调控TGF-β1-PI3K/AKT轴对骨肉瘤细胞恶性增殖和干性表达的影响
2023-10-25马琨张川
马琨 张川
1河南省洛阳正骨医院(河南省骨科医院)病理科,郑州 450002;2河南省洛阳正骨医院(河南省骨科医院)肩肘二科,郑州 450002
骨肉瘤是最常见的骨原发恶性肿瘤,好发于青少年的股骨远端、胫骨近端和肱骨近端等长骨干骺端[1]。虽然“术前新辅助化疗”治疗模式已经把骨肉瘤患者的5年生存率提高到了60%~70%,但近数十年来,骨肉瘤治疗效果停滞不前[2-3]。近年来研究证实肿瘤干细胞(cancer stem cells,CSCs)在肿瘤复发和化疗耐药中发挥重要作用[4-5]。既往研究表明,转化生长因子-β1(transforming growth factor-β1,TGF-β1)可参与调控成骨细胞自噬,诱导干细胞、心肌细胞纤维化,且异常增高的TGF-β1水平与多种肿瘤的临床分期和不良预后呈正相关[6-10]。虽目前已有报道认为TGF-β1参与抑制骨肉瘤细胞增殖和侵袭[11]。然而,关于TGF-β1与骨肉瘤恶性增殖和肿瘤干细胞特征之间的关系尚无人探讨。本研究旨在了解TGF-β1与骨肉瘤细胞恶性增殖和肿瘤干细胞特征的关系,为骨肉瘤治疗靶点提供新选择。
材料与方法
1.细胞与试剂
人骨肉瘤细胞系U2OS、MG-63、SaoS2、HOS和人正常成骨细胞hFOB1.19均购自美国ATCC细胞库,用含有10%胎牛血清的DMEM培养。TGF-β1抗体购自英国Biorbyt公司;基因敲减和过表达TGF-β1的重组慢病毒和阴性对照慢病毒均购自上海吉玛生物科技有限责任公司;一抗[抗TGF-β1、抗E-钙黏蛋白(E-cadherin)、抗波形蛋白(vimentin)、抗磷酸甘油醛脱氢酶(GAPDH)、抗细胞表面粘附分子 (CD133)、抗八聚体结合转录因子(OCT)4、抗Nanog、抗p-磷脂酰肌醇3-激酶(PI3K)、抗p-蛋白激酶B(AKT)、抗半胱氨酸天冬氨酸蛋白酶3(caspase-3)、抗α-平滑肌肌动蛋白抗体(α-SMA)、抗性别决定相关基因簇-2抗体(Sox-2)]、二抗均购自美国Santa Cruz公司。反转录聚合酶链反应(qRT-PCR)和反转录试剂盒购自日本TaKaRa公司。
2.细胞培养
将细胞放置在含有10%胎牛血清、1%青霉素、1%链霉素的DMEM/F12,在37 ℃、5% CO2饱和度的细胞培养箱中培养。待细胞生长至70%左右时,采用0.25%的胰蛋白酶进行消化传代。
3.细胞增殖能力检测
用四甲基偶氮唑盐(MTT)实验检测骨肉瘤细胞增殖能力,将经过不同因素处理后的MG-63细胞消化、计数,以密度为1×104ml-1接种于96孔板,然后在37 ℃、5% CO2浓度培养箱孵育,使用酶标仪在450 nm波长下测量。
4.腺病毒与小干扰RNA转染设计
常规种植MG-63细胞到6孔板内,37 ℃、5% CO2的培养箱中过夜。根据Lipofectamine 2000转染说明书,将相应的质粒转染至细胞中。转染24 h后更换新鲜的培养液继续培养,嘌呤霉素筛选。以稳定感染过表达TGF-β1重组慢病毒的MG-63细胞为Lv-TGF-β1组,以稳定感染阴性对照慢病毒的MG-63细胞为Lv-con组。TGF-β1 siRNA序列:#1正向引物5'-CCCACAACGAAAUCUAUGATT-3',反向引物5'-UCAUAGAUUUCGUUGUGGGTT-3';#2正向引物5'-GACACCAACUAUUGCUUCATT-3',反向引物5'-UGAAGCAAUAGUUGGUGUCTT-3';#3正向引物5'-GUCAACUGUGGAGCAACACDT-3',反向引物5'-GUGUUGCUCCACAGUUGACDTDT-3'。干扰阴性对照组:正向引物5'-UUCUCCGAACGUGUCACGUTT-3',反向引物5'-ACGUGACACGUUCGGAGAATT-3'。
5.实时荧光定量PCR
利用Trizol试剂提取细胞RNA,按反转录试剂盒说明书将RNA逆转录为cDNA。Real-time PCR反应体系及条件参照试剂盒说明书进行操作,2-△△CT法计算mRNA的相对表达量,以GAPDH作为内参。每组实验重复3次。TGF-β1正向引物:5'-TCTCCAGGCATTTCCACTATTC-3',反向引物:5'-CTCAGGCATTCGTCA ACATCTA-3';GAPDH正向引物:5'-GCACCGTCAAGGCTGAGAAC-3',反向引物:5'-TGGTGAAGACGCCAGTGGA-3'。
6.流式细胞术测定细胞凋亡
采用FITC-annexinⅤ/PI双染法,分别收集各处理组细胞到离心管中,1 000 r/min离心5 min(离心半径8 cm)。室温下避光孵育15 min,沉淀细胞孵育缓冲液洗1次。加入FITC-annexinⅤ/PI溶液5 µl 4 ℃下孵育20 min,以流式细胞仪检测细胞凋亡情况。
7.悬浮成球实验
将细胞分散接种于超低吸附六孔板中,使用无血清DMEM/F12培养基,其中添加20 µg/L人表皮生长因子(EGF)、20 µg/L人碱性成纤维细胞生长因子(bFGF)、5 µg/L胰岛素、2% B-27。培养1~2周后,显微镜下观察并计算肿瘤球数目。
8.流式细胞术检测肿瘤干细胞表面标志物
待各处理组细胞生长至对数期时,0.25%胰蛋白酶消化,调整为1×104ml-1的单细胞悬液。按照说明书每组分别加入1∶1 500的CD133抗体1 ml,充分混匀后常温孵育,用流式细胞仪检测。
9.蛋白免疫印迹(Western blotting)
收集各组细胞,添加RIPA细胞裂解液,4 ℃,3000 r/min(离心半径15 cm)离心30 min;经BCA法定量以后,取适量的总蛋白煮沸变性。以100 g/L的分离胶和50 g/L的浓缩胶进行电泳,每孔上样30 µg蛋白样品。把凝胶放在缓冲液中室温检测显影,凝胶成像系统拍照。浸泡25 min,以100 V的电压转膜。转膜结束后,将PVDF膜放在脱脂奶粉中室温封闭,加入特异性一抗TGF-β、E-cadherin、Vimentin、GAPDH、CD133、OCT-4、Nanog、p-PI3K、p-AKT、caspase-3、α-SMA、Sox-2室温反应1.5 h,4 ℃孵育过夜;加二抗(1∶3 000稀释)室温孵育1 h,用ECL发光检测显影,凝胶成像系统拍照。
10.移植瘤实验(体内研究):动物模型建立、分组和给药
BALB/c裸鼠20只,日龄28~35 d,体质量18~22 g,常规裸鼠饲养。实验分为4组:Lv-con、Lv-TGF-β1、si-con、si-TGF-β1组;调整细胞悬液浓度为1×105ml-1。每组取0.1 ml分别接种于裸鼠左或右侧腋窝皮下,每天观察小鼠一般情况,隔日测量1次移植瘤体积。在骨肉瘤细胞种植后5 d,将动物随机分4组,每组5只,对照组只给予生理盐水;实验组采用预先实验方案给予处理,隔天腹腔内注射。饲养至27 d,处死动物,完整取出移植瘤称重,立即行超低温储藏。
11.统计学处理
采用SPSS 21.0软件分析,符合正态分布的计量资料采用()表示,各组间比较采用独立样本t检验,以P<0.05为差异有统计学意义。
结果
1.TGF-β1在骨肉瘤细胞系中高表达(图1)

图1 TGF-β1在骨肉瘤细胞系中的表达。A:qRT-PCR检测骨肉瘤各细胞系的TGF-β1 mRNA水平;B、C:Western blotting检测骨肉瘤各细胞系的TGF-β1蛋白水平
与正常人成骨hFOB1.19细胞比较,骨肉瘤U2OS、MG-63、HOS及SaoS2细胞系中的TGF-β1 mRNA和蛋白印迹水平表达均有不同程度的上调;在U2OS细胞系中上调最显著(t=8.76,P<0.05),在MG-63细胞系中增长程度最低(t=4.33,P<0.05)。因此,后续实验中U2OS细胞用于TGF-β1沉默和MG-63细胞用于TGF-β1过表达实验。
2.TGF-β1 siRNA转染骨肉瘤细胞的效率(图2)
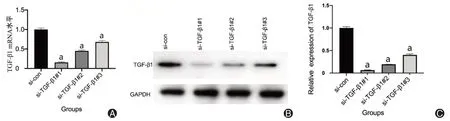
图2 qRT-PCR和Western blotting检测si-TGF-β1沉默效率。A:qRT-PCR检测各组mRNA水平;B、C:Western blotting检测TGF-β1沉默后各组蛋白水平
本研究中,3组siRNA瞬时转染U2OS细胞后,qRT-PCR和Western blotting用于检测TGF-β1基因的表达情况。结果显示,与对照组相比,3组si-TGF-β1均不同程度降低了TGF-β1的mRNA和蛋白表达水平,其中siRNA#1干扰效果最明显(t=10.88,P<0.05)。因此,选用siRNA#1用于后续实验。
3.调控TGF-β1影响骨肉瘤细胞增殖和凋亡(图3)
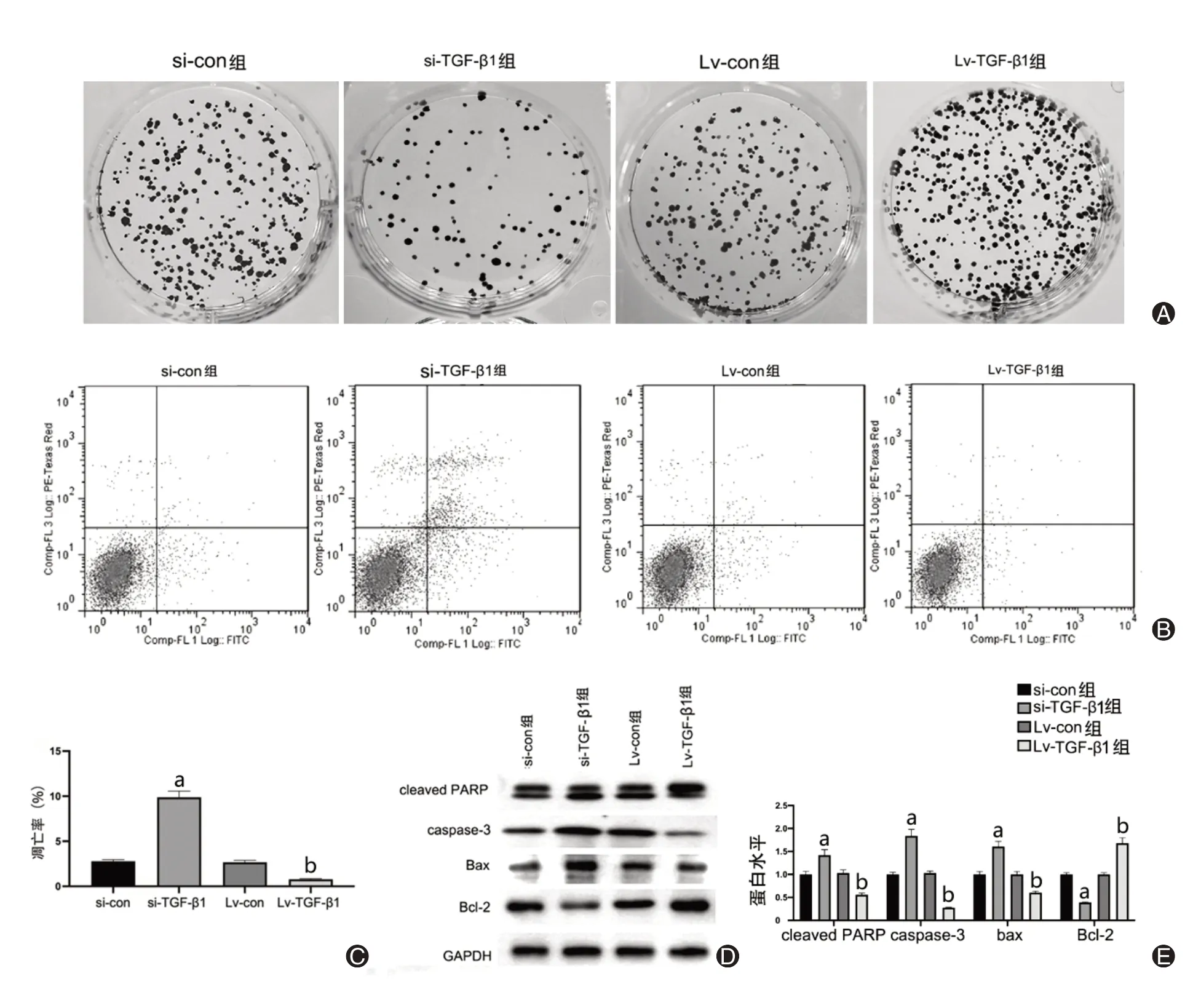
图3 调控TGF-β1对骨肉瘤细胞增殖和凋亡情况的影响。A:克隆平板实验检测si-TGF-β1转染后U2OS/MG-63细胞的增殖能力;B、C:流式细胞术检测si-TGF-β1转染后U2OS/MG-63细胞的凋亡率;D、E:Western blotting检测cleaved PARP、caspase-3、Bax、Bcl-2蛋白水平
克隆形成实验结果展示,si-TGF-β1能抑制MG-63细胞的平板克隆数;过表达TGF-β1能上调MG-63细胞的增殖能力(t=3.28,P<0.05)。同时流式细胞术被用于检测U2OS/MG-63细胞凋亡情况。数据表明,si-con组与Lv-con组之间差异无统计学意义(P>0.05);相对于si-con组,si-TGF-β1的细胞凋亡率显著升高,而与Lv-con相比,Lv-TGF-β1的细胞凋亡率有所下降(t=5.11,P<0.05)。并且我们从Western blotting实验的结果进一步证实,与si-con比较,si-TGF-β1组的cleaved PARP、caspase-3、Bax表达明显上调,而Bcl-2水平下降;反之与Lv-con相比,Lv-TGF-β1组的cleaved PARP、caspase-3以及Bax水平有所下降,而Bcl-2表达上调。上述实验结果表明,si-TGF-β1可促进骨肉瘤细胞的凋亡。
4.TGF-β1调控骨肉瘤干细胞样特性表达(图4)
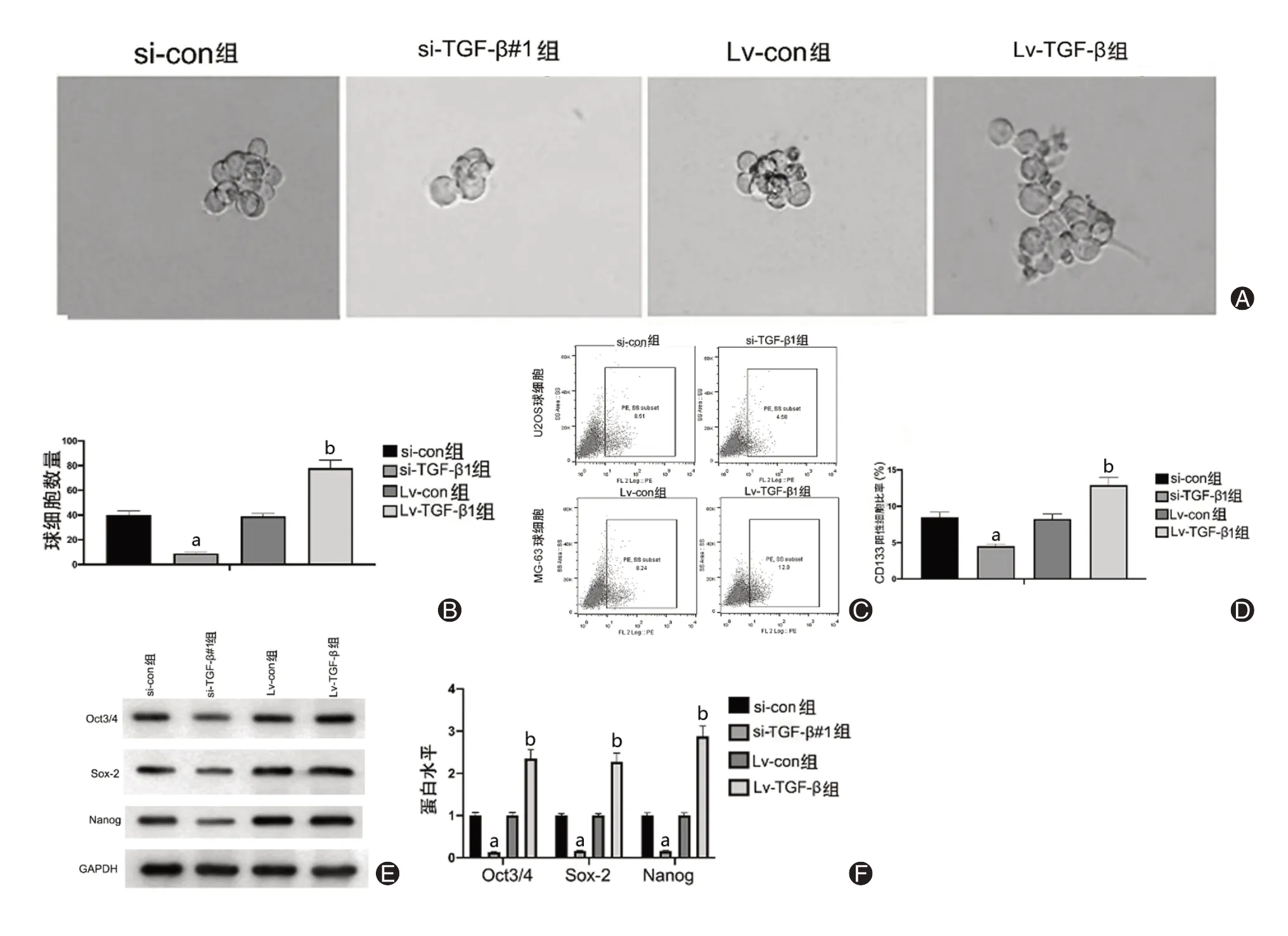
图4 TGF-β1调控骨肉瘤干细胞表型特征变化。A、B:调控TGF-β1对骨肉瘤干细胞球形成的影响;C、D:调控TGF-β1对骨肉瘤干细胞球CD133阳性细胞比例的影响;E、F:调控TGF-β1对骨肉瘤干细胞球表面标志物表达水平的影响
肿瘤成球实验显示,相较于对照组,si-TGF-β1组肿瘤球的长径明显缩小(t=6.88,P<0.05);且si-TGF-β1组肿瘤球克隆数量明显减少(t=3.07,P<0.05)。CD133阳性细胞比例是肿瘤干细胞的特征;图中显示,与干细胞球形成实验的结果趋势相同,基因沉默TGF-β1使CD133阳性细胞比例下降,即为降低骨肉瘤干细胞的干性特征表达;过表达TGF-β1使CD133阳性细胞比例上升,即为增高骨肉瘤干细胞的干性特征表达。Westen blotting检测结果表明,与si-con组比较,si-TGF-β1组的干细胞表面特征标志物Sox-2、Oct3/4、Nanog的蛋白表达显著降低;与Lv-con组比较,TGF-β1过表达上调Sox-2、Oct3/4、Nanog的蛋白水平。进而证实了影响TGF-β1水平可调控骨肉瘤的干细胞样特性。
5.TGF-β1通过PI3K/AKT信号通路调控骨肉瘤细胞的恶性增殖和干性表达(图5)

图5 Western blotting检测调控TGF-β1后p-P13K和p-AKT蛋白表达的变化
Western blotting结果显示,si-con组与Lv-con之间无明显差异;与si-con组相比,抑制TGF-β1的表达可抑制p-PI3K、p-AKT表达;而与Lv-con组相比,过表达TGF-β1的骨肉瘤细胞中p-PI3K、p-AKT表达水平升高(t=4.12,P<0.05)。
6.TGF-β1对裸鼠皮下移植瘤增殖能力的影响(图6)

图6 裸鼠体内成瘤试验。A:TGF-β1调控形成的异种移植瘤图像;B、C:异种移植瘤的体积和质量
构建裸鼠皮下移植瘤,在体内水平检测TGF-β1对骨肉瘤细胞增殖能力的影响,结果表明,si-con组与Lv-con组的肿瘤相比,体积和质量的差别差异无统计学意义(均P>0.05)。si-TGF-β1组的肿瘤体积和质量明显低于si-con组(t=3.79,P<0.05);同理,Lv-TGF-β1的肿瘤体积和质量显著高于Lv-con组(t=5.01,P<0.05)。提示TGF-β1可促进裸鼠体内骨肉瘤细胞的增殖。
讨论
骨肉瘤是青少年高发的骨原发性恶性肿瘤之一。虽然骨肉瘤手术、放疗、生物治疗等方式日趋完善,但部分患者由于远处转移和复发,预后较差。因此,探讨复发转移发生机制、探寻相应干预措施对于改善骨肉瘤预后具有重要意义。
在本研究中,我们首次发现TGF-β1在骨肉瘤细胞系中均有不同程度的高表达。有文献报道TGF-β1相关通路参与结肠癌/肝癌细胞凋亡[12-13]。Fig2A显示与si-con组相比,si-TGF-β1组平板克隆数明显减少,U2OS细胞凋亡百分比增加到9.91%。反之,Lv-TGF-β1组明显增加克隆平板数,MG-63细胞的凋亡数减少到0.85%(P<0.05)。这提示我们过表达TGF-β1水平有助于骨肉瘤增殖,抑制其凋亡。
TGF-β1是超家族多肽类细胞因子成员[14]。Hariyanto等[15]发现,外源性TGF-β1可增强乳腺癌肿瘤干细胞的OCT4和ALDH1A1 mRNA的表达。Leng等[16]也发现TGF-β1相关通路参与KLF4增强结肠癌Lgr5 CD44 EpCAM阳性细胞比例。与既往研究结果类似,我们亦证实TGF-β1有助于增强骨肉瘤细胞的干性表达:与si-con组相比,si-TGF-β1可缩小MG-63细胞球的直径和数量(P<0.05),下调CD133阳性细胞百分比,Oct-4、Sox-2、Nanog、CD133的蛋白表达水平。经腺病毒TGF-β1过表达后,MG-63球囊的直径和细胞个数增加(P<0.05),上调CD133阳性细胞百分比,上调干性标志物的表达水平。
PI3K/AKT通路是介导多种肿瘤细胞增殖和转移的“明星通路”之一[17-18]。有报道研究证实,TGF-β1通过调控PI3K/AKT通路参与乳腺癌、肝癌、胃癌上皮间质转化[19-20]。但至今未有文献报道TGF-β1可通过PI3K/AKT通路调控肿瘤干细胞的干性特征。在本实验中,与si-con相比,下调TGF-β1水平的p-AKT和p-PI3K蛋白表达水平随之下调;反之,过表达TGF-β1组的p-AKT和p-PI3K表达量上调;与申力和赵立志[21]报道TGF-β1组的PI3K/p-AKT/mTOR蛋白表达量高于NC组一致。因此,本研究中我们首次发现PI3K/AKT通路可能参与TGF-β1介导的骨肉瘤恶性生物学行为。且我们从裸鼠移植瘤体内实验进一步证实,与体外试验结论相同,下调TGF-β1的表达可减少肿瘤的体积和质量(P<0.05);相反过表达TGF-β1的表达可增高肿瘤的体积和质量。
综上所述,调控TGF-β1的表达可影响骨肉瘤细胞的恶性增殖和干性特征;这一切是通过激活PI3K/AKT通路进而实现的。本研究发现了TGF-β1具有作为骨肉瘤治疗潜在靶点的价值,但需要进一步的体内及体外实验加以验证其作用及促癌机制,为骨肉瘤的复发、耐药及靶向治疗提供了一定的理论基础。
利益冲突所有作者均声明不存在利益冲突
作者贡献声明马琨:酝酿和设计试验,实施研究,采集、分析/解释数据,文章撰写,统计分析,获取研究经费,行政、技术或材料支持;张川:指导,支持性贡献
