药隔期倒春寒对小麦颖果发育的影响*
2023-10-20王捧娜代雯慈蔡洪梅郑宝强李金才
翁 颖,王捧娜,于 敏,代雯慈,吴 宇,蔡洪梅,郑宝强,李金才,2**,陈 翔**
药隔期倒春寒对小麦颖果发育的影响*
翁 颖1,王捧娜1,于 敏1,代雯慈1,吴 宇1,蔡洪梅1,郑宝强1,李金才1,2**,陈 翔1**
(1.安徽农业大学农学院/农业部华东地区作物栽培科学观测站,合肥 230036;2.江苏省现代作物生产协同创新中心,南京 210095)
以抗倒春寒性弱的小麦品种新麦26(XM26)和抗倒春寒性强的烟农19(YN19)为试验材料,进行大田盆栽试验。于小麦幼穗分化的药隔形成期在智能超低温光照培养箱内进行低温(2℃和−2℃)胁迫处理,以10℃作为对照组(CK)。于花后每隔7d进行取样,对小麦强势和弱势颖果的长度、宽度和厚度进行观测分析,同时采用石蜡切片技术观察果皮、胚乳细胞及大、小淀粉粒的显微结构,以探明药隔期倒春寒胁迫下小麦颖果发育的显微结构特征。结果表明:(1)药隔期倒春寒会降低小麦颖果的长度、宽度和厚度,且降幅随倒春寒胁迫程度的加强(2℃→−2℃)而增大,成熟期小麦颖果长度、宽度和厚度分别减小1.17%~4.41%、1.57%~10.22%和1.42%~9.40%,倒春寒胁迫下成熟期两品种小麦各处理弱势粒颖果长/宽、长/厚、宽/厚比值均高于强势粒颖果。(2)药隔期倒春寒会减缓早期颖果果皮降解速率,同时还会抑制两品种小麦颖果胚乳细胞淀粉粒的发育。(3)药隔期倒春寒降低小麦强、弱势颖果胚乳细胞内大、小淀粉粒的周长和面积,花后28d强势粒颖果胚乳细胞内大、小淀粉粒周长和面积分别下降8.17%~14.66%和14.08%~17.98%、0.94%~5.00%和2.59%~10.03%,弱势粒颖果分别下降3.08%~10.31%和6.56%~8.70%、3.17%~6.39%和11.85%~16.17%,且强势粒颖果内大淀粉粒周长和面积下降较多,而弱势粒颖果内小淀粉粒周长和面积下降较多。综上所述,药隔期倒春寒通过减缓小麦颖果发育早期果皮降解,抑制胚乳细胞淀粉粒的发育,降低胚乳细胞内大、小淀粉粒的周长和面积,从而降低小麦颖果的长度、宽度和厚度,导致小麦籽粒库容降低,最终降低小麦粒重。
小麦;倒春寒;强弱势;颖果;淀粉粒
小麦(L)籽粒营养价值高,含有蛋白质、脂肪、淀粉、维生素等营养物质,是主要口粮作物[1−2]。随着城市化的发展,对小麦的需求量也随之持续增加。近10a来,中国小麦总产量从2012年的1.22×108t增至2021年的1.37×108t,年平均增长率为1.17%;对小麦的需求量也从2012年的1.28×108t增至2021年的1.52×108t,年均增长率为1.73%(http://www.agdata.cn/)。因此,小麦生产的高产高质高效发展对保障国家粮食安全和促进国民经济发展均具有极其重要的意义。
小麦在不同生育时期对温度的要求不尽相同,若在敏感期遭遇低温胁迫,则会对其生长发育及产量品质产生不利影响[3]。近年来,全球气候变化造成大气环流异常,导致极端低温气候灾害事件的发生频率、强度和持续时间不断增加[4−6]。倒春寒主要指小麦在拔节−孕穗期遭遇突然降温天气,造成幼穗受伤或死亡,部分小穗不结实甚至全穗不结实,从而导致小麦减产的一种农业气象灾害[7]。作为中国最大的小麦主产区,黄淮麦区倒春寒灾害的年际发生频率更是达到了40%[8]。一般轻中度倒春寒危害后小麦减产10%~30%,重度倒春寒危害后小麦减产幅度可达50%以上[3],其已成为限制小麦生产高产高质高效发展的重要因素之一。
前人关于倒春寒胁迫对小麦的光合特性[9−13]、ROS代谢[14]、蔗糖代谢[15−17]、籽粒灌浆[18]和基因的差异表达[19]等生理生化过程做了大量研究。刘蕾蕾等[12]研究发现倒春寒会降低小麦叶片最大光化学效率和净光合速率,从而降低光合同化有机物的积累。柯媛媛等[10]研究表明药隔期倒春寒通过降低小麦花前干物质的积累量、转运量、转运效率以及花前贮藏干物质对籽粒的贡献率,从而减少光合同化有机物向小麦穗部的转运和分配。Zhang等[17]研究发现,孕穗期低温通过诱导小麦穗部脱落酸积累,改变了蔗糖代谢关键酶活性,从而影响幼穗的正常发育,造成穗粒数的减少。孕穗期低温还会降低小麦籽粒灌浆过程中ADP−葡萄糖焦磷酸化酶(AGPase)、可溶性淀粉合成酶(SSS)、颗粒结合淀粉合成酶(GBSS)等关键酶活性,进而降低灌浆速率,最终造成粒重降低[18]。此外,Jiang等[19]对药隔期不同程度倒春寒胁迫下的小麦幼穗进行转录组测序,其中−4℃处理下从敏感型小麦发现1353个差异表达基因(Differential expressed gene,DEG),从抗倒春寒性强的小麦品种发现1882个DEGs,经生物信息学分析发现这些DEGs主要参与蔗糖和淀粉代谢、激素信号转导和转录因子调控等过程。但鲜有研究报道倒春寒灾害对小麦颖果发育,尤其是强、弱势颖果发育的影响。小麦穗部籽粒的位置决定了籽粒灌浆进程与干物质积累的空间分布特征,因此,强弱势籽粒对养分的供应存在竞争关系,且在粒重、蛋白质含量等方面存在差异性[20]。本研究以抗倒春寒性强弱不同的小麦品种烟农19和新麦26为材料,在小麦幼穗分化的药隔形成期(低温敏感期[10])模拟大田倒春寒灾害的温变规律进行低温胁迫处理,从颖果形态和解剖生理等方面探究倒春寒灾害对小麦强弱势颖果发育的影响,以期为小麦生产的防灾减灾和高产高质高效发展提供理论依据。
1 材料与方法
1.1 试验材料
在前期研究的基础上[21],选用抗倒春寒性弱的小麦品种新麦26(XM26)和抗倒春寒性强的小麦品种烟农19(YN19)为供试材料。试验于2021年11月1日−2022年5月18日在安徽农业大学农萃园基地(31°52′01″N,117°16′58″E,海拔21.3m)进行。该地区属于北亚热带湿润季风气候,年平均气温、降水量、日照时数和无霜期分别为15℃、875mm、2100h和230d[10]。供试土壤类型为黄棕壤,0−20cm土层含有机质16.3g·kg−1,pH值为6.5,速效氮、速效磷和速效钾含量分别为112.2、23.0和161.6mg·kg−1。
1.2 试验设计
采用大田盆栽方式进行种植,盆栽土壤取自于农萃园土壤耕层(0−20cm土层)。PVC盆栽桶直径为25cm,高度为30cm,每盆装土10kg。播种日期为2021年11月1日,盆栽桶内土壤经水沉实后播种,每盆播种18粒种子,三叶期定苗,最终留苗9株。播种前每盆施6.67g氮磷钾复合肥(N:P2O5:K2O= 15:15:15)和0.44g尿素(N=46.4%),拔节期每盆追施尿素0.87g。盆栽桶周围大田种植相同品种的小麦,田间管理措施按当地小麦高产栽培管理要求进行。
通过伸长节间诊断法(第三节间开始伸长)、叶龄余数诊断法(倒二叶出生)结合显微镜镜检观察小麦幼穗发育进程,试验于小麦幼穗分化的药隔形成期(2022年3月17日)选取植株长势基本一致的盆栽,于当日18:00放入智能超低温光照培养箱(DGXM−1008型,长1300mm ×宽 630mm ×高1305mm)内,进行温度处理,试验期间培养箱内温度变化如图1所示。处理结束后,将盆栽小麦转移至大田生长直至成熟。

图1 试验期间培养箱内温度变化
注:小麦幼穗分化的药隔形成期(2022年3月17日)进行低温胁迫处理(2℃和−2℃)4h,处理时间为1:00−5:00,箱内相对湿度设置为70%,光照强度0Lx,温度变幅±0.5℃,以10℃培养箱的小麦盆栽作为对照组CK,低温处理于次日7:00上升至CK处理温度。
Note:Low temperature stress treatments (2°C and −2°C) were applied for 4h (treatment time 1:00−5:00am, humidity set to 70% in the incubator, light intensity 0Lx, temperature variation ±0.5°C) during the anther differentiation period of young wheat spike differentiation (17 March, 2022), with wheat potted plants in a 10°C incubator as control CK, rising to CK treatment temperature at 7:00am the following day.
1.3 项目测定
1.3.1 花后不同时期小麦颖果形态观察
于小麦花后7d(2022年4月17日)、14d(2022年4月24日)、21d(2022年5月1日)、28d(2022年5月8日)和成熟期(2022年5月18日)取主茎穗第10小穗位不同粒位籽粒,在体视显微镜(SMZ171,中国)下观察拍摄颖果形态。根据前人研究结果[22−23],小穗第1、2位籽粒(G1、G2)为强势籽粒,第3、4位(G3、G4)籽粒为弱势籽粒,但不同品种间也存在一定差异,故以G2(强势粒)和G3(弱势粒)作为研究对象。
1.3.2 花后不同时期小麦颖果长度、宽度和厚度的测定
于小麦花后7d、14d、21d、28d和成熟期(38d)取主茎穗,各时期同一处理选取大小、长短基本一致的3株小麦穗,取麦穗第10小穗位颖果,用游标卡尺(MNT-150,中国)分别测量颖果的长度、宽度和厚度,重复测定3次,同时计算长/宽比、长/厚比和宽/厚比值。
1.3.3 花后不同时期小麦颖果显微结构观察
于小麦花后7d、14d、21d和28d取主茎穗第10小穗位不同粒位颖果,采用石蜡切片法进行颖果组织结构观察,将制成的切片在装有CCD(电荷耦合器件)相机的光学显微镜(DMLS,德国)下观察并拍照。通过CaseViewer 2.4(济南)软件对小麦颖果大、小淀粉粒的周长和面积进行测定。
1.4 数据统计与分析
采用Microsoft Excel 2019软件对实验数据进行整理和作图。采用SPSS 26.0进行方差分析,通过LSD法进行处理间的差异显著性比较(P<0.05)。
2 结果与分析
2.1 药隔期倒春寒对小麦颖果外观形态的影响
2.1.1 颖果外观形态观察
由图2可知,随着花后天数的增加,小麦颖果体积不断变大,28d时达到最大,之后随着颖果成熟,含水量降低,颖果体积变小。−2℃倒春寒胁迫下,XM26和YN19的G3粒位颖果均未结实。花后7d,XM26第10小穗位不同粒位颖果体积较小,YN19颖果体积较小且呈乳白色;花后14d,XM26在CK和2℃处理下的颖果为绿色,−2℃处理下的颖果为乳白色,而YN19颖果均为浅绿色。随着小麦颖果灌浆进程的推进,颖果体积增大,颜色逐渐变为黄绿色;成熟期(38d)小麦颖果皱缩,长度、宽度和厚度均变小。
2.1.2 颖果的长度、宽度和厚度
由图3和图4可知,随着花后天数的增加,XM26和YN19各处理G2、G3颖果的长、宽和厚度均呈递增趋势,于花后28d达到最大,在成熟期(38d)由于颖果含水量降低呈下降趋势。
与CK相比,成熟期XM26在2℃和−2℃倒春寒胁迫下G2粒位长度、宽度和厚度分别下降2.75%、4.41%,4.30%、10.22%和4.18%、9.40%;G3粒位2℃处理长度、宽度和厚度分别下降3.30%、2.05%和6.80%。与CK相比,成熟期YN19在2℃和−2℃倒春寒胁迫下G2粒位长度、宽度和厚度分别下降1.17%、2.77%,3.41%、5.68%和1.42%、1.70%;G3粒位2℃处理长度、宽度和厚度分别下降1.95%、1.57%和6.23%。因此,药隔期倒春寒导致两品种小麦颖果长度、宽度和厚度均减小,且降幅随着倒春寒胁迫程度的加强(2℃→−2℃)而增大,抗倒春寒性弱的品种XM26降幅更明显。此外,药隔期倒春寒对两品种小麦弱势粒位(G3)的长度、宽度和厚度的影响大小顺序均表现为厚度>长度>宽度;对两品种小麦强势粒位(G2)的影响则存在差异性,其中对XM26的G2颖果长度、宽度和厚度的影响大小表现为宽度>厚度>长度,对YN19的G2颖果影响则表现为宽度>长度>厚度。

图2 两品种小麦第10小穗位G2、G3粒位花后不同天数颖果形态发育观测结果
注:XM26(新麦26)为抗倒春寒性弱的小麦品种,烟农19(YN19)为抗倒春寒性强的小麦品种。第1、2位(G1、G2)籽粒为强势粒;第3、4位(G3、G4)籽粒为弱势粒,在−2℃处理下不结实。下同。
Note: XM26 (Xinmai 26) is a wheat variety with weak resistance to late spring coldness and YN19 (Yanong 19) is a wheat variety with strong resistance to late spring coldness. The 1stand 2nd(G1 and G2) grains are the superior grains; the 3rdand 4th(G3 and G4) grains are the inferior grains, which did not grow caryopsis at −2℃. The same as below.
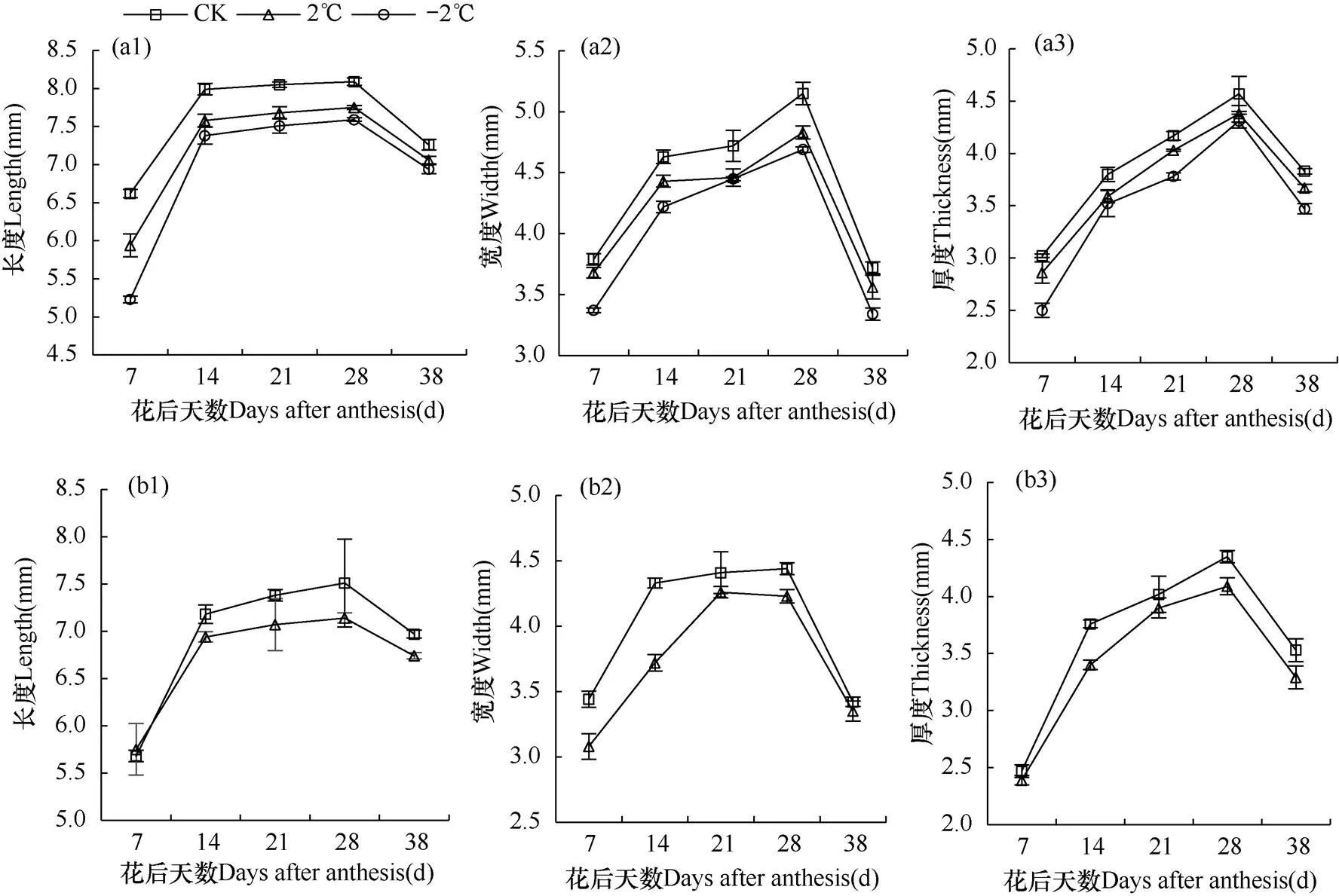
图3 品种XM26花后不同天数第10小穗位G2(a)、G3(b)粒位颖果长度(1)、宽度(2)和厚度(3)的观测结果
注:短线表示标准误差。下同。
Note: The bar is standard error. The same as below.
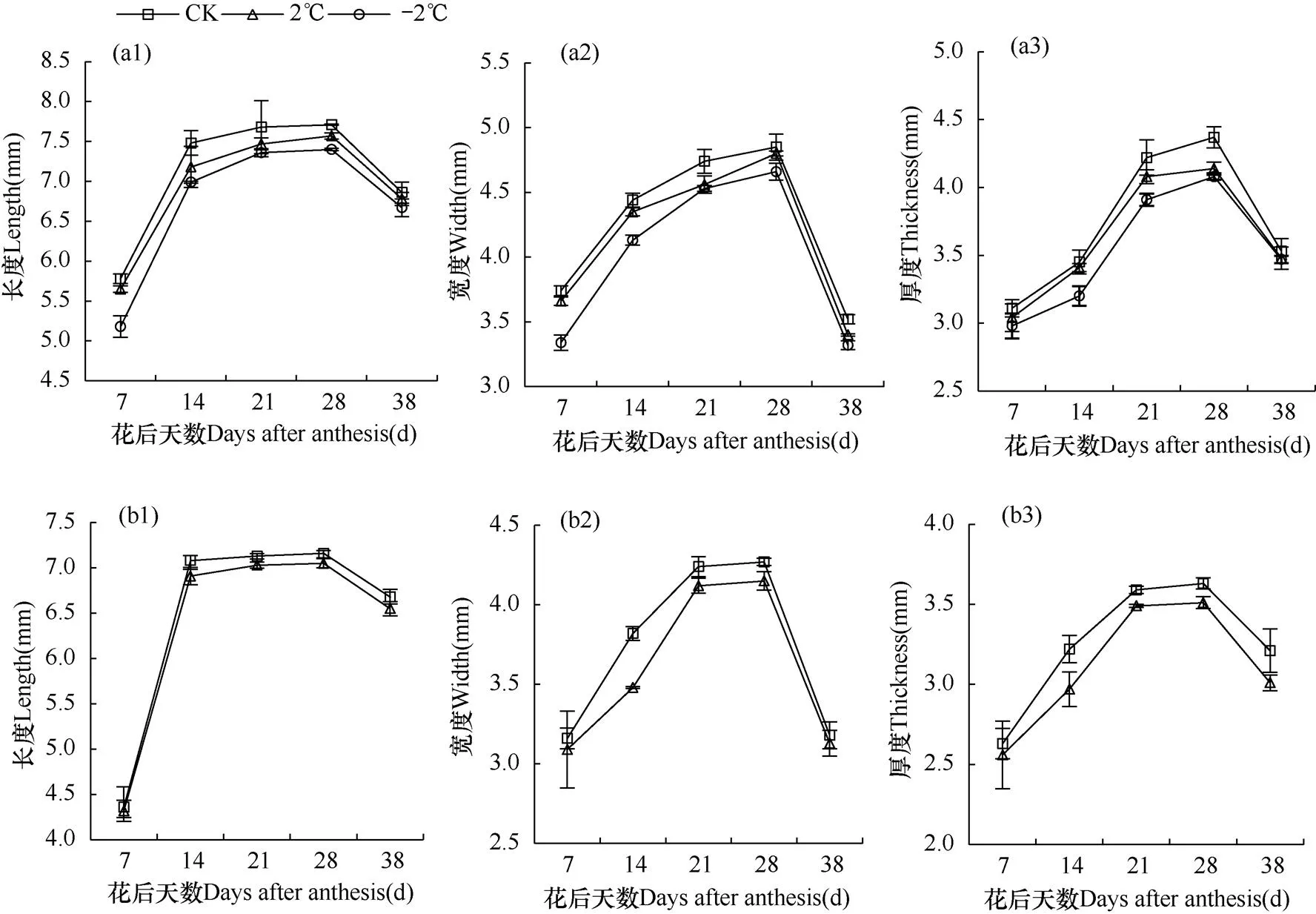
图4 品种YN19花后不同天数第10小穗位G2(a)、G3(b)粒位颖果长度(1)、宽度(2)和厚度(3)的观测结果
2.2 药隔期倒春寒对小麦颖果长宽比、长厚比及宽厚比的影响
2.2.1 长宽比
由图5可知,两品种小麦在成熟期由于颖果含水量降低、体积缩小,长度/宽度比值达到最大值。成熟期XM26和YN19的G2粒位颖果在CK、2℃、−2℃处理下比值分别为1.95、1.98、2.08和1.95、1.99、2.01;G3粒位在CK、2℃处理下分别为2.04、2.01和2.10、2.09。总体来看,随着倒春寒胁迫程度的加强(2℃→−2℃),成熟期XM26和YN19的G2粒位颖果长/宽度比值均呈增加趋势,而G3粒位则均呈降低趋势。
2.2.2 长厚比
由图6可知,成熟期XM26和YN19的G2粒位颖果在CK、2℃、−2℃处理长度/厚度比值分别为1.90、1.92、2.00和1.95、1.95、1.92;G3粒位在CK、2℃处理下分别为2.04、2.01和2.08、2.17。总体来看,随着倒春寒胁迫程度的加强(2℃→−2℃),成熟期XM26的G2粒位颖果长度/厚度比值呈增加趋势,G3粒位呈降低趋势;而成熟期YN19的G2粒位颖果长度/厚度比呈降低趋势,G3粒位呈增加趋势。
2.2.3 宽厚比
由图7可知,XM26的G2、G3粒位颖果在CK、2℃、−2℃处理下宽度/厚度比值均呈下降趋势,成熟期达到最小值。成熟期XM26和YN19的G2在CK、2℃、−2℃处理颖果宽度/厚度比值分别为0.97、0.97、0.96和1.00、0.98、0.96;G3粒位在CK、2℃处理下分别为0.97、1.02和0.99、1.04。总体来看,随着倒春寒胁迫程度的加强(2℃→−2℃),成熟期XM26和YN19的G2粒位颖果宽/厚比值均呈降低趋势,G3粒位均呈增加趋势。
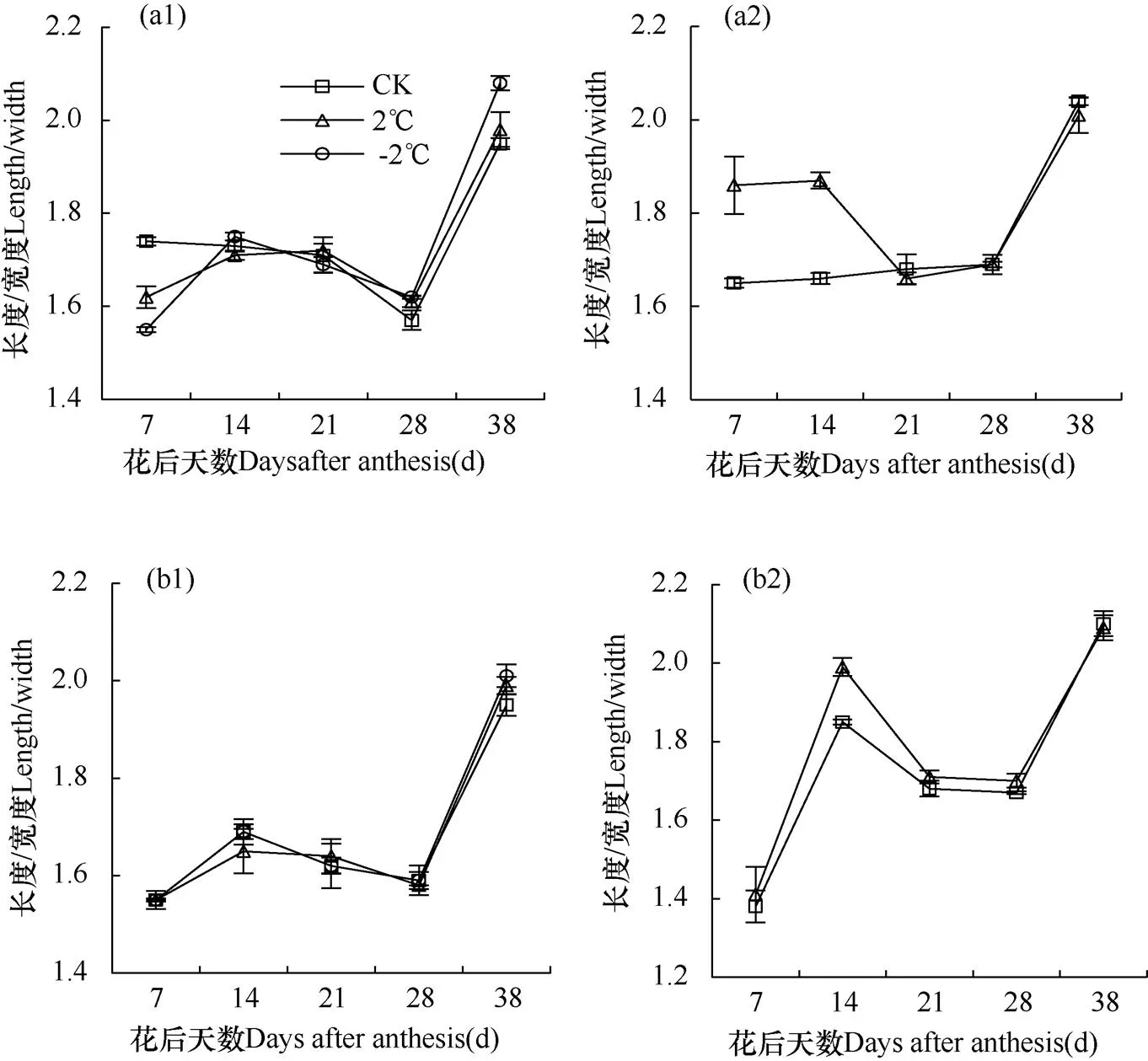
图5 两品种小麦XM26(a)和YN19(b)花后不同天数第10小穗位G2(1)、G3(2)粒位颖果长/宽比值变化

图6 两品种小麦XM26(a)和YN19(b)花后不同天数第10小穗位G2(1)、G3(2)粒位颖果长/厚比值变化

图7 两品种小麦XM26(a)和YN19(b)花后不同天数第10小穗位G2(1)、G3(2)粒位颖果宽/厚比值变化
综上所述,成熟期两品种小麦各处理弱势粒(G3)颖果长/宽、长/厚、宽/厚比值均高于强势粒(G2)。药隔期倒春寒胁迫对成熟期小麦强、弱势颖果的长/宽、长/厚、宽/厚比值影响均呈相反趋势。
2.3 药隔期倒春寒对小麦颖果显微结构的影响
2.3.1 小麦颖果果皮显微结构观察
由图8可见,药隔期倒春寒对两品种小麦果皮显微结构影响明显,主要表现在中果皮的凋亡及其细胞内淀粉粒的降解。XM26的G2粒位颖果在花后7d各处理颖果的中果皮细胞开始凋亡,出现凋亡空腔,CK较2℃和−2℃处理颖果的中果皮细胞内淀粉粒降解较多;花后21d各处理中果皮细胞内淀粉粒未完全降解;花后28d各处理中果皮凋亡(图8a)。XM26的G3粒位颖果在花后7d时CK处理中果皮凋亡较多,2℃处理中果皮刚开始凋亡;花后21d在2℃处理下中果皮凋亡;花后28d时CK处理中果皮凋亡(图8c)。YN19的G2粒位颖果在花后 7d各处理中果皮开始凋亡,出现凋亡空腔;21d时各处理中果皮凋亡(图8b)。YN19的G3粒位颖果在花后7d时CK处理中果皮开始凋亡,2℃处理中果皮未出现凋亡;花后21d在CK和2℃处理下果皮凋亡(图 8d)。由此可知,药隔期倒春寒会减缓早期颖果果皮细胞的降解及中果皮细胞中淀粉粒的降解速率。
2.3.2 小麦颖果胚乳大、小淀粉粒观察
从图9可见,XM26的G2粒位颖果在CK处理花后7d颖果胚乳细胞内出现淀粉粒和细胞核,花后14d淀粉粒聚集在细胞壁周围,花后21~28d出现大、小淀粉粒,并充满整个胚乳细胞;2℃和−2℃处理7d胚乳细胞暂未出现淀粉粒,只有细胞核,14d开始出现淀粉粒(图9a)。XM26的G3粒位颖果在CK和2℃处理7d胚乳细胞只有细胞核,暂未出现淀粉粒,14d开始出现淀粉粒(图9c)。YN19的G2粒位颖果在CK、2℃和−2℃处理7d均未出现淀粉粒,只有细胞核,14d才出现淀粉粒,21~28d大、小淀粉体逐渐充满整个胚乳细胞(图9b);YN19的G3粒位颖果在CK和2℃处理7d均未出现淀粉粒,只有细胞核,14d才出现淀粉粒(图9d)。综上所述,花后14d之前胚乳先积累大淀粉粒,花后14d之后开始积累小淀粉粒并逐渐充满整个胚乳细胞。药隔期倒春寒抑制了两品种小麦颖果胚乳细胞淀粉粒的发育,抗倒春寒性弱的品种XM26较抗倒春寒性强的品种YN19先积累淀粉粒,且弱势粒(G3)较强势粒(G2)受倒春寒影响更大。
由图10可知,随着花后天数的增加,颖果胚乳中大淀粉粒周长和面积也随之增加。两品种小麦强势粒(G2)胚乳细胞中大淀粉粒的周长和面积均大于弱势粒(G3)。抗倒春寒性弱的品种XM26在花后28d强势粒(G2)胚乳细胞中大淀粉粒在2℃、−2℃处理下周长和面积分别下降8.17%、14.27%和14.11%、16.49%;弱势粒(G3)在2℃处理下周长和面积下降10.31%和6.56%。抗倒春寒性强的品种YN19在花后28d强势粒(G2)颖果胚乳细胞中大淀粉粒在2℃、−2℃处理下周长和面积下降8.67%、14.66%和14.08%、17.98%;弱势粒(G3)在2℃处理周长和面积下降3.08%和8.70%。综上所述,随着倒春寒胁迫程度的加强(2℃→−2℃),强势粒颖果胚乳中大淀粉粒周长和面积较弱势粒下降较多。
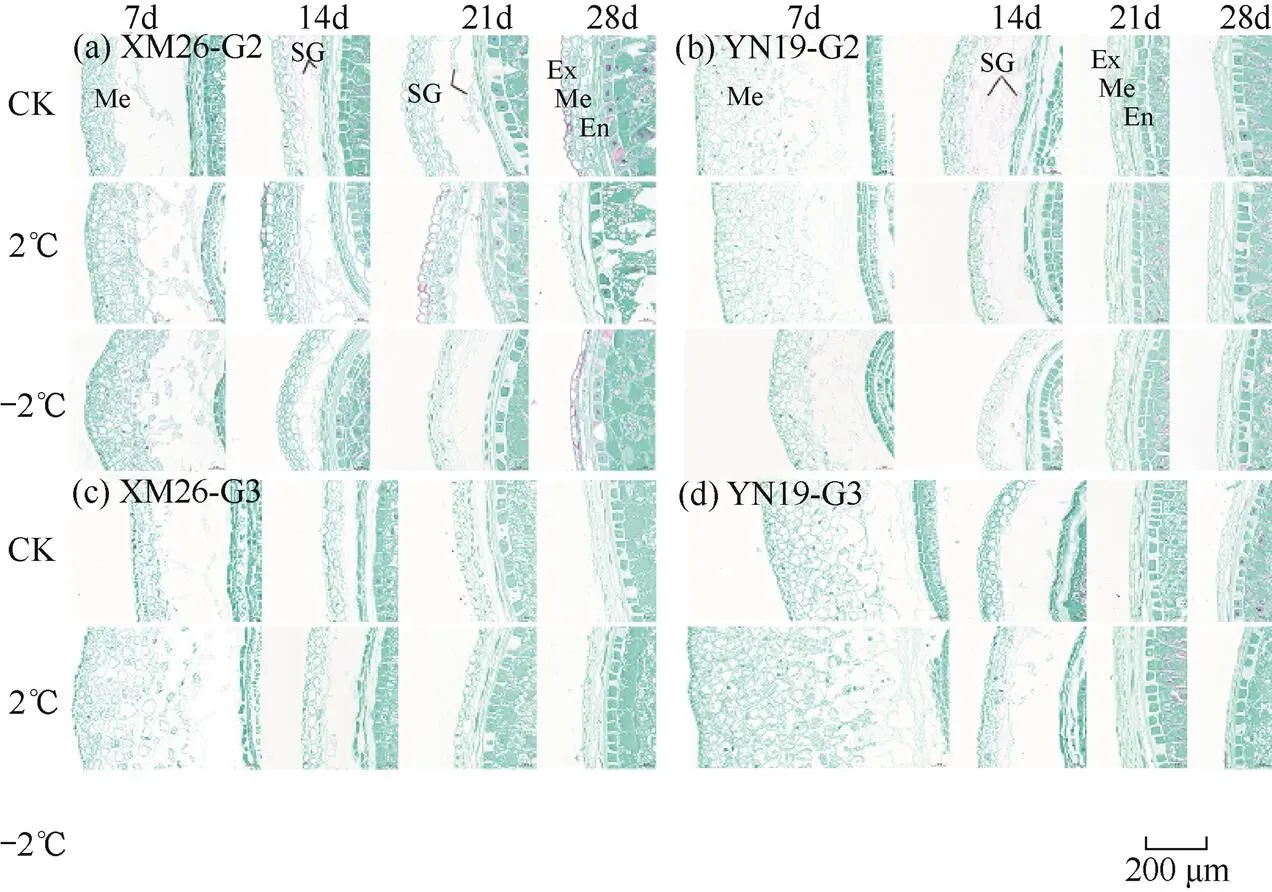
图8 两品种小麦花后不同天数第10小穗位G2、G3粒位颖果果皮细胞显微结构
注:Me为中果皮;Ex为外果皮;En为内果皮;SG为淀粉粒。G3粒位颖果在−2℃处理下不结实,故不显示。下同。
Note: Me is mesocarp; Ex is exocarp; En is endocarp; SG is starch granule. G3 grain caryopsis did not set at −2℃, so it did not show. The same as below.
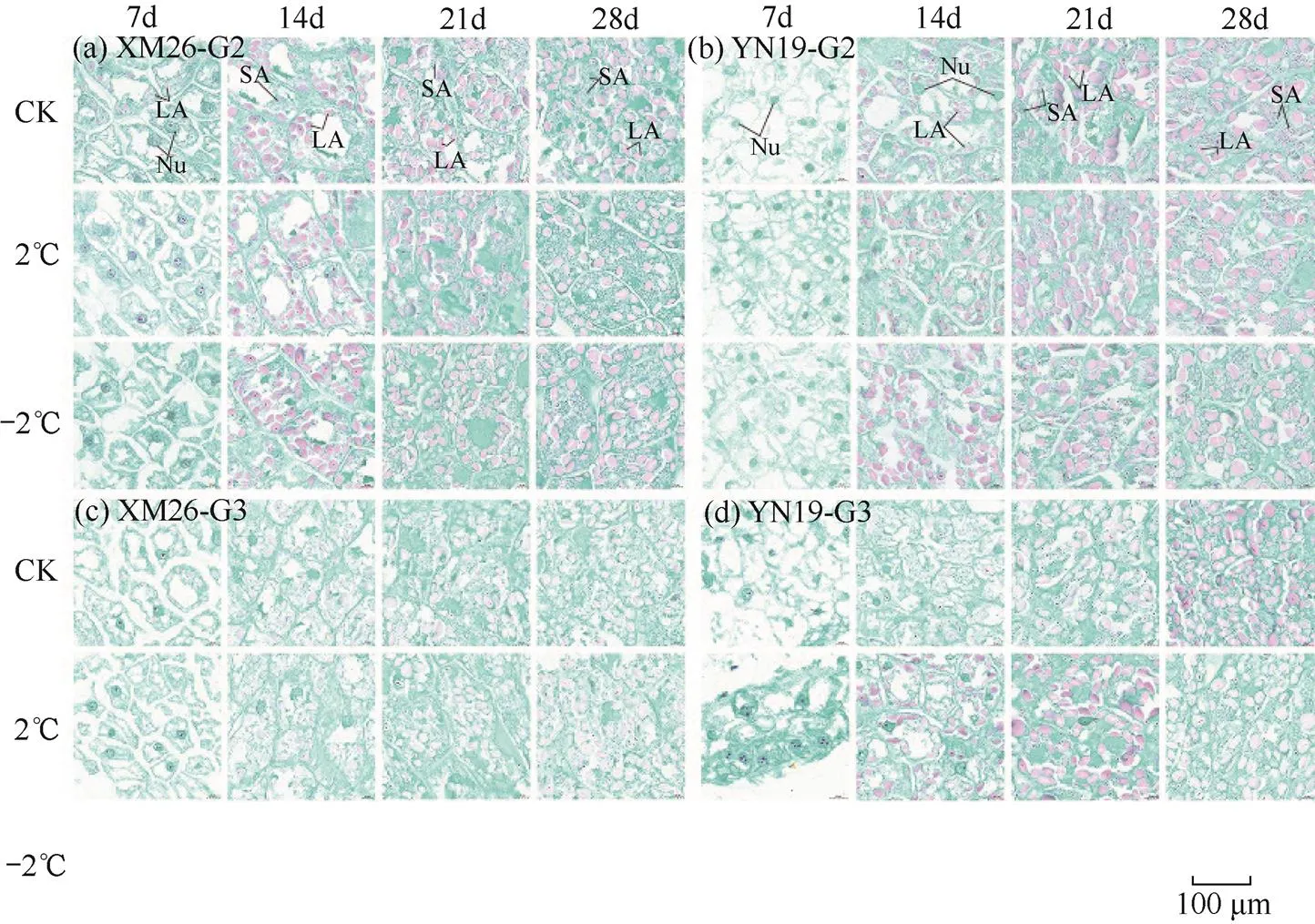
图9 两品种小麦花后不同天数第10小穗位G2、G3粒位颖果胚乳细胞显微结构
注:Nu为细胞核;LA为大淀粉粒;SA为小淀粉粒。
Note: Nu is nucleus; LA is large amyloplast; SA is small amyloplast.

图10 两品种小麦花后不同天数第10小穗位G2、G3粒位籽粒大淀粉粒周长和面积
注:小写字母表示处理间在0.05水平上的差异显著性。下同。
Note:Lowercase indicates the difference significance among treatments at 0.05 level. The same as below.
由图11可知,花后7~14d籽粒胚乳细胞中暂未出现小淀粉粒,花后21~28d小淀粉粒的周长和面积逐渐增加。两品种小麦强势粒(G2)胚乳细胞中小淀粉粒的周长和面积均大于弱势粒(G3)。抗倒春寒性弱的品种XM26在花后28d强势粒(G2)颖果胚乳细胞中小淀粉粒在2℃、−2℃处理下周长和面积分别下降1.55%、2.58%和3.96%、9.57%;弱势粒(G3)在2℃处理下周长和面积下降3.17%和11.85%。抗倒春寒性强的品种YN19在花后28d强势粒(G2)颖果胚乳细胞中小淀粉粒在2℃、−2℃处理下周长和面积下降0.94%、5.00%和2.59%、10.03%;弱势粒(G3)在2℃处理下周长和面积下降6.39%和16.17%。综上所述,随着倒春寒胁迫程度的加强(2℃→−2℃),弱势粒颖果胚乳中小淀粉粒周长和面积较强势粒下降较多。
3 结论与讨论
3.1 讨论
3.1.1 药隔期倒春寒对小麦颖果形态的影响
朱启迪等[24]研究表明从受精到籽粒成熟,小麦籽粒历时30~40d,随着花后天数的增加,其籽粒长和宽整体呈增加趋势,后期随着籽粒成熟和含水量的下降,籽粒长宽略有降低。本研究结果与其一致,但倒春寒胁迫会延缓小麦颖果发育进程,降低颖果体积。低温胁迫影响小麦籽粒的粒径大小,Liu等[25]研究表明拔节期低温会降低小麦颖果的长度和宽度。本研究也表明药隔期倒春寒会降低两品种小麦颖果的长度、宽度和厚度,降幅随着倒春寒胁迫程度的加重而增大,且抗倒春寒性弱的品种XM26较抗倒春寒性强的品种YN19降幅更大。此外,倒春寒胁迫对弱势粒颖果的厚度影响最大,对强势粒颖果的影响则存在差异。这可能是因为对粒形相关性状的QTL进行定位分析发现相较于籽粒长度和宽度,颖果厚度最易受环境变化的影响[26−27]。Liu等[25]研究表明拔节期低温导致小麦颖果的长/宽值增加,本研究也表明成熟期两品种小麦的G2粒位的颖果长/宽均随着倒春寒胁迫程度的加强呈增加趋势。成熟期两品种小麦颖果长/宽、长/厚比值约为2.0,而宽/厚比值约为1.0,颖果形状呈现“长大宽小厚小”的形态,表明倒春寒胁迫会降低颖果的库容和充实度,这可能是粒重降低的主要原因。
3.1.2 药隔期倒春寒对小麦胚乳淀粉粒分布的影响
小麦颖果的生长发育伴随着胚乳发育和淀粉积累,淀粉的合成与积累过程直接影响小麦的产量和品质[28−29]。淀粉是小麦颖果胚乳中碳储备的主要形式,占籽粒最终干重的65%~75%[30]。积累在胚乳细胞淀粉体内的淀粉可分为大、小两种类型,即A型和B型淀粉粒[31]。前人研究表明,小麦花后14~16d之前颖果胚乳中先积累较大的A型淀粉粒,随后较小的B型淀粉粒快速增加[32−34]。这与本研究结果基本一致,且倒春寒胁迫并没有改变小麦颖果胚乳细胞中大、小淀粉体发育的时间顺序,但抗倒春寒性弱的品种XM26较抗倒春寒性强的品种YN19先积累淀粉粒。谭秀山等[22,35]研究表明,强势籽粒中大、小淀粉体积累量均高于弱势籽粒,这与幼穗发育过程中强势小花形成早、开花早及花后营养物质积累早且淀粉积累速率高有着密切关系。谭植等[36]研究表明,拔节期低温对小麦颖果大、小淀粉粒体积和表面积百分比有显著影响,小淀粉粒体积和表面积百分比受低温胁迫后显著降低。本研究则表明,药隔期倒春寒会降低小麦强、弱势颖果中的大、小淀粉粒周长和面积,强势粒位籽粒中大淀粉粒周长和面积下降较多,而弱势粒位籽粒中小淀粉粒周长和面积下降较多。这可能是因为不同大小的淀粉粒在胚乳发育过程中表现出不同的理化和功能特性[37],药隔期倒春寒对不同粒位籽粒中的大、小淀粉粒影响也不同。
Calderini等[38]研究指出花前是决定小麦粒重潜力的关键时期。杨佳庆等[39]研究表明,花后低温降低小麦胚乳发育早期和中期淀粉体和蛋白体的积累量,缩短灌浆时期,降低小麦粒重。汪巧菊等[40]研究表明强势粒较弱势粒的胚乳质地更加致密且背部胚乳细胞中淀粉体和蛋白体的相对面积大、充实度高。陈红阳等[41]研究表明结实期低温胁迫对弱势粒中淀粉合成关键酶活性变化影响大于强势粒,且弱势粒淀粉合成积累速率慢、含量降低,最终导致千粒重显著降低。本研究表明,药隔期倒春寒抑制了小麦颖果胚乳细胞淀粉粒的发育,从而导致小麦粒重降低,且弱势粒较强势粒受倒春寒影响更大。这可能是因为小麦花后各时期强势粒的粒重、蛋白质积累量和蛋白质含量显著大于弱势粒[20],且强势粒在灌浆期间获得的光合产物多、物质积累快,而弱势粒同化物供应不足、物质积累慢[42]。
本研究分析了药隔期倒春寒对小麦中部小穗(第10小穗位)不同粒位颖果的粒径、胚乳细胞和大、小淀粉粒的影响,明确了倒春寒胁迫对弱势粒颖果产生的影响更大。由于试验低温胁迫处理是培养箱内进行的,与大田自然倒春寒有所不同,会存在一定差异性。故后续还需增加倒春寒胁迫降温的梯度和幅度以及胁迫持续时间,同时选取上部和下部小穗不同粒位颖果进行测定,以进一步量化不同等级倒春寒胁迫对小麦颖果发育的伤害机理,并与大田自然倒春寒灾害和防灾减灾措施相结合,以期最大限度地降低倒春寒灾害对小麦产量的影响。
3.2 结论
(1)药隔期倒春寒会降低小麦颖果的长度、宽度和厚度,且降幅随着倒春寒胁迫程度的加强(2℃→ −2℃)而增大,对小麦弱势粒颖果的长度、宽度和厚度的影响均表现为厚度>长度>宽度,对强势粒颖果的影响则存在差异。倒春寒胁迫下成熟期小麦各处理弱势粒颖果长/宽、长/厚、宽/厚比值均高于强势粒颖果。
(2)药隔期倒春寒会减缓早期颖果果皮降解速率,同时还会抑制两品种小麦颖果胚乳细胞淀粉粒的发育。药隔期倒春寒降低小麦强、弱势颖果胚乳中大、小淀粉粒的周长和面积,随着倒春寒胁迫程度的加强,强势粒中大淀粉粒周长和面积较弱势粒下降较多,而弱势粒中小淀粉粒周长和面积较强势粒下降较多。
(3)药隔期倒春寒对小麦弱势粒的影响大于强势粒,且对小麦颖果的影响随胁迫强度的加重而增加。生产中可通过选用抗倒春寒小麦品种加上氮磷钾肥的合理运筹来提高植株对倒春寒的抵抗能力,减轻倒春寒胁迫对小麦弱势粒的不利影响,从而提高小麦生产的防灾减灾能力。
[1] Šramková Z,Gregová E,Šturdík E.Chemical composition and nutritional quality of wheat grain[J].Acta Chimica Slovaca,2009,2(1):115-138.
[2] Mallick S A,Azaz K,Gupta M,et al.Characterization of grain nutritional quality in wheat[J].Indian Journal of Plant Physiology,2013,18(2):183-186.
[3] 陈翔,于敏,蔡洪梅,等.小麦倒春寒研究现状与进展[J].应用生态学报,2021,32(8):2999-3009.
Chen X,Yu M,Cai H M,et al.Current status and research advances of late spring coldness in wheat[J].Chinese Journal of Applied Ecology,2021,32(8):2999-3009.(in Chinese)
[4] Li X N,Cai J,Liu F L,et al.Wheat plants exposed to winter warming are more susceptible to low temperature stress in the spring[J].Plant Growth Regulation,2015,77(1):11-19.
[5] Liu L L,Ji H T,An J P,et al.Response of biomass accumulation in wheat to low-temperature stress at jointing and booting stages[J].Environmental and Experimental Botany,2019,157:46-57.
[6] 王妍,张晓龙,石嘉丽,等.中国冬小麦主产区气候变化及其对小麦产量影响研究[J].中国生态农业学报(中英文), 2022,30(5):723-734.
Wang Y,Zhang X L,Shi J L,et al.Climate change and its effect on winter wheat yield in the main winter wheat production areas of China[J].Chinese Journal of Eco-Agriculture,2022,30(5):723-734.(in Chinese)
[7] 李金才,陈翔,刘惠惠,等.安徽省地方标准《DB34 /T 3736- 2020》:小麦倒春寒综合防控技术规程[S].合肥:安徽省市场监督管理局,2020.
Li J C,Chen X,Liu H H,et al.Local Standards of Anhui Province (DB34/T 3736-2020):Technical regulation for comprehensive prevention and control of late spring coldness in wheat[S].Hefei:Market Supervision and Administration Bureau of Anhui Province,2020.(in Chinese)
[8] 张乐乐,陈翔,柯媛媛,等.冬小麦抗倒春寒性能鉴定方法和指标的比较[J].中国农业气象,2021,42(2):146-157.
Zhang L L,Chen X,Ke Y Y,et al.An inclusive comparison of identification methods and indices of winter wheat tolerance against late spring coldness[J].Chinese Journal of Agrometeorology,2021,42(2):146-157.(in Chinese)
[9] Li X N,Pu H C,Liu F L,et al.Winter wheat photosynthesis and grain yield responses to spring freeze[J].Agronomy Journal,2015,107(3):1002-1010.
[10] 柯媛媛,陈翔,张乐乐,等.药隔期低温胁迫对小麦干物质积累、转运和分配及产量的影响[J].安徽农业大学学报,2021,48(5):701-706.
Ke Y Y,Chen X,Zhang L L,et al.Effects of low temperature stress at anther connective stage on dry matter accumulation, translocation and distribution and grain yield of wheat[J]. Journal of Anhui Agricultural University,2021, 48(5):701- 706.(in Chinese)
[11] Xiao L,Asseng S,Wang X,et al.Simulating the effects of low-temperature stress on wheat biomass growth and yield[J].Agricultural and Forest Meteorology,2022,326: 109191.
[12] 刘蕾蕾,纪洪亭,刘兵,等.拔节期和孕穗期低温处理对小麦叶片光合及叶绿素荧光特性的影响[J].中国农业科学, 2018,51(23):4434-4448.
Liu L L,Ji H T,Liu B,et al.Effects of jointing and booting low temperature treatments on photosynthetic and chlorophyll fluorescence characteristics in wheat leaf[J]. Scientia Agricultura Sinica,2018,51(23):4434-4448. (in Chinese)
[13] 马泉,张玉雪,陶源,等.春季不同时期低温对小麦光合特性和粒重形成的影响[J].麦类作物学报,2022,42(2):226- 235.
Ma Q,Zhang Y X,Tao Y,et al.Effects of low temperature at different stages in spring on photosynthetic characteristics and grain weight formation of wheat[J].Journal of Triticeae Crops,2022,42(2):226-235.(in Chinese)
[14] 李春燕,徐雯,刘立伟,等.低温条件下拔节期小麦叶片内源激素含量和抗氧化酶活性的变化[J].应用生态学报, 2015,26(7):2015-2022.
Li C Y,Xu W,Liu L W,et al.Changes of endogenous hormone contents and antioxidative enzyme activities in wheat leaves under low temperature stress at jointing stage[J].Chinese Journal of Applied Ecology,2015,26(7): 2015-2022.(in Chinese)
[15] 刘丽杰,苍晶,王军虹,等.外源ABA对苗期低温下冬小麦蔗糖含量及其关键酶基因表达的影响[J].麦类作物学报, 2015,35(10):1333-1340.
Liu L J,Cang J,Wang J H,et al.Effect of exogenous abscisic acid on sucrose content and expression of key enzyme genes controlling sucrose metabolism in seedling period of winter wheat under cold stress[J].Journal of Triticeae Crops,2015,35(10):1333-1340.(in Chinese)
[16] Zeng Y,Yu J,Cang J,et al.Detection of sugar accumulation and expression levels of correlative key enzymes in winter wheat() at low temperatures[J]. Bioscience, Biotechnology,and Biochemistry,2011,75(4):681-687.
[17] Zhang W J,Wang J Q,Huang Z L,et al.Effects of low temperature at booting stage on sucrose metabolism and endogenous hormone contents in winter wheat spikelet[J]. Frontiers in Plant Science,2019,10:498.
[18] Zhang W J,Zhao Y,Li L Y,et al.The effects of short-term exposure to low temperatures during the booting stage on starch synthesis and yields in wheat grain[J].Frontiers in Plant Science,2021,12:684784.
[19] Jiang G,Hassan M A,Muhammad N,et al.Comparative physiology and transcriptome analysis of young spikes in response to late spring coldness in wheat (L.)[J].Frontiers in Plant Science,2022,13:811884.
[20] 李豪圣,曹新有,宋健民,等.不同小麦品种粒重和蛋白质含量的穗粒位效应分析[J].作物学报,2017,43(2):238-252.
Li H S,Cao X Y,Song J M,et al.Effects of spikelet and grain positions on grain weight and protein content of different wheat varieties[J].Acta Agronomica Sinica,2017,43(2): 238-252.(in Chinese)
[21] Zheng J C,Liu T,Zheng Q X,et al.Identification of cold tolerance and analysis of genetic diversity for major wheat varieties in Jianghuai region of China[J].Pakistan Journal of Botany,2020,52(3):839-849.
[22] 谭秀山,毕建杰,王金花,等.冬小麦不同穗位籽粒淀粉粒差异及其与粒重的相关性[J].作物学报,2012,38(10): 1920-1929.
Tan X S,Bi J J,Wang J H,et al.Differences of starch granules in grains from different spikelet positions and their correlation with grain weight in winter wheat[J].Acta Agronomica Sinica,2012,38(10):1920-1929.(in Chinese)
[23] 吴云飞,段玉仁,张勇,等.小麦强、弱势籽粒品质研究进展[J].麦类作物学报,2022,42(7):808-814.
Wu Y F,Duan Y R,Zhang Y,et al.Progress on wheat grain quality of superior and inferior grains[J].Journal of Triticeae Crops,2022,42(7):808-814.(in Chinese)
[24] 朱启迪,李艳艳,卢萌,等.小麦不同穗位籽粒品质和形态性状分析[J].作物杂志,2022,211(6):88-92.
Zhu Q D,Li Y Y,Lu M,et al.Analysis of wheat kernel quality and morphological characteristics at different spikelet positions[J].Crops,2022,211(6):88-92.(in Chinese)
[25] Liu L,Song H,Shi K,et al.Response of wheat grain quality to low temperature during jointing and booting stages:on the importance of considering canopy temperature[J]. Agricultural and Forest Meteorology,2019,278:107658.
[26] 王松凤,李辉,刘喜,等.水稻粒形相关性状及千粒重QTL的稳定性分析[J].南京农业大学学报,2008,31(3):1-7.
Wang S F,Li H,Liu X,et al.Stability analysis of QTL for the traits related to grain shape and 1000-grain weight in rice[J].Journal of Nanjing Agricultural University,2008,31 (3):1-7.(in Chinese)
[27] 陈佳慧,兰进好,王晖,等.小麦籽粒形态及千粒重性状的QTL初步定位[J].麦类作物学报,2011,31(6):1001-1006.
Chen J H,Lan J H,Wang H,et al.QTL mapping for traits of kernel morphology and grain weight in common wheat[J]. Journal of Triticeae Crops,2011,31(6):1001-1006.(in Chinese)
[28] 刘娟,夏文荣,李运,等.紫糯小麦籽粒石蜡切片制作技术优化及淀粉粒累积分析[J].麦类作物学报,2022,42(1): 42-53.
Liu J,Xia W R,Li Y,et al.Optimization of paraffin sectioning technology and accumulation analysis of starch granules for grains of purple waxy wheat[J].Journal of Triticeae Crops,2022,42(1):42-53.(in Chinese)
[29] Wang Z,Li W,Qi J,et al.Starch accumulation, activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch contents[J].Journal of Food Science and Technology,2014,51(3):419-429.
[30] Yang J,Zhang J,Wang Z,et al.Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling[J].Plant Physiology,2004, 135(3):1621-1629.
[31] 余徐润,周亮,荆彦平,等.Image-Pro Plus软件在小麦淀粉粒显微图像分析中的应用[J].电子显微学报,2013,32(4): 344-351.
Yu X R,Zhou L,Jing Y P,et al.Application of Image-pro plus in analysis of wheat starch granule microscopic image[J].Journal of Chinese Electron Microscopy Society, 2013,32(4):344-351.(in Chinese)
[32] 鲁平,陈俊,吴媛媛,等.糯小麦胚乳发育过程中淀粉粒形态变化及A、B型淀粉粒的显微观察[J].中国农业大学学报,2015,20(4):29-35.
Lu P,Chen J,Wu Y Y,et al.Microscopic study of morphological changes of A and B type starch grains during the development of endosperm in waxy wheat[J]. Journal of China Agricultural University,2015,20(4):29-35. (in Chinese)
[33] 李文阳,闫素辉,王振林.小麦胚乳A、B型淀粉粒的形成与生长特征及氮素调节[J].中国粮油学报,2016,31(5): 22-26.
Li W Y,Yan S H,Wang Z L.Formation and developmental characteristics of A and B type starch granule in wheat endosperm and response to nitrogen[J].Journal of the Chinese Cereals and Oils Association,2016,31(5):22-26.(in Chinese)
[34] 陶莉敏,周琴,蔡剑,等.施硫时期对小麦颖果和胚乳发育的影响[J].南京农业大学学报,2018,41(3):402-412.
Tao L M,Zhou Q,Cai J,et al.Effects of sulfur fertilizer application periods on caryopsis and endosperm development in wheat[J].Journal of Nanjing Agricultural University,2018, 41(3):402-412.(in Chinese)
[35] 荆彦平,刘大同,李栋梁,等.小麦内胚乳细胞及其淀粉体的发育[J].麦类作物学报,2013,33(4):818-824.
Jing Y P,Liu D T,Li D L,et al.Development of starch endosperm cells and amyloplasts in wheat[J].Journal of Triticeae Crops,2013,33(4):818-824.(in Chinese)
[36] 谭植,闫素辉,刘良柏,等.拔节期低温对小麦穗花发育与籽粒淀粉粒分布的影响[J].西北农业学报,2021,30(5): 637-644.
Tan Z,Yan S H,Liu L B,et al.Effect of low temperature at jointing stage on spikelet and floret development and starch granule size distribution of wheat[J].Acta Agriculturae Boreali-occidentalis Sinica,2021,30(5):637-644.(in Chinese)
[37] 王磊磊.麦稻不同粒位颖果发育差异的机制及其对氮素响应研究[D].扬州:扬州大学,2022.
Wang L L.Mechanism of caryopsis developmental differences at different grain positions in wheat and rice and its response to nitrogen[D].Yangzhou:Yangzhou University, 2022.(in Chinese)
[38] Calderini D F,Reynolds M P.Changes in grain weight as a consequence of de-graining treatments at pre- and post- anthesis in synthetic hexaploid lines of wheat (×.)[J].Functional Plant Biology,2000,27 (3):183-191.
[39] 杨佳庆,郝朵,冉莉萍,等.花后低温条件下小麦颖果发育的显微结构观察[J].西北植物学报,2022,42(4):578-588.
Yang J Q,Hao D,Ran L P,et al.Microstructural observation of caryopsis development in wheat under low temperature after anthesis[J].Acta Botanica Boreali-Occidentalia Sinica, 2022,42(4):578-588.(in Chinese)
[40] 汪巧菊,陈昕钰,冉莉萍,等.小麦强弱势粒胚乳淀粉体和蛋白体发育及物质积累研究[J].麦类作物学报,2019, 39 (1):89-95.
Wang Q J,Chen X Y,Ran L P,et al.Study on amyloid and protein development and their accumulation in endosperm between superior and inferior wheat grains[J].Journal of Triticeae Crops,2019,39(1):89-95.(in Chinese)
[41] 陈红阳,贾琰,赵宏伟,等.结实期低温胁迫对水稻强、弱势粒淀粉形成与积累的影响[J].中国水稻科学,2022, 36(5):487-504.
Chen H Y,Jia Y,Zhao H W,et al.Effects of low temperature stress during grain filling on starch formation and accumulation of superior and inferior grains in rice[J]. Chinese Journal of Rice Science,2022,36(5):487-504. (in Chinese)
[42] 程敦公,王灿国,郭军,等.不同光合器官对小麦强势和弱势籽粒产量及蛋白质含量的影响[J].华北农学报,2019, 34(S1):43-48.
Cheng D G,Wang C G,Guo J,et al.Effects of different photosynthetic organs on the yield and protein content of superior and inferior wheat kernels[J].Acta Agriculturae Boreali-Sinica,2019,34(S1):43-48.(in Chinese)
Effect of Late Spring Coldness during the Anther Differentiation Period on the Caryopsis Development of Wheat
WENG Ying1, WANG Peng-na1, YU Min1, DAI Wen-ci1, WU Yu1, CAI Hong-mei1, ZHENG Bao-qiang1, LI Jin-cai1,2, CHEN Xiang1
(1. College of Agronomy, Anhui Agricultural University/Crop Cultivation Science Observatory in East China of the Ministry of Agriculture and Rural Affairs, Hefei 230036, China; 2. Collaborative Innovation Center of Modern Crop Production in Jiangsu, Nanjing 210095)
A field pot experiment was conducted using Xinmai26 (XM26) with weak resistance to late spring coldness and Yannong19 (YN19) with strong resistance to late spring coldness. Low temperature (2℃ and −2℃) stress treatment was carried out in intelligent ultra-low temperature light incubator during the anther differentiation period of young wheat spike differentiation, and 10℃ was used as CK control group. The length, width and thickness of the superior and inferior caryopsis of wheat were analyzed by sampling every 7 days after anthesis. At the same time, the microstructures of pericarp, endosperm cells and large and small starch grains were observed by paraffin sectioning technique, so as to explore the microstructures of caryopsis development in wheat under the late spring coldness during the anther differentiation period. The results showed as follows: (1) the length, width and thickness of wheat caryopsis decreased with the increase of cold stress (2℃→−2℃). The length, width and thickness of wheat caryopsis decreased by 1.17%−4.41%, 1.57%−10.22% and 1.42%−9.40%, respectively, at the mature stage. The ratios of length/width, length/thickness and width/thickness of inferior grain caryopsis were higher than that of superior grain caryopsis in all treatments at maturity stage. (2) The degradation rate of early caryopsis pericarp was slowed down. Meanwhile, the development of starch granules in caryopsis endosperm cells of two wheat cultivars was inhibited. (3) The circumference and area of large and small starch grains in wheat superior and inferior caryopsis were decreased by 8.17%−14.66%, 14.08%−17.98%, 0.94%−5.00% and 2.59%−10.03% in endosperm cells of dominant caryopsis at 28 days after anthems. The circumference and area of large and small starch grains decreased by 3.08%−10.31%, 6.59%−8.70%, 3.17%−6.39% and 11.85%−16.17% in endosperm cells of inferior grain caryopsis. The circumference and area of large starch grains decreased more in dominant grain caryopsis, while the circumference and area of small and medium starch grains decreased more in inferior grain caryopsis. In conclusion, late spring coldness during the anther differentiation period can slow down the degradation of pericarp cells of early wheat caryopsis development, inhibit the development of starch grains in endosperm cells, reduce the circumference and area of large and small starch grains in endosperm cells, and thus reduce the length, width and thickness of wheat caryopsis, resulting in the decrease of grain storage capacity of wheat, and ultimately reduce wheat grain weight.
Wheat; Late spring coldness; Superior and inferior; Caryopsis; Amyloplast
10.3969/j.issn.1000-6362.2023.10.006
收稿日期:2022−10−26
安徽省高校自然科学研究项目(2023AH051016);安徽省自然科学基金(2008085QC122);淮北市重大科技专项(HK2021013);安徽省重大科技专项(202003b06020021);“十四五”安徽省现代农业产业技术体系建设专项资金
通讯作者:陈翔,讲师,主要从事作物生理生态研究,E-mail:cxagricultural@163.com;李金才,教授,主要从事作物生理生态研究,E-mail:ljc5122423@126.com
翁颖,E-mail:wengying0918@163.com
翁颖,王捧娜,于敏,等.药隔期倒春寒对小麦颖果发育的影响[J].中国农业气象,2023,44(10):929-942
