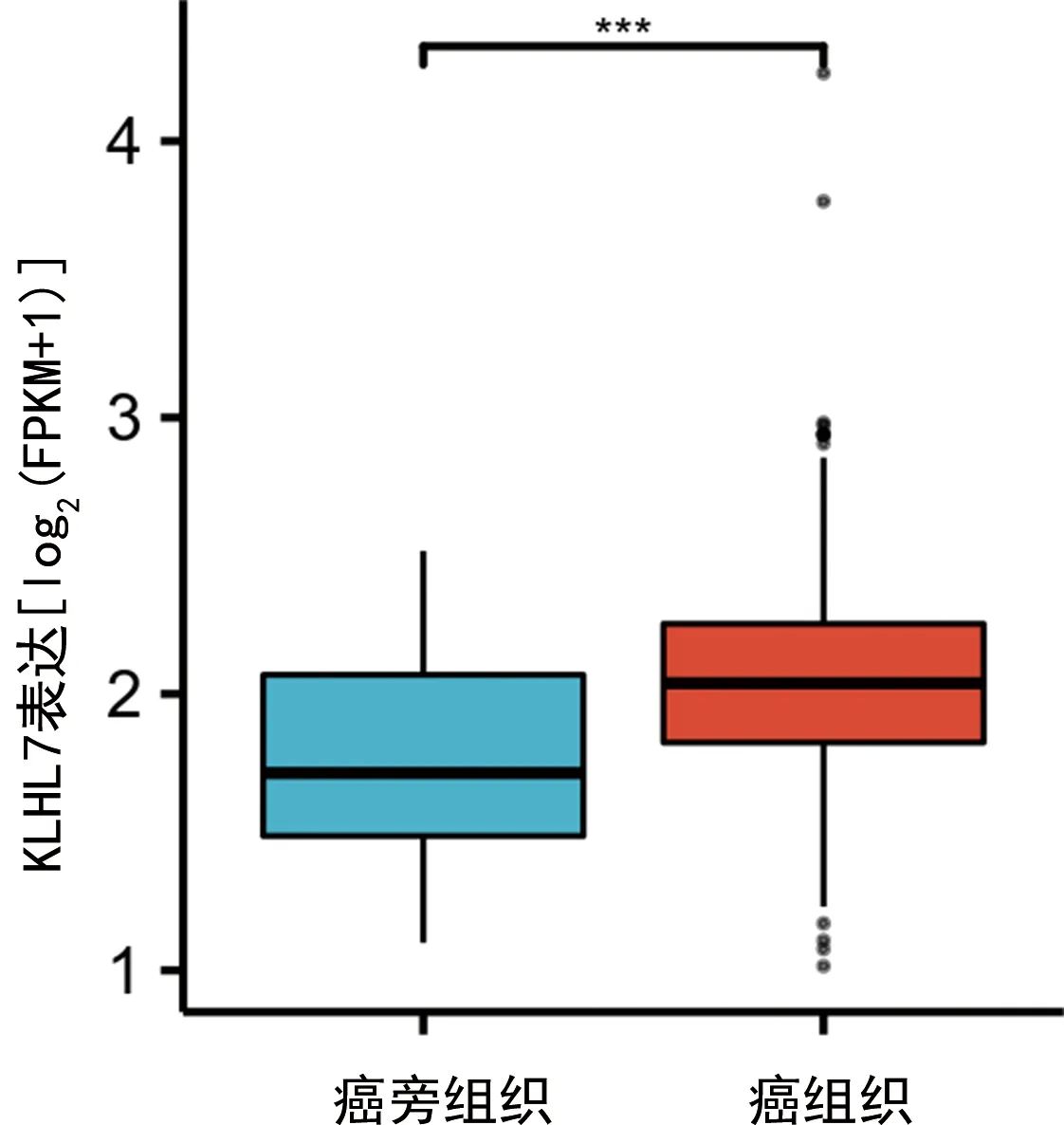
注:***P<0.001。图1 KLHL7在配对胃癌组织及癌旁组织的表达

注:*P<0.05。图2 KLHL7在非配对胃癌组织及癌旁组织的表达
2.2KLHL7基因在人胃癌组织异常表达的临床病理特征分析 基于仙桃学术数据库对高表达KLHL7基因胃癌患者的临床病理特征进行分析,结果表明,相对于正常组织,KLHL7基因在M0和M1期胃癌组织中表达水平升高(P<0.05),见图3;相对于正常组织,KLHL7基因在胃癌组织中N0、N1、N2和N3期淋巴结中表达水平升高(P<0.05),见图4;相对于正常组织,KLHL7基因在肿瘤分期为Ⅰ、Ⅱ、Ⅲ、Ⅳ期的胃癌组织中表达水平升高(P<0.05),见图5。ROC曲线分析发现,KLHL7基因诊断胃癌的曲线下面积(AUC)为0.722(95%CI:0.606~0.837),提示其对胃癌具有潜在诊断价值。对免疫浸润特征进行分析发现,KLHL7基因高表达与中央记忆型T细胞(Tcm)和辅助性T细胞(Th细胞)浸润呈正相关,与浆细胞样树突状细胞(pDC细胞)和Th17细胞浸润呈负相关,表明其与T细胞免疫和抗原呈递有关,见图6。
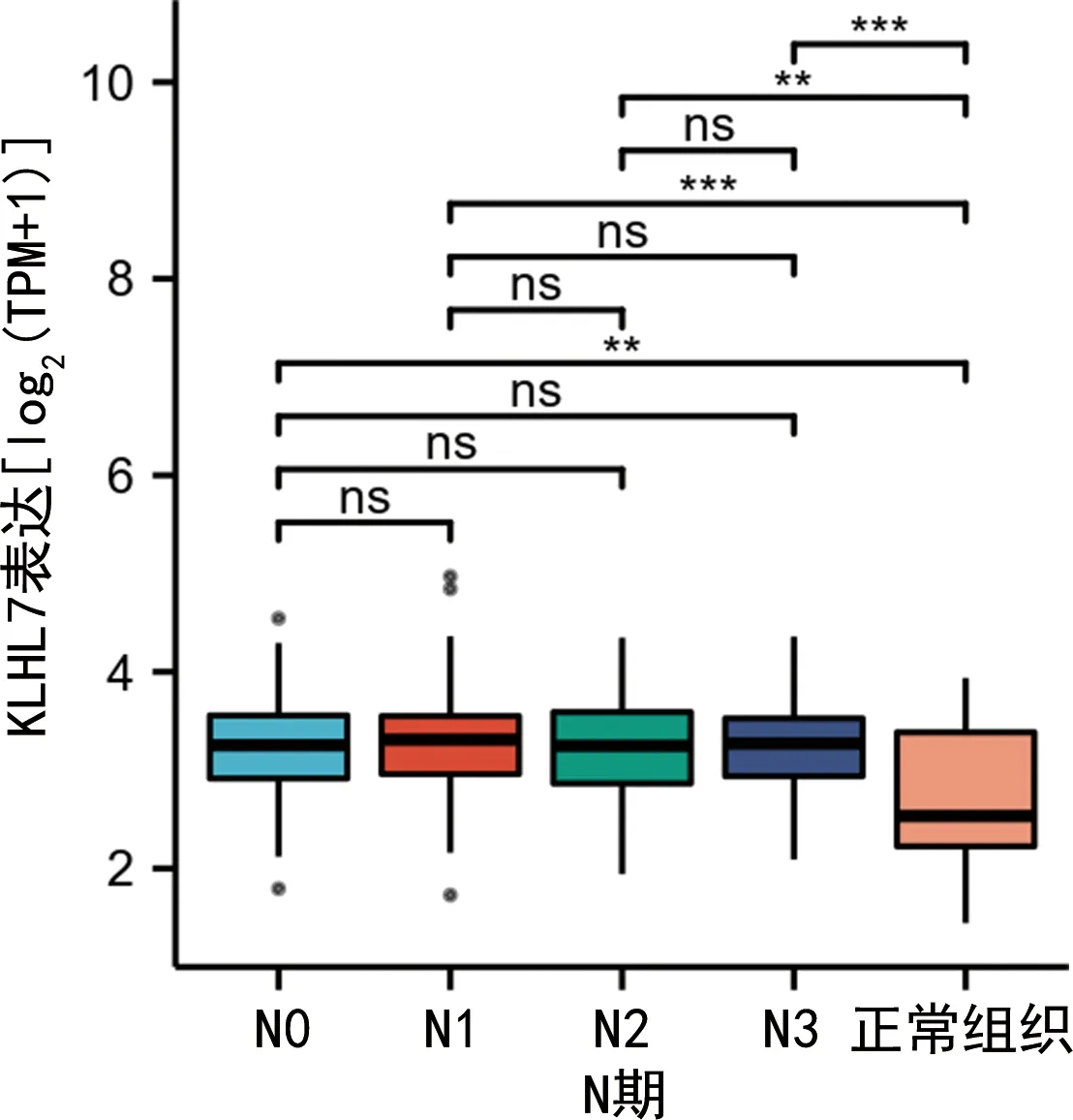
注:*P<0.05,**P<0.01,***P<0.001,ns为差异无统计学意义。图4 KLHL7在N0、N1、N2和N3淋巴结中的表达
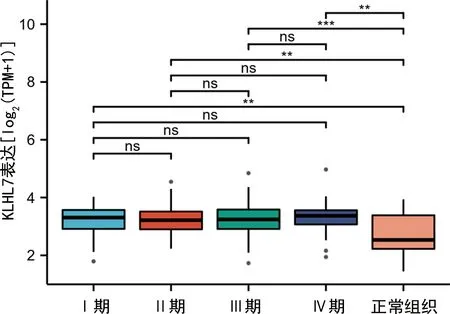
注:*P<0.05,**P<0.01,***P<0.001,ns为差异无统计学意义。图5 KLHL7在不同STAGE期胃癌组织中的表达

注:Tcm为中央记忆型T细胞;T helper cells为辅助性T细胞(Th细胞);Tem为效应记忆T细胞;NK cells为自然杀伤细胞;Tgd为gamma delta T细胞;Th2 cells为Th2细胞;Eosinophils为嗜酸性粒细胞;Th1 cells为Th1细胞;NK-CD56 bright cells为NK-CD56亮细胞;Macrophages为巨噬细胞;Mast cells为肥大细胞;TFH为滤泡辅助性T细胞;DC为树突状细胞;iDC未成熟的树突状细胞;aDC为活化的树突状细胞;Neutrophils为中性粒细胞;CD8 T cells为CD8 T细胞;T cells为T细胞;NK-CD56 dim cells为NK-CD56暗细胞;TReg为调节性T细胞;B cells为B细胞;Cytotoxic T cell为毒性T细胞;pDC为浆细胞样树突状细胞;Th17 cells为Th17细胞。图6 KLHL7在胃癌组织表达的免疫浸润特征分析
2.3与KLHL7基因具有相关性的基因的信号通路富集分析 通过cBioPortal在线数据库和Sanger学术平台分析工具分析发现,与KLHL7基因具有相关性的共表达基因有2 785个,包括上调的2 486个基因(Pearson相关系数为0.30~0.63)和下调的299个基因(Pearson相关系数为-0.47~-0.30)。这些基因富集在73个信号通路中,其中富集倍数>2为筛选标准的信号通路有33个,其中与肿瘤关系密切的有25个,分别是自噬、凋亡、细胞周期、慢性白血病、子宫内膜癌、肿瘤坏死因子(TNF)、缺氧诱导因子1(HIF-1)、结肠癌、Wnt等信号通路,见图7。
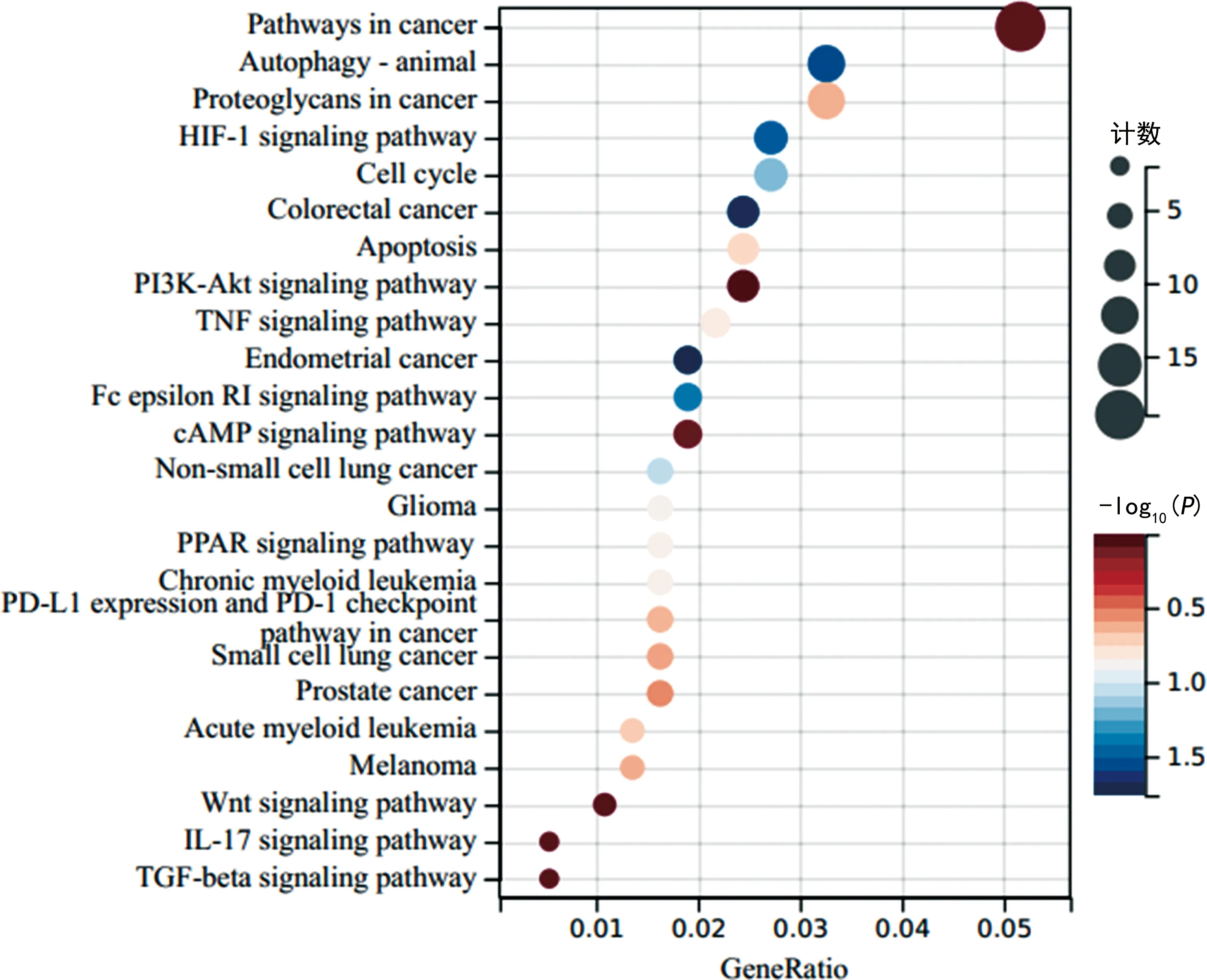
注:图中的信号通路依GeneRatio分数自上而下依次为:癌症通路、自噬、癌症中的蛋白聚糖、HIF-1通路、细胞周期、结肠癌、凋亡、PI3K-Akt通路、TNF通路、子宫内膜癌、环腺苷酸(cAMP)通路、非小细胞肺癌、胶质瘤、过氧化物酶体增殖物激活受体(PPAR)通路、慢性白血病、细胞程序性死亡配体1(PD-L1)和细胞程序性死亡受体1(PD-1)检查点通路、小细胞肺癌、前列腺癌、急性白血病、黑色素瘤、Wnt通路、白细胞介素(IL)-17通路、转化生长因子β(TGF-β)通路。图7 与KLHL7具有相关性的共表达基因富集信号通路的气泡图
2.4共表达基因中核心基因筛选分析结果 共表达基因分析结果显示,有93个基因富集在肿瘤密切相关的信号通路中;将其输入在线软件STRING进行PPI网络分析,分析其节点,使用Cytoscape 3.8 软件的CytoNCA插件计算参数筛选发现,LYN、PPP1CB、NCF1、P4HB、GAK、BBS10、RPS6KA1、H2AFV、IFT88、CD68、NFKB1、SMC3、ARL13B、RAB23、CD74、MEAF6、HYDIN、GMPPB、SEC24C、RDX共20个基因为关键基因。经GEPIA和UALCAN数据库检索,除P4HB、HYDIN、GMPPB和RDX 4个基因外,包括LYN、PPP1CB、NCF1、GAK、BBS10、RPS6KA1、H2AFV、IFT88、CD68、NFKB1、SMC3、ARL13B、RAB23、CD74、MEAF6、SEC24C的16个基因是胃癌组织与癌旁组织的差异表达基因,见图8。在以上基因中,与KLHL7相关系数>0.5的基因为BBS10、PPP1CB、ARL13B和MEAF6(Spearman相关系数分别为0.508、0.529、0.535、0.538,均P<0.001)。另外,针对这4个基因,利用仙桃学术数据库分析其癌组织与癌旁组织之间的差异表达情况,结果显示,BBS10、PPP1CB、ARL13B和MEAF6在胃癌组织中表达显著高于癌旁组织(P<0.05)。见图9。

图8 与KLHL7相关的共表达基因的核心基因筛选结果
3 讨 论
KLHL7是Kelch样家族蛋白的一员。迄今为止,已发现42种KLHL成员分子,该家族蛋白成员在结构上包含N端的BTB/POZ结构域和中间的BACK结构域,以及C端的6个Kelch重复序列。研究显示,KLHL基因参与了DNA的损伤修复、凋亡、免疫反应和维持骨骼肌结构等多种生物学过程[5]。多项研究证明,KLHL的异常表达与肿瘤的发生发展紧密关联[6-9]。有研究指出,KLHL6在胃癌中高表达,下调KLHL6的表达能显著降低MGC-803胃癌细胞集落形成、增殖和转移,增强细胞凋亡[10]。KLHL19(KEAP1)的功能缺失可激活Nrf2,为胃癌细胞的生长提供有利条件[11]。在胆管癌中,敲低KLHL21的表达显著降低胆管癌细胞的增殖、侵袭和转移,诱发G0/G1期细胞阻滞,以及降低ERK1/2的磷酸化来阻碍ERK信号通路的激活[12]。KLHL37(ENC1)表达上调可增加结直肠癌和毛细胞白血病的风险,且与卵巢癌患者的预后密切相关[13-15]。以上研究提示,KLHLs家族的标志物的异常表达在肿瘤的恶性生物学行为中扮演着重要角色,有望作为肿瘤诊断和预后判断的生物标志物。
近年研究表明,KLHL7的突变与一些遗传性疾病有关[16-19],但KLHL7在胃癌中的研究相对较少。KUROZUMI等[20]发现,KLHL7在乳腺癌中的表达水平与组织学分级和分子亚型相关,其高表达较低表达预后差。施潇娴[1]发现,KLHL7在肺癌、胃癌、卵巢癌和结直肠癌中高表达,沉默KLHL7可显著抑制肺癌、胃癌和骨肉瘤细胞的增殖。本研究采用生物信息学的方法分析发现,KLHL7在胃癌组织中高表达,其表达水平与TNM分期密切相关,提示KLHL7可能是胃癌发生发展过程中的一个重要生物标志物。免疫细胞浸润分析发现,胃癌中KLHL7高表达与Tcm和Th细胞浸润呈正相关,与pDC细胞和Th17细胞等细胞浸润呈负相关。KEGG富集分析表明,KLHL7的相关基因主要富集自噬、凋亡、细胞周期、慢性白血病、子宫内膜癌、TNF、HIF-1、结肠癌和Wnt等信号通路中。与肿瘤通路相关的16个核心基因在胃癌与癌旁组织中存在异常表达。由此推测,KLHL7在胃癌中的异常表达可能通过以上相关共表达基因和所富集的信号通路等促进胃癌的发生和发展。
与KLHL7相关系数较高的共表达相关基因BBS10、PPP1CB、ARL13B和MEAF6在胃癌组织中表达均显著高于癌旁组织。BBS10是Bardet-Biedl综合征基因(BBS)基因家族的成员,BBS10作为分子伴侣,可能影响其他睫状体或基底体蛋白的折叠或稳定性。有研究指出,该基因在乳腺癌组织中是下调表达的热休克蛋白亚型分子[21],而在胃癌中的表达和意义则鲜见研究。蛋白磷酸酶1催化亚单位β(PPP1CB)基因编码蛋白磷酸酶1(PP1)的3个催化亚基之一,而PP1是一种丝氨酸/苏氨酸特异性蛋白磷酸酶,已知参与多种细胞过程的调节,如细胞分裂、糖原代谢、肌肉收缩力、蛋白质合成和人类免疫缺陷病毒1(HIV-1)的转录。PPP1CB在胃癌中普遍表达,说明其在胃癌细胞生长中的起重要作用[22]。研究表明,PPP1CB基因与胃癌侵袭转移有关[23]。BADP核糖基化因子样GTP酶13B(ARL13B)作为ADP核糖基化因子样家族的一个成员,是一种小的GTP酶。体外和体内实验均表明,ARL13b可刺激胃癌细胞增殖、迁移和侵袭[24]。在胃癌研究中,ARL13B的表达与肿瘤大小和侵袭深度密切相关,ARL13B高表达的患者预后较差。MYST/Esa1相关因子6(MEAF6)为几种不同组蛋白乙酰转移酶复合物的组成部分,选择性剪接导致多种转录变体。沉默MEAF6基因的表达可以抑制卵巢癌细胞的增殖[25]。通过构建SDR16C5、MEAF6和SOX4 3个基因特征的结肠癌预测模型发现,MEAF6在预测结肠癌的预后中显示出较高的灵敏度和特异度,具有不同预后风险的患者还表现出基因特征的差异表达、活化的CD4记忆T细胞浸润,以及对比卡鲁胺、吉非替尼、来那度胺和伊马替尼的药物敏感性[26],而该基因在胃癌中的表达和意义尚鲜见报道。以上研究提示,与KLHL7相关系数>0.5的共表达基因BBS10、PPP1CB、ARL13B和MEAF6可能参与肿瘤的生物学行为,值得进一步深入研究其在胃癌中的共表达意义。
综上所述,本研究对胃癌组织中KLHL7表达和意义的分析结果表明,KLHL7是胃癌中高表达的癌基因,其表达与胃癌侵袭和转移有关,且与胃癌的免疫细胞浸润和预后密切相关,可作为胃癌诊断和预后评估的良好生物标志物。
