土壤中锑的形态转化及其影响因素
2023-10-29韩玉琳杨丹幸孙海涛高雅王帆罗军
韩玉琳,杨丹幸,孙海涛,高雅,王帆,罗军
(污染控制与资源化研究国家重点实验室,南京大学环境学院,南京,210023)
锑,第五周期,第ⅤA 族元素,元素符号为Sb.硫化锑矿(Sb2S3)是Sb 的主要矿石.Sb 的用途广泛,历史上,Sb 广泛用于黄金的提炼;近年来,Sb 通常用于与铅混合,以增强硬度,用于生产合金、子弹和电缆护套等[1];在聚氯乙烯、橡胶和纺织品的生产中用作阻燃剂;此外Sb 还可以作为聚对苯二甲酸乙二醇酯(PET)生产的催化剂以及玻璃器皿和颜料油漆的添加剂[2];一些治疗热带病(如内脏利什曼病)的药物也含有Sb[1].
据美国地质调查局2022 年统计,全球已探明Sb 总储量超过200 万吨.中国拥有世界上最大的Sb 储量,是世界上最大的Sb 生产国,占全球Sb 产量的55%[3].土壤中的Sb 污染大多来自采矿和冶炼,以及射击场的排污口.对锑矿区、射击场及其周围土壤中的Sb 浓度的大量调查表明:锑矿区和射击场附近土壤中的Sb 含量较高(表1).湖南、贵州和江西等大型锑矿区周围土壤中Sb 的最高含量可达4613,5681 和593 mg·kg-1,远远超过了我国土壤中Sb 的本底浓度0.8~3.0 mg·kg-1[4],挪威和博茨瓦纳的射击场附近土壤中Sb 浓度可达830 和283 mg·kg-1,高于世界卫生组织建议的土壤中Sb 的最大允许污染物浓度36 mg·kg-1[4].
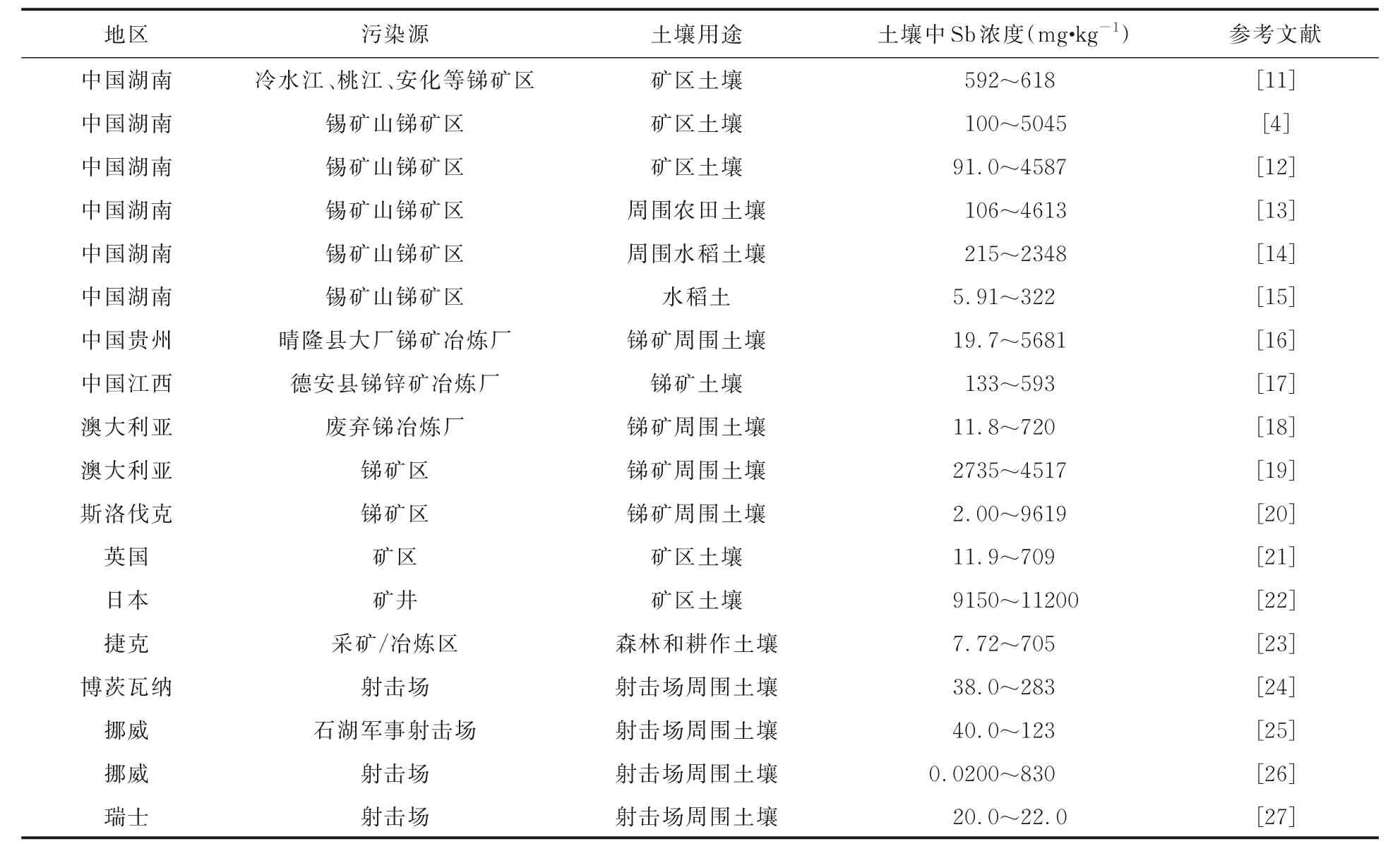
表1 不同地区矿区及射击场土壤中Sb 浓度Table 1 Sb concentration in soil of mining areas and shooting ranges in different regions
Sb 在自然界中的价态为-Ⅲ,0,Ⅲ和Ⅴ,但在土壤中主要是Ⅲ和Ⅴ两种价态,Sb 在土壤中的行为和生物有效性很大程度上取决于它的形态[5].好氧条件下Sb(Ⅴ)是热力学上最稳定的形态,以形式存在;在还原条件(如淹水的水稻土)中Sb(Ⅲ)是主要形态,以Sb(OH)3形式存在[6-7].通常,相比于Sb(Ⅴ),土壤中的Sb(Ⅲ)与膨润土、水铁矿、针铁矿、腐殖酸等土壤中的主要成分有更强的亲和力[2,8-9],更不易迁移.可能是由于的电荷密度较小,离子半径较大[8],在无定形铁氧化物上既形成内球表面络合物又形成外球表面络合物,而不带电的Sb(OH)3通过特定的内球结合,牢固地结合在铁氧化物上[10].铁矿物对Sb(Ⅲ)的吸附在较宽的pH 范围内保持不变,而对Sb(Ⅴ)的吸附随着pH 的升高而减少,酸性条件下(pH <4),针铁矿和水铁矿更有利于吸附Sb(Ⅴ),这可能是因为铁氧化物表面带正电荷,优先吸附带负电荷的.
Sb 的化学形态除了影响它在土壤中的迁移性,还决定其毒性大小.Sb 不是人体和植物必需的元素,其化合物对动、植物具有毒性,对微生物系统和土壤酶活性产生不利影响[28],对人体健康有害,是一种潜在的致癌物质[29].因此,Sb 及其化合物已被美国环境保护署和欧盟列为优先污染物,《控制危险废物越境转移及其处置巴塞尔公约》将Sb 列为危险废物[30].不同形态的Sb 毒性也不同,与砷类似,有机锑的毒性小于无机锑.在无机锑中,Sb(Ⅴ)的毒性小于Sb(Ⅲ),Sb(Ⅲ)的毒性是Sb(Ⅴ)的十倍[2].与Sb(Ⅴ)相比,植物根更易吸收Sb(Ⅲ),Sb(Ⅲ)处理组的植物体内会积累更多的Sb,而且Sb(Ⅲ)的添加刺激水稻根部脱落酸的形成,抑制根的伸长和植物的生长,刺激植物产生更高的氧化应激水平[31].然而,也有研究结果显示Sb(Ⅴ)处理组中向日葵对Sb 的累积高于Sb(Ⅲ)处理组,玉米中这两种Sb 处理组之间的Sb 吸收则没有差异[32].关于不同形态的Sb 对微生物的影响,有研究表明高剂量的Sb(Ⅲ)降低了土壤中细菌门水平的多样性指数和丰度,而Sb(Ⅴ)对细菌多样性无明显影响[33].对于不同Sb 形态对动、植物和微生物的毒性作用,还需要进行更深入的研究.
在2002-2022 年关于Sb 的综述文章中,有一部分综述了Sb 在自然水体中的分布、形态、化学行为、归趋、与微生物的相互作用、水体中除Sb技术的研究进展[30,34-35];有学者综述了Sb 的生物地球化学过程,总结了Sb 的污染现状、吸附解析、溶解浸出、氧化还原和迁移机制,归纳了受Sb 污染的土壤的修复方法[29,36].还有一些关于Sb 在土壤-植物体系中的化学行为和生物有效性方面的综述,总结了Sb 在植物中的行为、形态转化和植物缓解Sb 毒性的解毒机制[5,8].此外,有学者总结了Sb 氧化还原相关的微生物和作用机理[37-38].
土壤中的Sb 形态与其在土壤中的迁移性、生物有效性和毒性密切相关,因此了解土壤中的Sb形态转化及其影响机制至关重要.然而,目前关于Sb 在土壤中的形态及其影响因素方面的综述文章比较少[39].因此,本综述的目的在于总结淹水和好氧土壤中Sb 的存在形态,并阐述非生物因素(包括土壤基本理化性质、铁锰氧化物、N和硫化物等)和微生物对Sb 形态转化的影响机制,为更深入地了解土壤中Sb 的生物地球化学循环提供理论依据.
1 土壤中锑的形态
在好氧土壤中,Sb 在锑矿冶炼厂和射击场土壤及土壤孔隙水中以Sb(Ⅴ)形态为主[25,40].Scheinost et al[40]用Sb K 边X 射线吸收光谱测定瑞士靶场土壤样品中Sb 的形态,结果显示Sb(Ⅴ)是靶场土壤中金属Sb(0)风化后的主要氧化态,未检测到Sb(Ⅲ),金属Sb(0)的存在可能是未风化子弹碎片的一部分,Sb(Ⅴ)通过形成共边和共角内球吸附络合物吸附到铁氧化物上.Takaoka et al[41]发现锑冶炼厂排放的Sb 或Sb2O3在土壤中转化为Sb(Ⅴ)化合物.Okkenhaug et al[25]的结果表明受污染的射击场土壤中的孔隙水中仅发现Sb(Ⅴ),Sb(Ⅲ)浓度低于检测限;Sb(Ⅴ)在孔隙水中占主导地位可能是由于Sb(Ⅲ)优先吸附到土壤颗粒上,且铁氧化物可以快速氧化Sb(Ⅲ),Sb(Ⅴ)从土壤表面解吸[42].
有研究表明,在淹水条件下,当Eh 从200 mV降至-150 mV 时,大量的Sb(Ⅲ)被释放到土壤溶液中,且Sb(Ⅲ)浓度随着Eh 的降低而稳定增加[43].土壤由好氧转变为厌氧时,由本地微生物活动引发的还原条件导致了Sb(Ⅴ)被转化为Sb(Ⅲ),在土壤溶液中也发现了Sb(Ⅲ)的存在[44].研究发现只有在厌氧条件下才能观察到Sb(Ⅲ)的积累,稻田土厌氧处理的第90 天时会产生大量的Sb(Ⅲ),而在好氧条件下则没有[45].
与此相反的是,许多研究表明,虽然在还原条件下,Sb 在热力学上倾向于转化为Sb(Ⅲ),但是Sb(Ⅴ)仍然是淹水土壤中Sb 的主要形态.Mitsunobu et al[22]发现在土壤-水系统中,Sb 在较宽的氧化还原范围内(Eh 为360 mV 至-140 mV)仅以Sb(Ⅴ)形态存在.Okkenhaug et al[46]也发现在Eh 为-40 mV 的淹水土壤中,Sb(Ⅴ)仍然是土壤基质和孔隙水中Sb 的主要形态.Xia et al[47]的研究也发现溶解的Sb(Ⅴ)是厌氧土壤中Sb 的主要形态,在厌氧培养的前十天里Sb(Ⅲ)的浓度略微增加,从9.83 μg·L-1增加至32.6 μg·L-1,随后逐渐下降,在厌氧培养60 d 结束时未检测到Sb(Ⅲ).
此外,有机锑在土壤中含量较少,毒性低,因此对其的关注和研究比较少.有研究者在锡矿山土壤剖面的上层中检测到了痕量的甲基锑(<0.4%)[48].还有研究发现Sb 的甲基化可能发生在诸如涝渍土壤等环境中,土壤样品中Sb 化合物的生物介导还原和甲基化已得到证实[49].
Sb 的氧化还原循环受到土壤的基本理化性质(如pH,Eh 等)影响,并和土壤中其他元素(如Fe,Mn,N 和S 等)的生物地球化学循环相耦合[39],土壤中的化学反应过程和微生物介导的生物化学过程,都会造成Sb 的形态转化[50],改变土壤中Sb 的形态.
2 土壤中锑形态转化的影响因素
2.1 土壤理化性质对锑形态的影响如前所述,好氧和厌氧土壤中Sb 主要的存在形态有所不同,可见土壤的氧化还原状态会影响土壤中Sb 的形态.土壤的氧化还原电位会影响可用的电子受体,改变土壤中的微生物群落结构,影响土壤中Sb 氧化还原菌的生长状态,从而影响土壤中Sb的形态.在好氧土壤中(Eh>300 mV),土壤中存在的O2为主要的电子受体,负责Sb 氧化的基因(如aioA)显著增加,促进Sb(Ⅲ)氧化[47,51];在环境中,以O2为唯一氧化剂的Sb(Ⅲ)的氧化非常缓慢,因为高岭石等物质具有较大的比表面积,增加了吸附的Sb(Ⅲ)与O2的接触面积,促进Sb(Ⅲ)氧化为Sb(Ⅴ)的速率,Sb(Ⅲ)氧化产生大量的Sb(Ⅴ)[52].当淹水导致土壤或沉积物缺氧时,微生物逐渐被迫选择能量较低的末端电子受体进行呼吸反应.在Fe(Ⅲ)还原为Fe(Ⅱ)之后,硫酸盐还原为硫化物之前,Sb(Ⅴ)被还原到Sb(Ⅲ),Sb(Ⅲ)还原为单质锑或锑化氢[49].
土壤pH 也是影响土壤中Sb 形态转化的原因之一.O2和H2O2氧化Sb(Ⅲ)的反应都与pH 有关,相比于O2,过氧化氢的氧化反应要快得多[53].除此之外,Sb(Ⅲ)容易被各种化合物吸附,如Fe,Mn,Al 的氢氧化物、腐植酸和土壤中的黏土矿物,这些化合物可以诱导Sb(Ⅲ)的催化氧化,诱导过程受pH 的限制[37].Kong et al[54]发现Sb(Ⅲ)在黄铁矿表面被氧化为Sb(Ⅴ),随着pH 的升高,Sb(Ⅲ)的氧化效率逐渐提高.pH 还会影响针铁矿上Sb(Ⅲ)的氧化,在不同的pH 条件下,Sb(Ⅲ)的氧化途径也不同.在pH 为3 时,Sb(Ⅲ)的氧化效率最高,这是由于Fe(OH)2+浓度高,生成·OH的产率高,H+可以诱导产生HO2·,促使反应中产生过氧化氢,加快Sb(Ⅲ)的氧化速率[55].此外,Sb(Ⅲ)的形态与pH 有关,Sb(Ⅲ)的两个半反应Sb(OH)3(酸性和中性条件下Sb(Ⅲ)的形态)氧化为Sb(碱性条件下Sb(Ⅲ)的形态)氧化为的自由能分别为110.6 kJ·mol-1和45.8 kJ·mol-1,因此,比Sb(OH)3更容易被氧化剂氧化[55].pH 可以改变土壤氢离子的浓度,从而影响土壤中细菌群落的组成和多样性[12],且相较于淹水土壤,pH 对好氧土壤中细菌群落的影响更大[56].其次,pH 是影响Sb(Ⅲ)微生物氧化速率的关键因素,pH 通过影响Sb 氧化菌的生长繁殖速率、酶蛋白的活性、Sb(Ⅲ)非酶促反应(H2O2催化反应)等方式影响Sb(Ⅲ)的微生物氧化速率[37].细菌的能量呼吸、生理和生长直接受到pH 的影响,如果土壤pH 落在相对较窄的生存窗口外,大多数个体分类群就无法生存[12].
2.2 土壤中铁矿物对锑形态的影响
2.2.1 Fe(Ⅲ)介导的光催化氧化Sb(Ⅲ)铁离子和铁(氢)氧化物广泛分布于天然土壤中,Sb(Ⅲ)与Fe(Ⅲ)的相互作用可能是控制Sb 形态的重要过程,不同形态Fe(Ⅲ)的光催化氧化在光照下表层土壤中Sb和Fe的地球化学循环中起重要作用[57].Sb(Ⅲ)的光氧化机理因Fe(Ⅲ)形态不同而不同.
在酸性溶液(pH=1~3)中,铁羟基络合物Fe(OH)2+、羟基自由基(·OH)、二氯自由基(Cl2·-)和超氧阴离子(·)是Sb(Ⅲ)氧化的主要氧化剂.Fe(OH)2+形态由Fe(Ⅲ)的水解产生,是在光照(hv)下产生羟基自由基(·OH)的有效来源[55,58-59].在低pH 条件下,与Fe(OH)2+的光化学反应可能对Sb(Ⅲ)的氧化有很大贡献[58].在酸性溶液中,Fe(C2O4)可与H2O2反应生成·OH,将Sb(Ⅲ)快速光氧化[59].然而,在Cl-存在下,Sb(Ⅲ)的氧化速度要快得多,上升了两个数量级.这种催化作用可以归因于铁-氯络合物(FeCl2+)的形成,该络合物吸收光子(hv)以产生高度氧化的二氯自由基(·C),FeCl2+在光催化下可以生成·OH[58].在光的诱导下,针铁矿可以将Sb(Ⅲ)氧化为Sb(Ⅴ),针铁矿表面在辐照过程中产生空穴电子对(ecb--hvb+),起半导体作用,在该反应中会产生羟基自由基(·OH)和超氧阴离子(·).此外,Sb(Ⅲ)的氧化是由Fenton 反应(H2O2和Fe2+)引起的,会产生更多的·OH.因此,Fe2+的存在对针铁矿上Sb(Ⅲ)的光氧化是有利的[55].
在pH=4~10 的条件下,Fe(Ⅲ)逐渐转化为胶体氢氧化铁,Sb(Ⅲ)通过内球络合作用吸附在氢氧化铁上形成络合物,光氧化是通过Sb(Ⅲ)向Fe(Ⅲ)的电子转移以及Fe(Ⅲ)向Fe(Ⅱ)的配体-金属电荷转移(LMCT)实现[57].H2O2是中性和碱性下黄铁矿诱导Sb(Ⅲ)氧化的主要氧化剂,光照条件下,黄铁矿诱导产生更多的·OH 和H2O2,Sb(Ⅲ)的氧化效率显著提高[54].Kong and He[59]发现在近中性(pH=6.2)条件下,Fe(C2O4)可与H2O2反应生成Fe(Ⅳ),是Sb(Ⅲ)的主要氧化剂.
2.2.2 无光照时铁矿物氧化Sb(Ⅲ)在没有光照时,铁(氢)氧化物可以对Sb(Ⅲ)非生物氧化.Sb(Ⅲ)在铁(氢)氧化物上的吸附加速了Sb(Ⅲ)的氧化,吸附到颗粒表面的O2是铁(氢)氧化物存在下Sb(Ⅲ)氧化的原因.针铁矿具有较大的比表面积,增加了吸附的Sb(Ⅲ)与氧气之间的接触面积,从而促进氧化速率[60].Guo et al[2]还发现吸附在水合氧化铁(HFO)上或与FeCl3共沉淀的大部分Sb(Ⅲ)被氧化成Sb(Ⅴ),这可能是由于长时间的样品保存过程中O2的参与.此外,Sb(Ⅲ)与针铁矿表面的配位增加了Sb 原子的电子密度,从而促进了氧化过程[6].非生物电子转移是吸附在铁氢氧化物上的Sb(Ⅲ)被氧化的另一个机制,首先Sb 的吸附和铁氢氧化物形成表面络合物,一个Sb(Ⅲ)在两个Fe(Ⅲ)位上,然后两个电子从Sb 转移到两个铁原子,生成Sb(V)和Fe2+[61].
Fe(Ⅱ)引起的铁矿物的转化也是氧化Sb(Ⅲ)的重要途径.水铁矿吸附Sb(Ⅲ)的过程中,亚铁水合物表面作为催化剂被溶解氧氧化,通过非生物电子从Sb(Ⅲ)转移到Fe(Ⅲ)促进Sb(Ⅲ)的氧化[10].Fe(Ⅱ)氧化过程中,会产生活性氧物种(ROS),在Sb(Ⅲ)的氧化中发生重要作用.Fe(Ⅱ)介导的水铁矿的转化过程中,超氧阴离子自由基(·)在Fe(Ⅱ)和结构Fe(Ⅲ)之间的电子转移过程中产生,促进了Sb(Ⅲ)的氧化.ROS 也可以在Fe(Ⅱ)和Fe(Ⅲ)矿物结构内部的电子转移过程中产生,使得Sb(Ⅲ)的氧化不仅发生在铁羟基氧化物的表面上,而且发生在所产生的针铁矿和/或纤铁矿的晶体结构中.Sb(Ⅲ)在铁羟基氧化物表面的氧化速度远快于晶体结构内部[62].Zhu et al[63]得到了类似的结论,Fe(Ⅱ)氧化生成铁(氢)氧化物,过程中通过电子转移生成ROS/Fe(Ⅳ),将吸附的Sb(Ⅲ)氧化.Kong et al[54]发现Sb(Ⅲ)可以在黄铁矿表面被氧化,在缺氧条件下,黄铁矿中原始的Fe(Ⅲ)可与H2O 反应生成H2O2和·OH,一旦原始的Fe(Ⅲ)耗尽,氧化反应停止.但在氧气充足时,Fe(Ⅱ)可与O2反应生成过氧化氢,同时在缺硫缺陷位提供Fe(Ⅲ),然后吸附的H2O 与Fe(Ⅲ)反应,生成吸附的·OH 和H2O2,完成对Sb(Ⅲ)的氧化.Leuz et al[64]发现在酸性时,Fe(Ⅱ)和H2O2协同产生·OH自由基,氧化速度加快,而在中性的条件下,可能存在中间体Fe(Ⅳ),在Sb(Ⅲ)的氧化过程中发挥了重要作用.
另外,水稻根释放氧气时,Fe(Ⅱ)可能被氧化并以铁膜的形式覆盖在根表面,这些铁膜主要由亚铁水合物和少量针铁矿组成[7].即使在Sb(Ⅲ)处理组中,水稻植株中仍以Sb(Ⅴ)为主,这可能是由于Sb(Ⅲ)被根表面的铁膜氧化[65].
2.2.3 铁矿物还原Sb(Ⅴ)除了氧化Sb(Ⅲ),有些铁矿物可以将Sb(Ⅴ)还原.在土壤中,水铁矿通过异化铁还原菌或硫化物还原,可形成一系列含Fe(Ⅱ)的矿物,包括菱铁矿、磁铁矿、绿锈和硫化亚铁,这些都是潜在的Sb 还原剂[66].Johnson et al[66]发现四方硫铁矿是将Sb(Ⅴ)还原为Sb(Ⅲ)的主要还原剂,在四方硫铁矿存在的情况下,Sb(Ⅴ)通过还原为Sb(Ⅲ)和无定形Sb2S3的沉淀从溶液中去除.同样地,Kirsch et al[67]发现磁铁矿和四方硫铁矿可通过表面Fe(Ⅱ)的氧化将Sb(Ⅴ)还原为Sb(Ⅲ),而Sb(Ⅲ)没有进一步还原.绿锈存在于许多土壤和沉积物系统中,是铁(羟基)氧化物(如针铁矿、纤铁矿和磁铁矿)形成的中间相.Mitsunobu et al[68]研究表明,绿锈可以将Sb(Ⅴ)还原为毒性更大的Sb(Ⅲ),可能是Sb(V)最重要的还原剂之一,与其他铁矿物不同的是,绿锈对Sb(Ⅴ)的亲和力大于Sb(Ⅲ).
2.3 土壤中锰氧化物对锑形态的影响锰氧化矿物在土壤中也广泛存在,是土壤和沉积物中Sb的重要吸附剂和氧化剂.锰氧化物对Sb(Ⅲ)有明显的快速氧化作用,并在几分钟内就可以将Sb(Ⅲ)氧化,形成可溶的Sb(Ⅴ)[69].锰氧化物氧化Sb(Ⅲ)主要有两种途径:(1)一个Sb(Ⅲ)向一个Mn(Ⅳ)提供两个电子,然后一个Sb(Ⅴ)和一个还原的Mn(Ⅱ)被释放到溶液中;(2)两步反应机制:一个Sb(Ⅲ)分子向两个Mn(Ⅳ)分子提供两个电子,生成一个Sb(Ⅴ)和两个Mn(Ⅲ)分子,然后在矿物表面新形成的两个活性Mn(Ⅲ)中心将一个新的Sb(Ⅲ)氧化成Sb(Ⅴ)[70].
有研究表明在铁锰氧化物吸附、氧化Sb 的过程中,锰氧化物主要负责氧化Sb(Ⅲ),铁氧化物则是主要的吸附中心,例如铁锰复合氧化物FMBO 和生物铁锰氧化物[71-72].铁锰复合氧化物FMBO 通过Sb(Ⅲ)向Mn(Ⅳ)的界面电子转移实现氧化[71].Mn(Ⅱ)和Mn 氧化细菌共存时,锰氧化细菌原位氧化生成生物锰氧化物(BMO),能将Sb(Ⅲ)氧化成Sb(Ⅴ),但是Sb(Ⅴ)离子由于静电电荷排斥作用没有被BMO 明显吸附.在引入Fe(Ⅱ)后,混合形成生物铁锰氧化物(BFMO),氧化Sb(Ⅲ)并改善Sb(Ⅴ)吸附.与合成的铁锰复合氧化物类似,BFMO 的锰氧化物部分主要负责Sb(Ⅲ)的氧化,不同之处在于,由于其生物性质,Mn(Ⅱ)和Fe(Ⅱ)离子不会再次释放到水中,细菌会再次将Mn(Ⅱ)离子氧化为BMO[72].
2.4 土壤中硝酸盐和硫化物对锑形态的影响在缺氧土壤和沉积物中,在Sb 的形态转化方面发挥着重要作用.硝酸盐可以促进淹水土壤中的Sb(Ⅲ)氧化.文献表明,土壤中硝酸盐浓度的增加显著增加了土壤中As(Ⅲ)氧化基因aioA基因的丰度[73],由于Sb 和As 具有相似的化学性质,aioA基因可能是参与Sb(Ⅲ)氧化的关键基因[45].此外,在缺氧环境中,硝酸盐作为电子受体可以耦联Sb(Ⅲ)氧化,从而促进淹水土壤中Sb(Ⅲ)的氧化[74].硫酸盐在缺氧环境中下被异化还原为硫化物,生成的硫化物可能将Sb(Ⅴ)还原为Sb(Ⅲ),生成元素硫或者沉淀,极大地降低了Sb 的迁移性.除了SO,Ye et al[76]发现了一个通过S0将针铁矿上的SbⅢ-O 转化为SbV-S的新途径.即异化金属还原菌还原S0,生物硫化物诱导SbⅢ-O 生成硫代锑矿(SbⅢ-S),然后SbⅢ-S被S0氧化成SbV-S.
2.5 土壤中微生物对锑形态的影响
2.5.1 锑氧化菌氧化Sb(Ⅲ)除了非生物因素的影响,微生物在Sb 的形态转化上也发挥着不可或缺的作用.在受Sb 污染的土壤中广泛存在多种典型的Sb 氧化细菌,能将毒性较强的Sb(Ⅲ)转化为毒性相对较弱的Sb(Ⅴ),是Sb 生物地球化学循环的主要参与者[37].表2 展示了在锑矿区土壤中鉴定分离的Sb 氧化菌.目前已鉴定出97株Sb 氧化细菌,其中97.7%属于变形菌门;在属水平上,Sb 氧化菌主要分布于Pseudomonas,Comamonas,Acinetobacter,Rhizobium,Sphingomonadaceae,Paraccocus,Stenotrophomonas,Hydrogenophaga等[37,77-80];Zhang et al[74]发现Azoarcus,Azospira,Chelativorans等能以硝酸盐为电子受体厌氧氧化Sb(Ⅲ),含有aioA基因和参与反硝化作用和碳固定的基因.

表2 锑矿区土壤和沉积物中的Sb 氧化菌Table 2 Antimony-oxidizing bacteria in soil and sediments in antimony mining areas
Sb(Ⅲ)的微生物氧化机制是酶促氧化机制和非酶促氧化机制共同作用的结果.Sb 氧化菌细胞特异性Sb 氧化基因-酶系统主要包括aioAAioA,anoA-AnoA,aoxR-AoxR,aoxAB-Aox-AB,iscR-IscR,arsH-ArsH,mrpB-MrpB 等[37].基因和酶系统在不同的细菌中所起的作用有差异,Li et al[81]发现A.tumefaciensGW4 中AnoA 对Sb(Ⅲ)氧化的影响可能高于AioAB,但是在A.tumefaciens5A 中,AioAB 对Sb(Ⅲ)氧化效率有重大贡献.非特异性氧化体系主要由GHS,ROS,H2O2等组成,它们构成了Sb 氧化细菌的氧化应激调节机制,可以作为非生物氧化剂催化细菌Sb(Ⅲ)的氧化[37].
2.5.2 锑还原相关菌还原Sb(Ⅴ)在厌氧土壤中,Sb 的异化还原途径占上风,负责Sb 还原的基因显著丰富,与Sb 还原相关的细菌类群得到富集,Geobater和Clostridiaceae等潜在的Sb 还原菌显著丰富,促进Sb(Ⅴ)还原[45-56].表3 显示了Sb污染土壤中的Sb 还原菌,属水平上主要分布于Geobacter,Pseudomonas,Clostridium,Arthrobacter,Rhizobium,Bacillus等[47,83,86-88].这些微生物在Sb 污染土壤和沉积物中富集,这些属中的许多物种可以还原金属和类金属,因此具有还原Sb(Ⅴ)的潜力[75],但是相较于Sb 氧化菌,Sb 还原菌的相关数据较少,仍需要进一步确定微生物在Sb(Ⅴ)还原中的作用,分离鉴定更多的Sb(Ⅴ)还原菌.有研究者在Sb 污染土壤和沉积物中分离出可以还原Sb(Ⅴ)的菌株,例如Lysinibacillus,Dechloromonas,菌株MLFW-2(芽孢杆菌目)、菌株Juk-1(中华根瘤菌属)和菌株CNZ-1[86,89-91].由于As 和Sb 具有相似的结构,因此微生物可能使用相似的代谢途径来转化As 和Sb,As(Ⅴ)还原基因,如arsC和arrA等都可以转化Sb(Ⅴ),Sb 还原基因anrA也可以参与Sb(Ⅴ)的还原[86,92].

表3 锑污染土壤和沉积物中的Sb 还原菌Table 3 Antimony-reducing bacteria in antimony-contaminated soils and sediments
另外,铁和硫的地球化学循环可能会与Sb 的化学循环结合起来,影响Sb 的转化.硫还原菌的存在可以将硫酸盐离子转化为硫化物,生成的硫化物实现对Sb(Ⅴ)的非生物还原[93];Fe(Ⅱ)的存在可以通过提高氢化酶的活性加速H+和硫酸盐的消耗,进而促进硫还原菌的代谢活性,使Sb(Ⅴ)以硫化物沉淀的形式生物还原为Sb(Ⅲ)(Sb2S3)[94].这些对于缺氧环境如淹水土壤和沉积物中Sb 的生物化学转化具有重要意义.
3 总结展望
Sb 的形态会影响其迁移性与毒性,土壤中多种因素会影响Sb 的形态,本文综述了土壤中Sb的形态及其非生物与生物影响因素,主要得出以下结论.
(1)好氧条件下,Sb(Ⅴ)是Sb 污染土壤中主要的存在形态;淹水土壤中存在大量的Sb(Ⅲ),但即使热力学上Sb(Ⅲ)在还原系统中占主导地位,Sb(Ⅴ)仍是厌氧土壤中Sb 的主要形态.
(2)土壤的Eh 和pH 可以通过影响土壤中微生物的群落结构和组成影响土壤中的Sb 形态,铁锰氧化物和腐殖酸等对Sb(Ⅲ)的氧化速率也受pH 的影响.土壤中存在的铁锰氧化物、N和SO在Sb 的形态转化方面也发挥着重要作用.Fe(Ⅲ)可以介导Sb(Ⅲ)的光催化氧化,且Sb(Ⅲ)的光氧化机理因Fe(Ⅲ)形态不同而不同.在没有光照时,铁(氢)氧化物也可以氧化Sb(Ⅲ),主要存在三个途径:增加Sb(Ⅲ)与O2的接触面积;通过非生物电子转移实现对Sb(Ⅲ)的氧化;产生·OH,·,Fe(Ⅳ),H2O2等物质,将吸附的Sb(Ⅲ)氧化.此外,一些含Fe(Ⅱ)的矿物可作为潜在的Sb 还原剂,如菱铁矿、磁铁矿、绿锈和硫化亚铁等,可将Sb(V)还原为Sb(Ⅲ).土壤中的锰氧化物主要通过电子转移进行Sb(Ⅲ)的氧化,且在铁锰复合氧化物中,锰氧化物部分主要负责氧化Sb(Ⅲ).在厌氧条件下,N会增加Sb 氧化基因的丰度,同时作为电子受体耦合Sb(Ⅲ)氧化,促进Sb(Ⅲ)的厌氧氧化;而SO被还原为S2-,硫化物可能将Sb(Ⅴ)还原为Sb(Ⅲ).此外S0可被异化金属还原菌还原,将硫代锑矿(SbⅢ-S)氧化成SbⅤ-S.
(3)土壤中的微生物也可以影响土壤中Sb 的形态.首先,Sb 污染土壤中广泛存在Sb 氧化菌,多种特异性Sb 氧化基因-酶系统(aioA-AioA,anoA-AnoA,aoxR-AoxR 等)以及非特异性氧化体系(GHS、ROS 和H2O2等物质)可通过酶促氧化机制和非酶促氧化机制,将毒性较强的Sb(Ⅲ)氧化为毒性相对较弱的Sb(Ⅴ);其次,Sb 污染土壤中多种Sb 还原菌通过Sb 还原基因(arsC,arrA,anrA等),参与Sb(Ⅴ)的还原;另外,硫酸盐还原菌可将SO还原为S2-,实现对Sb(Ⅴ)的还原.
上述研究已经基本明确土壤中Sb 的存在形态,多方面解释了影响Sb 化学形态转化的非生物和生物因素.基于上述总结,可概括今后需要继续深入研究的方向如下:
(1)研究更多不同性质缺氧土壤中Sb 的化学形态,进一步完善Sb(Ⅲ)的测定及保存方式,为更准确地测定厌氧土壤中Sb 的存在形态提供技术支持.
(2)与Sb 形态转化相关的微生物及其机制的研究较少,特别是与Sb 还原相关的微生物,可从Sb 污染土壤和沉积物中鉴定分离更多的Sb 转化菌,以便对土壤中Sb 的生物转化有更深入的了解.
(3)关于不同价态的Sb 在生物体内的毒性数据较少,需要进一步开展研究.
(4)由于有机锑毒性较小,而且在土壤中含量较低,本综述暂未考虑有机锑和无机锑之间的转化,后续可进一步研究有机锑在土壤及生物体中的迁移转化,更全面地探讨Sb 的生物地球化学循环.
