干旱及复水对冬小麦叶片生理指标和光合特性的影响
2023-10-28崔兆韵邹俊丽尹逊栋江梦圆
崔兆韵,徐 祎,邹俊丽,尹逊栋,江梦圆
(1.山东省气象防灾减灾重点实验室,济南 250031;2.泰安市生态与农业气象中心,山东 泰安 271000;3.泰安农业气象试验站,山东 泰安 271000;4.泰安市农业科学院,山东 泰安 271000;5.南京信息工程大学,南京 210044)
0 引 言
小麦是我国主要的粮食作物,黄淮海地区是我国最大的小麦产区,产量和种植面积分别约占全国小麦的76%和68%[1]。该地区属温带大陆性季风气候,降水在时间和空间上分布极不均匀,全年60%~80%的降水集中在6-9 月份[2],庄严等[3]研究表明,冬小麦的需水和耗水量在分蘖和拔节期出现峰值,拔节到开花期是小麦水分敏感重要期,此时正处在相对干旱的阶段,土壤水分对小麦的生长有很大的影响。近50 a来,当地气候变化呈现温度升高、降水减少态势,水资源短缺成为制约农业生产发展的关键因素[4],不同干旱条件下,冬小麦产量表现为水分条件适宜>轻度干旱>重度干旱[5]。
叶片是作物光合作用的重要场所,干旱会影响叶片的生理指标和光合特性,进而影响作物产量[6]。植物在受到干旱胁迫时,体内会产生大量的活性氧,清除活性氧的酶促系统就会被激活,超氧化物岐化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)等酶的活性显著升高[7]。随着干旱程度的加剧,当产生的活性氧不能被酶促系统及时清理时,就会导致细胞膜发生膜脂过氧化,丙二醛(MDA)的含量迅速上升[8],给细胞造成伤害。即使后期复水,细胞膜修复也缓慢或者不能修复[9]。复水后,叶片SOD 活性降低,MDA 的含量减少,膜透性恢复很快,酶促系统修复了光合机制[10]。同时,干旱胁迫可以引起植物光合作用减弱,植物光合作用的强弱通常用光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、细胞间隙CO2浓度(Ci)和水分利用效率(WUE)来反映。干旱胁迫可以使叶片的Gs、Pn、Tr 值降低,叶绿素降解,使叶片的光合作用能力显著降低[11],干旱胁迫程度越强,下降幅度越大[12]。复水后,叶片光合作用得到补偿,Pn 和Tr 明显增大[10],Gs 和Ci有所回升[13]。有研究表明,光合速率的降低与干旱程度有关,气孔因素在不同干旱程度下,对光合速率的抑制发挥不同的作用[14]。目前,有关干旱后复水对冬小麦叶片生理生化指标及光合特性影响的研究较多,而在冬小麦拔节-开花期的关键需水期,干旱胁迫程度达到特旱条件后复水,对冬小麦叶片生理生化指标及光合特性影响的研究较少。本研究拟通过研究拔节-开花期不同干旱强度胁迫条件下及复水后,“济麦22”生理指标和光合特性的变化规律,为冬小麦水分高效利用提供科学支撑。
1 材料与方法
1.1 试验地介绍
该试验于2018-2019年在山东省泰安农业气象试验基地土壤水分控制场进行,试验场设有全自动控制遮雨棚,棚顶及四周采用阳光板遮蔽,能完全隔绝降水,阳光板透光率85%,保证光照充足。试验所在地年均气温15.1 ℃,年均降水量约700 mm,土壤为沙壤土。控水试验前施肥及灌溉等管理措施与大田一致,保证出苗率。
1.2 试验设计
王东博等[15]研究表明,拔节期亏水对冬小麦生理指标影响显著。试验在小麦关键需水期拔节-开花期进行水分控制处理。试验开始前将20 cm 土层土壤相对湿度保持在60%左右。试验地4 月常年降水量在30 mm 左右,当地大田灌溉量4 月份在45 mm左右,试验以75 mm作为正常补水量,将正常补水量的80%、50%、25%和0%作为4 个处理(以T1、T2、T3 和T4表示),以正常补水量作为对照(CK),在冬小麦进入拔节期(4月2日)后进行一次性补水,之后不再补水。在冬小麦进入开花期(4月26日)后复水,将各试验小区土壤相对湿度提高至90%,与正常灌溉管理的大田冬小麦的土壤相对湿度一致,之后灌溉管理措施均相同。每个试验小区是一个长方体状土壤水分控制池,控制池上表面长4 m,宽4 m,高2.3 m,内装2 m 深土层,控制池四周及底部均已作20 cm 厚的防渗处理,保证各试验小区间互不影响,每个处理3 次重复,共15 个试验小区,随机区组设计。
1.3 指标测定
(1)土壤相对湿度(R)的测定。采用烘干称重法,从4月3 日开始每隔5 d 测定一次10~20 cm 土层的土壤相对湿度,复水后每隔10 d 测定一次。每个试验小区随机取4 个样点,4个样点的平均作为该小区的土壤相对湿度,计算公式如下:
式中:R 为土壤相对湿度,%;w 为土壤含水量,%;f为土壤田间持水量,%,根据试验站多年监测,土壤田间持水量约为11%。
(2)酶活性和丙二醛含量。从4 月3 日开始每隔10 d,于上午8∶00-10∶00,选取每个小区植株上部全展叶片,处理测定超氧化物岐化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和丙二醛(MDA)的含量,每个处理重复3 次。SOD活性采用氮蓝四唑法测定,POD 活性采用愈创木酚法测定,CAT 活性采用高锰酸钾滴定法测定,丙二醛含量采用硫代巴比妥酸(TBA)法测定[16]。
(3)光合参数的测定。从4 月3 日开始每隔10 d,于上午9∶00-12∶00,选取每小区植株生长健壮的叶片,利用Li-6400便携式光合测定系统,测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci),每个处理重复3次。叶片的水分利用效率(WUE)利用公式WUE = Pn/Tr 计算[17];气孔限制值(Ls)利用公式Ls = 1 - Ci/C0计算(C0为外界CO2浓度)[18]。
1.4 数据处理
利用Excel 2010 进行数据处理与作图,用SPSS 17.0 进行数据差异性分析。
2 结果与分析
2.1 各处理10~20 cm土层土壤相对湿度的变化
图1 为各处理10~20 cm 土层土壤相对湿度随时间的变化过程。国家《农业干旱等级》(GB/T 32136-2015)标准规定,根据土壤相对湿度指数(Rsm)的大小,将干旱等级划分为4级:轻旱(50%<Rsm≤60%)、中旱(40%<Rsm≤50%)、重旱(30%<Rsm≤40%)、特旱(Rsm≤30%)。由图1 可知,T1 处理大约在控水10 d 后进入轻旱水平,末期处于中旱水平;T2、T3处理大约在控水12 d 后进入中旱水平,末期处于重旱水平;T4 处理没有灌水,很快进入干旱阶段,大约12 d 后进入重旱水平,末期处于特旱水平,冬小麦在控水阶段基本都处于干旱胁迫状态。
2.2 干旱后复水对冬小麦叶片生理指标的影响
由图2可知,随着时间的延长,各处理SOD活性均呈现先升高后降低的趋势。除T4 处理外,SOD 活性最大值均在4 月23 日出现,也就是干旱程度最大的时候,T4 处理SOD 活性最大值出现在5 月3 日,较其他处理延迟了10 d 出现。控水第1天(4月3日),SOD活性各处理与对照之间差异不显著。控水第10 天(4 月13 日),T1 处理与CK 差异不显著;T2、T3 处理进入轻旱状态,SOD 活性较CK 升高30%~40%,且与T1、CK处理差异显著;T4 处理进入中旱状态,SOD 活性比CK 高58%,与CK、T1、T2、T3 处理差异显著。控水第20 天(4 月23 日),T1 处理进入中旱状态,SOD 活性较CK 升高20%,且差异显著;T2、T3 处理进入重旱状态,SOD 活性较CK 升高30%~38%,且差异显著;T4 处理进入特旱状态,SOD 活性比CK高28%,与CK差异显著,与T1、T2、T3处理差异不显著。复水第7 天(5 月3 日),T1、T2、T3 处理SOD 活性较CK 高15%~26%,且差异显著;T4 处理SOD 活性较CK 高44%,且差异显著。复水第17 天(5 月13 日),T1、T2、T3 处理SOD活性与CK 差异不显著,T4 处理与CK 差异显著。由图2 可知,复水后,SOD 活性呈现下降的趋势,T1、T2、T3 下降幅度较大,而T4 处理下降幅度较小。POD 活性变化规律与SOD 活性变化规律基本一致。CAT 活性在控水期变化规律与SOD 活性变化规律基本一致,复水后,CAT 活性较控水时先升高后降低,最大值出现在复水第7天,说明CAT活性对水分变化的敏感程度较SOD、POD 要低。MDA 含量随时间的延长呈现持续升高的趋势,最大值均出现在复水第17 天,除了控水第1 天外,其余时间T4处理与其他处理之间差异显著。
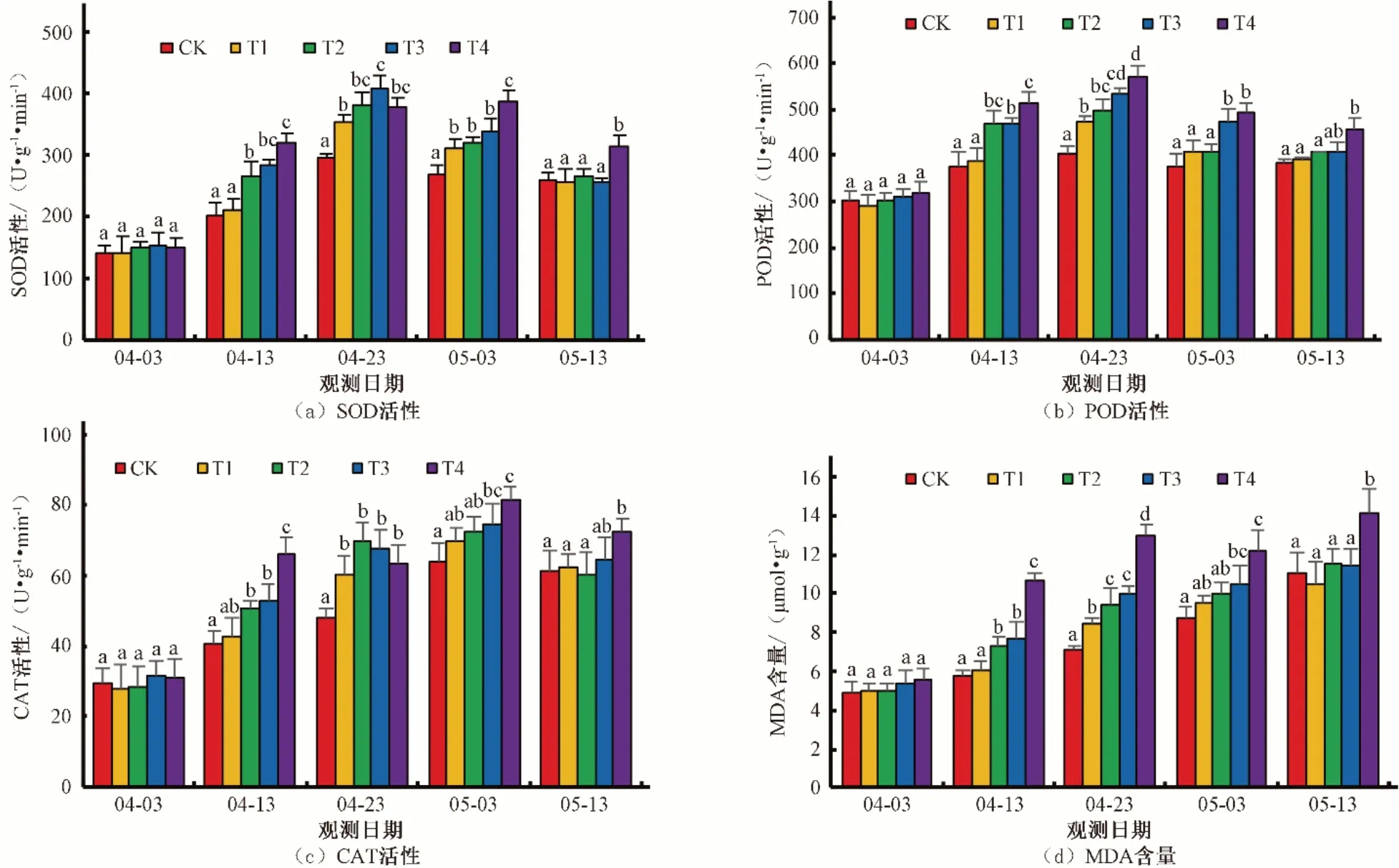
图2 不同干旱处理条件下冬小麦抗氧化酶活性及MDA含量的变化Fig.2 Changes of antioxidant enzyme activity and MDA content of winter wheat under different drought treatment conditions
由图2 可知,SOD、POD 活性在复水后均呈下降的趋势,CAT 活性小幅升高而后下降,在复水第17 天,T1、T2、T3 均能下降到与CK一致的水平,差异不显著;由于T4处理一致处于干旱胁迫状态,且程度较重,导致复水17 天后仍与其他处理及CK有较大差距,且差异显著。MDA含量随时间的延长呈现升高趋势,复水后仍保持持续升高的趋势,T4 处理在复水后有小幅回落,而后继续升高,说明复水对MDA 含量变化影响不大。
2.3 干旱后复水对冬小麦叶片光合特性的影响
由图3 可知,随着时间的延长,对照组(CK)的Pn、Tr、Gs 和Ls 均呈现先升高后降低的趋势,最大值均出现在控水第20天,Ci呈先降低后升高的趋势,最小值出现在控水第20天,WUE 持续降低。不同处理的Pn 值变化规律与CK 不同,由于T1、T2 处理受干旱胁迫程度较轻,在控水期呈现先升高后降低的趋势,控水第10 天较第1 天有小幅升高,而后又降低,复水后持续降低;T3、T4 处理受干旱胁迫程度较重,控水期持续降低,复水后有小幅升高,而后降低。控水第1天各处理Pn 值基本相同,差异不显著;控水第10 天,T1 处理与CK 差异不显著,T2、T3、T4 处理较CK 降低9%、15%、21%,且差异显著;控水第20 天,T1、T2、T3 处理较CK 降低15%、33%、33%,且差异显著,T4 处理较CK 降低46%,且与其他处理均差异显著;复水第7 天,T1 处理与CK 差异不显著,T2、T3 处理较CK 降低17%、16%,且差异显著,T4 处理较CK 降低28%,且与其他处理均差异显著;复水第17 天,T1、T2、T3 处理与CK 差异不显著,T4 处理较CK 降低18%,且与其他处理均差异显著。各处理Tr、Gs 和Ls 值变化规律与Pn 变化规律基本一致,Ls 值变化幅度较其他值略小。不同处理的Ci 值变化规律与CK 基本相同,控水第10 天,T1 处理与CK 差异不显著,T2、T3、T4 处理较CK 降低7%、7%、14%,且差异显著;控水第20 天,T1、T4 处理与CK 差异不显著,T2、T3处理较CK降低16%、19%,且差异显著;复水第7天,T1、T2、T3 处理与CK 差异不显著,T4 处理较CK 升高9%,且差异显著;复水第17 天,T1、T2、T3 处理与CK 差异不显著,T4处理较CK 升高15%,且与其他处理均差异显著。不同处理的WUE 值变化规律与CK 基本相同,控水第10 天,T1、T2、T3、处理与CK 差异不显著,T4 处理较CK 升高18%,且差异显著;控水第20 天,T2 处理与CK 差异不显著,T1、T3、T4处理较CK 升高14%、15%、13%,且差异显著;复水第7 天和第17 天,T1、T2、T3 处理与CK 差异均不显著,T4 处理较CK升高18%、53%,且与其他处理均差异显著。
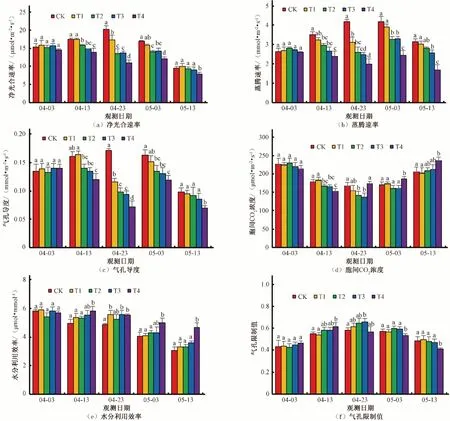
图3 不同干旱处理条件下冬小麦叶片光合参数的变化Fig.3 Changes of photosynthetic parameters of winter wheat Leaves under different drought treatment conditions
由图3 可知,干旱处理的Pn、Tr、Gs 和Ls 值在复水后有呈升高趋势,而后下降,在复水第17 天,T1、T2、T3 均能恢复到与CK 一致的水平,差异不显著;而干旱胁迫程度较重的T4 处理,复水17 天后仍与其他处理及CK 有较大差距,且差异显著。各处理Ci 值在复水后均呈升高的趋势,WUE 值呈缓慢下降趋势,且T1、T2、T3 处理复水后与CK 差异不显著,而T4处理显著高于其他处理。
3 讨论与分析
植物体内正常的代谢会产生活性氧,过量的活性氧会对植物体产生伤害。由SOD、POD、CAT等抗氧化酶组成的酶促系统可有效地清除多余的活性氧[19],避免引起细胞膜脂过氧化,保护植物细胞。干旱胁迫可以降低作物光合器官对光能的转化和利用效率,使作物同化能力下降,产生大量的活性氧,这时候作物会启动应对机制以减小干旱带来的影响[20]。本研究表明,干旱胁迫可以使植物体内SOD、POD、CAT等抗氧化酶活性迅速升高,MDA 含量也相应的增加,干旱胁迫程度越重,升高幅度越大,但当干旱胁迫时间长、程度过重时(控水第20 天的T4 处理),SOD、CAT 活性相比呈下降趋势,而POD 活性仍呈升高的趋势,说明冬小麦体内的酶促系统紊乱,细胞膜脂过氧化已不能被及时修复,这一点也可从MDA含量迅速升高看出。由于POD 酶具有吲哚乙酸氧化酶的性质,POD 活性在达到某一阈值前,主要表现为对活性氧的消除,而达到后则主要促进叶绿素的降解[21],因此,SOD、CAT 活性的下降和POD 活性的上升均不利于活性氧的清除,加剧冬小麦叶片细胞的破坏,从而影响叶片的光合作用。岳虹、张军的研究结果均表明,小麦在受到干旱胁迫时,SOD、POD、CAT 等抗氧化酶活性增加,脂质过氧化水平提高,MDA 含量随之增加[8,22],这与本研究结果一致。复水后,中旱和重旱胁迫条件下,冬小麦叶片内SOD、POD、CAT 活性和MDA 含量均能恢复至正常水平,而特旱胁迫条件下,显著高于其他处理,说明干旱胁迫对冬小麦叶片造成严重损伤,甚至不可逆的损伤,即使复水后,细胞修复过程也缓慢或者无法修复。王利彬的研究发现,干旱胁迫后复水产生补偿效应是有限的,复水后可使SOD 活性缓慢降低,SOD 活性仍保持较高的修复细胞膜结构的水平,但长时间重度胁迫会使植物抗氧化调节能力降低,补偿效应降低甚至产生伤害效应[23]。柳燕兰的研究表明,干旱胁迫条件下春玉米叶片SOD、POD、CAT等抗氧化酶活性均显著增加,复水后,酶活性有所下降,但仍高于正常灌溉处理,且重度干旱处理下降幅度较小[9]。王梦娇的研究表明也表明,不同品种的冬小麦在受到干旱胁迫时,SOD 活性和MDA 含量随干旱程度的增加而逐渐升高,复水后SOD 活性和MDA 含量降低,但仍高于对照,胁迫程度越深,恢复时间越长[24]。赵龙等对夏玉米拔节期进行不同程度和时长的干旱胁迫处理,结果表明,随胁迫程度加剧,MDA、SOD、POD 酶活性和CAT 含量呈升高趋势[25]。这些研究结果均与本研究结果一致。
干旱胁迫影响植物体细胞活性的同时,作物的自我保护机制会使气孔导度和蒸腾速率降低,减少体内水分的散失,降低作物的净光合作用能力[26]。冬小麦在受到干旱胁迫后,Pn、Tr、Gs 和Ci 值呈下降趋势,且下降幅度与受到的胁迫程度有关,干旱胁迫程度越重下降越大,重度干旱胁迫可使净光合速率降低50%以上[12,27,28]。光合作用的降低通常由气孔因素和非气孔因素造成,气孔因素导致的光合作用降低主要是指冬小麦受到干旱胁迫时Gs 降低导致Ci 值降低,非气孔因素导致的光合作用降低主要是指作物光合器官的光合活性和叶绿素含量下降等因素[13]。气孔因素和非气孔因素主要从Pn、Ci 和Ls 值的变化区分,当Pn 和Ci 值都减少,而Ls 值增大时,就认为是气孔因素导致光合作用降低;当Pn 值减小,Ci 值增大,而Ls 值减小,就认为是非气孔因素导致光合作用降低[29]。本研究中,控水第20 天的T4 处理可认为是非气孔因素造成的光合作用降低,其他处理可认为是由气孔因素造成的光合作用降低。李佳佳的研究表明,干旱胁迫条件下叶片光合速率降低是由气孔因素导致的,而在严重胁迫条件下,非气孔因素起主要作用[30],这与本研究一致,T4 处理受到的干旱胁迫程度相比其他处理要严重得多。对于叶片WUE 值,本研究表明干旱胁迫条件下,WUE 值呈下降的趋势。上官周平的研究表明,干旱条件下冬小麦叶片WUE 值比灌溉充分条件下低13%~26%[31];张雅倩等的研究表明,冬小麦叶片WUE 值随干旱胁迫程度的加重呈下降趋势[32]。这与本研究结果一致,但不同的研究呈现不同的结果。袁蕊的研究表明干旱胁迫使小麦在灌浆期的水分利用效率增加了50.66%[33]。这是由于叶片WUE 值是Pn 与Tr 的比值,与Pn 呈正相关,与Tr 呈负相关,蒸腾作用主要取决于气象条件,而光合作用除受环境因素的影响外,还与其他诸多因素有关,适度的干旱可提高叶片WUE 值[34]。另外,叶片WUE 值还与品种有关,干旱胁迫条件下抗旱或旱肥型品种叶片WUE 值高于其他类型的品种[35]。复水后,干旱胁迫条件下的冬小麦产生补偿效应,Pn、Tr、Gs和Ci 值均有升高趋势,干旱胁迫较轻的处理能恢复到与正常灌水条件相当的水平,而干旱胁迫较重的处理,恢复较为缓慢或恢复时间延长。周雪英的研究表明,与干旱处理相比,拔节期恢复供水可使受旱小麦叶片的Pn 和Tr 显著提高且高于正常对照水平[10]。李彦彬的研究表明,复水后干旱处理的Pn、Tr和Ci值均有所恢复,轻旱处理的Pn和Gs可以恢复到正常水平,而中旱和重旱却仍距正常水平有较大差异[36]。
4 结 论
在一定程度干旱胁迫条件下,冬小麦SOD、POD、CAT活性和MDA 含量显著升高,光合速率和水分利用效率下降,复水后产生补偿效应。而当干旱胁迫达到特旱条件时,SOD、CAT 活性反而有所降低,Ci 值有所升高,这说明特旱条件下,叶片细胞遭到损伤,已不能进行正常的生理生化反应和光合作用,即使复水,相较于干旱胁迫较轻的处理,也不能产生足够的补偿效应,以恢复叶片正常的生理生化反应和光合作用。所以,极端的干旱条件,会对冬小麦产生不可逆的损伤,造成小麦产量下降,在实际农业生产活动中,要避免麦田出现极端的干旱条件,才能保证小麦的稳产、高产。
