基于稳定同位素混合模型的热带东太平洋4种鲨鱼摄食差异
2023-10-19张嘉旭李泽政沈永富李云凯
张嘉旭,李泽政,沈永富,李云凯,2,3*
(1.上海海洋大学 海洋科学学院,上海 201306;2.农业农村部远洋与极地渔业创新重点实验室,上海 200009;3.大洋渔业资源可持续开发教育部重点实验室,上海 201306)
大洋中上层鲨鱼多为海洋生态系统中的顶级捕食者,通过下行效应(‘top-down’ control)影响低营养级物种的种群结构和数量[1]。鲨鱼具有生长缓慢、成熟期晚和繁殖率低等生活史特征,极易受到人为因素的影响造成死亡率增加[2]。在资源相对贫瘠的开阔大洋,鲨鱼正面临着过度捕捞、气候变化和栖息地丧失等诸多威胁[3],其中,锤头双髻鲨(Sphyrnazygaena)、大眼长尾鲨(Alopiassuperciliosus)和浅海长尾鲨(Alopiaspelagicus)被列入《濒危野生动植物种国际贸易公约》(CITES)附录Ⅱ,其国际贸易受到限制,镰状真鲨(Carcharhinusfalciformis)也被国际自然保护联盟(IUCN)列为易危物种。研究这些濒危鲨鱼物种的摄食习性对于理解鲨鱼种间营养相互作用、食物资源利用模式和食物网结构等具有重要的指示作用,可为制定科学的渔业监管措施提供理论依据。
稳定同位素混合模型(bayesian stable isotope mixing models,MixSIAR)是基于狄利克雷分布(dirichlet allocation),在贝叶斯框架下构建的一个逻辑先验分布模型,综合考虑了同位素的分馏作用和时空变异性等因素,可精确量化各饵料生物对捕食者的贡献比重[4]。该方法可避免在用胃含物分析大洋性物种食性过程中常出现的取样困难、空胃率高等问题[5],即使胃含物样本量较少,基于“取样-重要性-再取样”(sampling-importance-resampling,SIR)的运算法则,也可计算出准确率较高的食源贡献率[6],从而揭示鲨鱼种间和个体水平上的摄食差异。如Niella等[7]利用MixSIAR贝叶斯混合模型研究发现,低于2.5龄的公牛鲨幼鲨偏好捕食以颗粒有机物(particulate organic matter,POM)为碳源的猎物,而4.0~6.5龄的幼鲨则主要以栖息于盐沼湿地的物种为食,表明其食物来源及摄食习性随体长而改变。Madigan等[8]基于贝叶斯稳定同位素混合模型对比分析了长鳍真鲨(Carcharhinuslongimanus)肌肉和血浆的贡献率,结果表明,其在短期内摄食大型远洋硬骨鱼类较多,而较长时间尺度下大型远洋硬骨鱼的贡献率相对较低,表明长鳍真鲨存在时间和空间上的摄食差异。
本研究中,以东太平洋常见的4种鲨鱼为研究对象,测定了鲨鱼肌肉组织及其胃含物中饵料生物的δ13C和δ15N值,通过MixSIAR贝叶斯混合模型评估各饵料类群对鲨鱼的贡献率,分析了同种鲨鱼性别间和性成熟前后的摄食差异及不同种鲨鱼间的资源分配模式,以期为进一步掌握热带东太平洋鲨鱼种间关系及构建食物网营养通道提供参考资料。
1 材料与方法
1.1 材料
试验样品为2019年10月—2020年1月在东太平洋热带海域作业的延绳钓渔船兼捕渔获,渔船作业区间为107°W~115°W、17°S~6°N(图1)。共采集4种中上层鲨鱼119尾,其中,大眼长尾鲨31尾,浅海长尾鲨30尾,锤头双髻鲨26尾,镰状真鲨32尾。
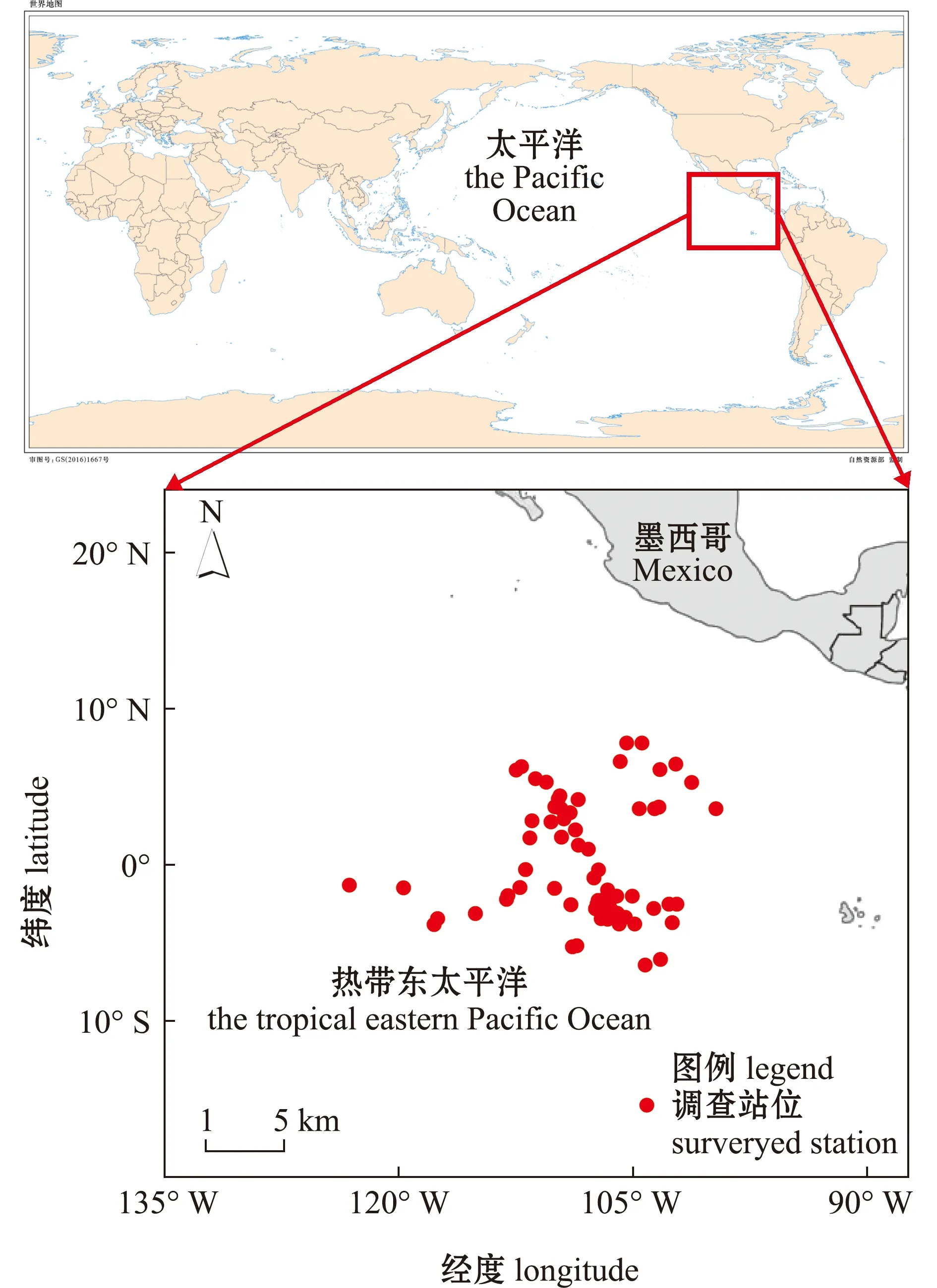
本图以自然资源部标准地图服务网站GS(2016)1667号标准地图为底图,底图边界无修改。The figure is based on the standard map GS(2016)1667 in the Standard Map Service website of Ministry of Natural Resources of the People’s Republic of China,with no modifications of the boundaries in the standard map.图1 热带东太平洋4种鲨鱼采样点Fig.1 Sampling sites of four shark species in the tropical eastern Pacific Ocean
1.2 方法
1.2.1 样品的处理 测量每尾鲨鱼的尾柄长,即上颌前端到尾柄处的长度,并通过观察鲨鱼是否具有鳍脚鉴定其性别。根据Walker[9]提出的修正成熟度指数将鲨鱼成熟期分为性成熟前后两个生长阶段,雄性成熟度取决于Flipper指数(C=1~3),雌性成熟度取决于卵巢状态(U=1~5)。从鲨鱼背鳍附近采集肌肉样本,并现场解剖鲨鱼取其胃含物。将所有样品编号装袋后于-20 ℃冷冻保存,运回实验室后于-80 ℃超低温冰箱中冷冻保存。
针对热带东太平洋4种鲨鱼及其饵料生物的肌肉样本,用去离子水清洗数次以去除尿素,而后将其装入5 mL离心管,置于冷冻干燥机内冻干24 h。由于样本中脂类的存在会使得蛋白质、碳水化合物等成分的δ13C值产生6‰~8‰的误差[10],故需将样本先进行脱脂处理,即取12 mL二氯甲烷-甲醇溶液(二者体积比为2∶1)加至盛有约2.0 mg样品的15 mL离心管中,静置20 h后置入离心机以6 000 r/min离心10 min,该过程重复3次。剩余沉积物在62 ℃烘箱中烘干以去除残留溶剂[11]。
1.2.2 饵料生物鉴定 胃含物经解冻后用吸水纸吸干多余水分,拍照鉴定胃中食物团的饵料生物种类,尽可能将其归入最小的分类单元。依据胃内容物中饵料生物的形态等特征,并参考已收集的资料对饵料残骸进行鉴定。对于肉眼观察无法鉴别的饵料生物,取适量的食物块,多次用无菌水冲洗组织表面残留的胃液,取9 mg左右样品置于灭菌离心管中,将组织剪碎,采用海洋动物组织基因组DNA提取试剂盒(天根生化科技有限公司)提取胃含物基因组DNA。
用一对通用引物COⅠ-FishF1(5′TCAACCAACCACAAAGACATTGGCAC 3′)COⅠ-FishR1(5′TAG- ACTTCTGGGTGGCCAAAGAATCA 3′)[12],选取线粒体基因细胞色素C氧化酶亚基 Ⅰ (mitochon-drial cytochrome oxidase subunit Ⅰ,COⅠ)作为分子标记用于PCR扩增。扩增体系(25 μL):ddH2O 6.5 μL,Premix Ex Taq 酶(TaKaRa)(2×)12.5 μL,正、反向引物(0.25 μmol/L)各0.5 μL,DNA模板5 μL。反应条件:95 ℃下预变性2 min;95 ℃下变性30 s,60 ℃下退火45 s,72 ℃下延伸1 min,共进行30个循环;最后在72 ℃下再延伸5 min,4 ℃下结束反应。DNA测序结果在NCBI (National Center for Biotechnology Information)网站进行序列比对,通过DNA分子鉴定饵料生物物种。
1.2.3 稳定同位素分析 将干燥后的鲨鱼和饵料生物的肌肉样本研磨成粉,取约1.5 mg并用锡箔纸包装,利用稳定同位素质谱仪(IsoPrime 100,英国)和元素分析仪(Vario ISOTOPE Cube,德国)测定碳、氮稳定同位素比值,以符号δ表示。δ13C和δ15N值的计算公式为
δX=(Rsa/Rst-1)×1 000。
其中:X为13C或15N;Rsa和Rst分别为样品和标准样品的同位素比值。δ13C的国际标准物为维也纳国际原子能中心制作的美洲拟箭石(vPDB),δ15N的国际标准物为大气中的氮气。每隔20个样品插入3个实验室标准品(蛋白质:δ13C=-26.98‰,δ15N=5.96‰)校正同位素数据,碳、氮稳定同位素比值的分析精度分别为0.15‰和0.20‰。
1.2.4 Kruskal-Wallis检验和MixSIAR贝叶斯混合模型分析 根据饵料鉴定结果,把饵料生物按照分类地位相似的原则合并成同一个食物源,经多元方差分析,3类潜在食物来源表现出一种或一种以上的稳定同位素显著差异(δ13C:P<0.05),且共线性特征不显著,适合进一步分析捕食者的食物来源[13]。采用Kruskal-Wallis检验分析4种鲨鱼种间食物源,以及同种鲨鱼性别间和性成熟前后的δ13C和δ15N值是否存在显著性差异。利用MixSIAR贝叶斯混合模型分析4种鲨鱼各食物源的贡献率,将模型中的营养富集因子(trophic discrimination factors,TDFs)Δ15N值设置为3.7‰±0.5‰,Δ13C值设置为1.7‰±0.5‰[14],将每类食物源的δ13C和δ15N值数据集输入到MixSIAR贝叶斯混合模型中。
在运行MixSIAR贝叶斯混合模型前,采取混合多边形迭代模拟的方法判别数据质量。4种鲨鱼的同位素数据绝大多数落在几种食物来源确定的混合多边形内,表明食物源分类及设置的TDFs值合理。MixSIAR贝叶斯混合模型在300 000次马尔科夫链蒙特卡洛法(MCMC)模拟下运行,且经Gelman-Rubin(所有变量均<1.01)和Geweke诊断后实现收敛,获得各饵料源对鲨鱼食物比例的概率分布数据。
1.3 数据处理
稳定同位素试验数据均采用平均值±标准差(mean±S.D.),使用R 4.1.3和ArcGIS 10.8软件进行数据处理和作图,显著性水平设为0.05。
2 结果与分析
2.1 胃含物组成
4种鲨鱼的胃含物共鉴定出饵料生物21种,其中,小型硬骨鱼7种,头足类11种,大型硬骨鱼3种(表1)。
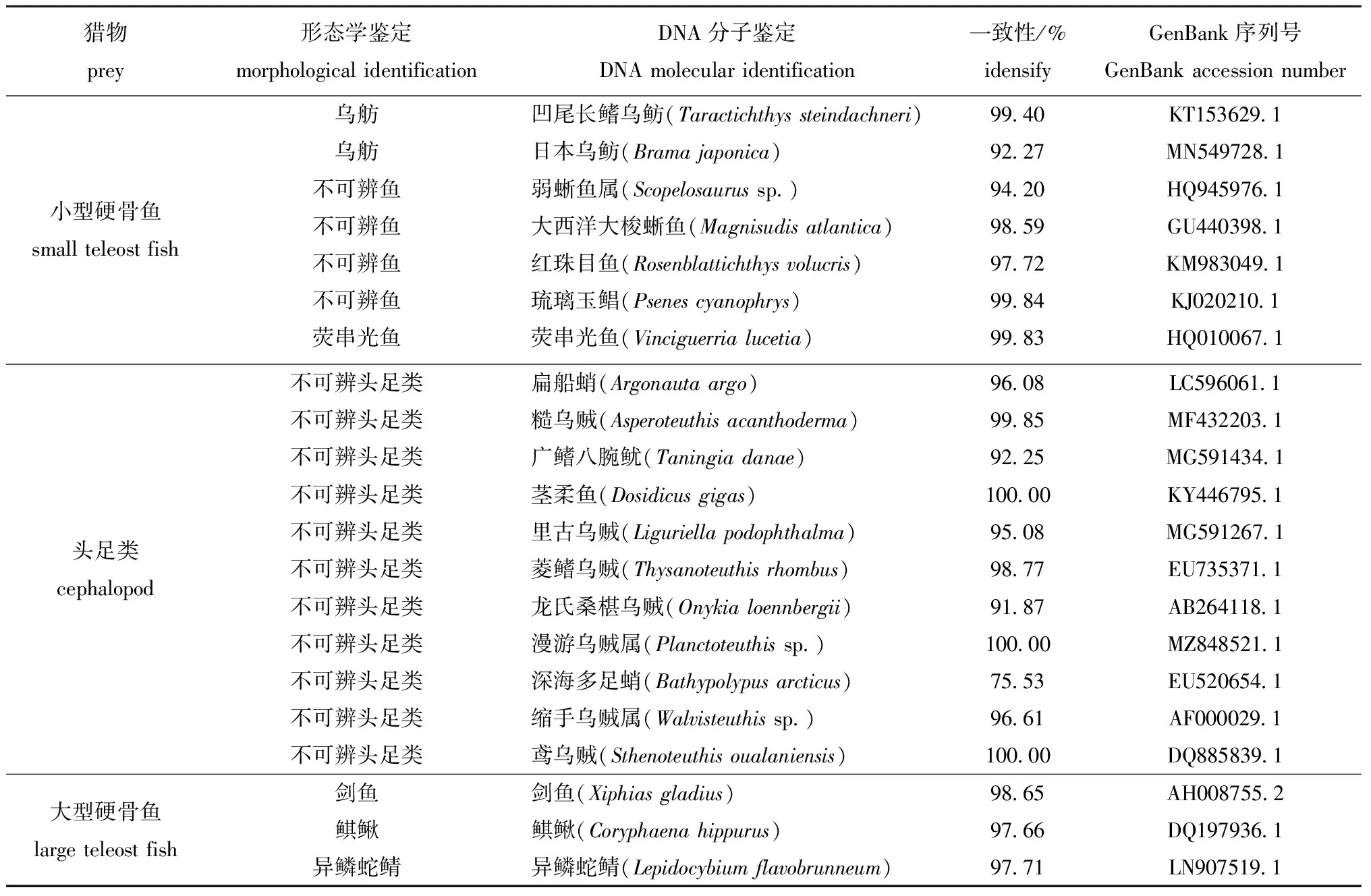
表1 DNA分子鉴定与形态学鉴定结果Tab.1 Comparison of partial morphological identification results with DNA molecular identification
2.2 肌肉碳、氮稳定同位素值特征
总体上,4种鲨鱼肌肉的δ13C和δ15N值均具显著性差异(P<0.05),锤头双髻鲨δ13C和δ15N值较高,其次为镰状真鲨、大眼长尾鲨和浅海长尾鲨(表2)。4种鲨鱼肌肉的δ13C和δ15N值性别间均无显著性差异(P>0.05);4种鲨鱼肌肉δ13C值性成熟前后均无显著性差异(P>0.05),除镰状真鲨性成熟后肌肉δ15N值显著低于性成熟前(P<0.05)外,其余3种鲨鱼肌肉δ15N值性成熟前后均无显著性差异(P>0.05)(表3)。
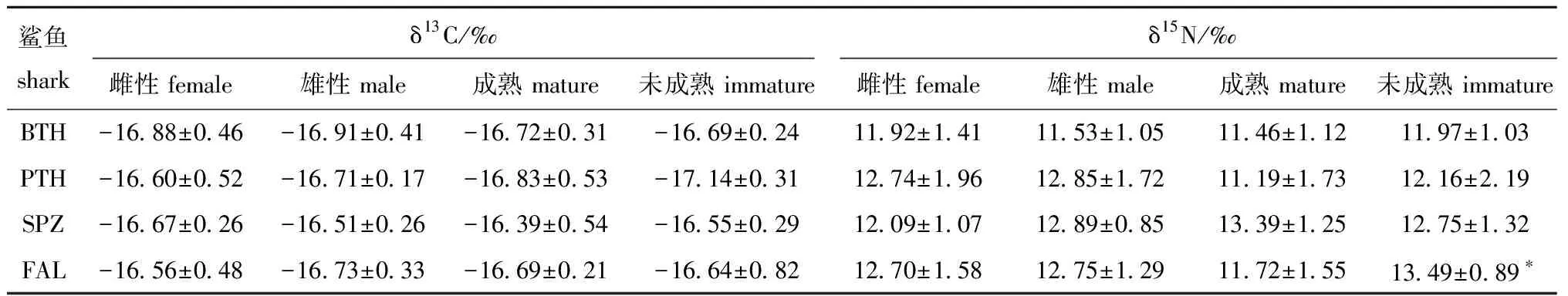
表3 热带东太平洋4种鲨鱼不同生长阶段及性别间肌肉的碳、氮稳定同位素比值
3类主要潜在饵料群中,饵料生物δ13C值为-19.70‰~-16.71‰,δ15N值为5.61‰~10.26‰,其中,小型硬骨鱼类的δ13C和δ15N值最低,分别为-19.70‰~-18.16‰和5.61‰~6.90‰,其次为头足类,δ13C和δ15N值分别为-19.20‰~-17.11‰和6.39‰~8.22‰,大型硬骨鱼类的δ13C和δ15N值最高,分别为-17.66‰~-16.71‰和8.65‰~13.26‰(表2,图2)。
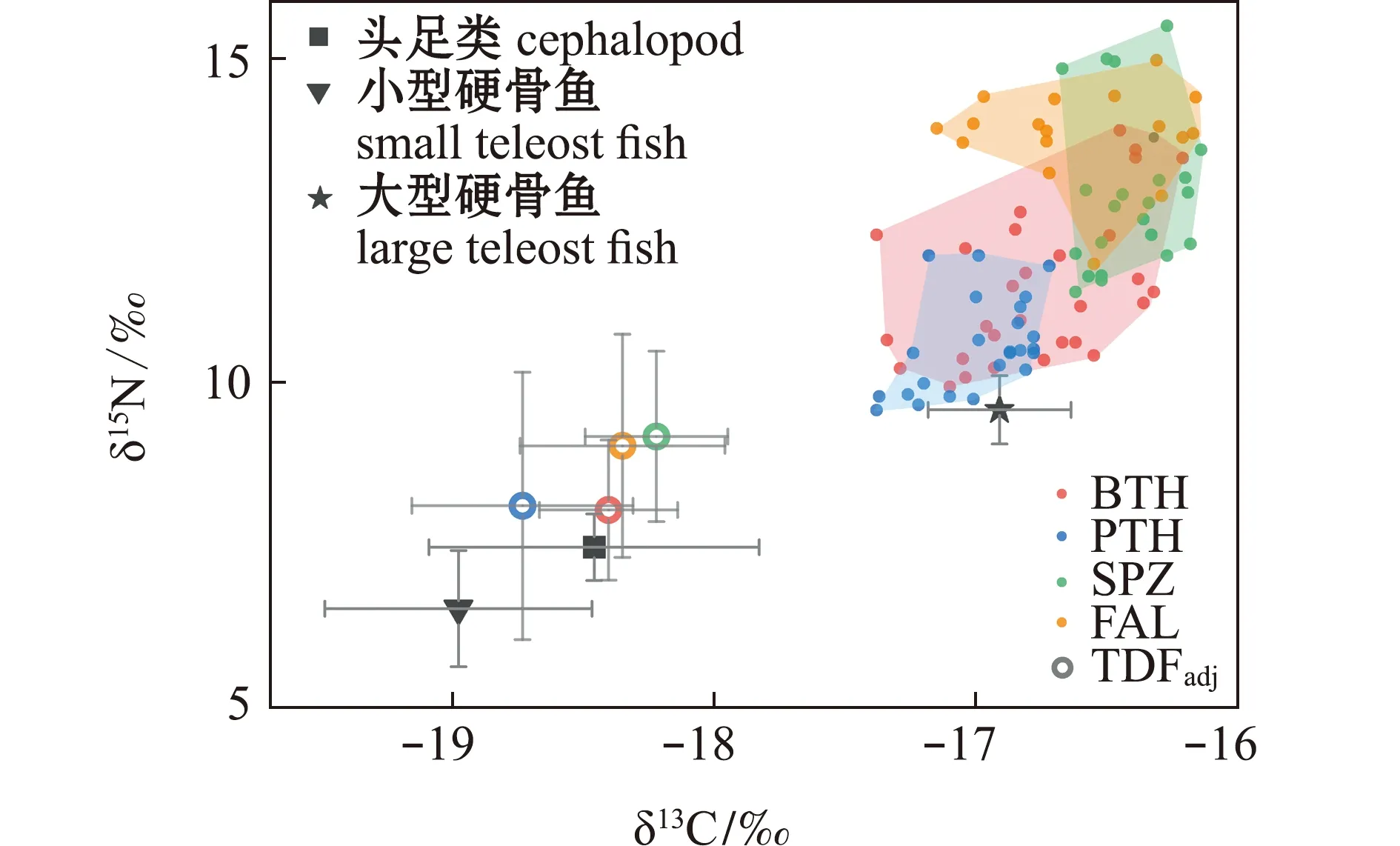
BTH—大眼长尾鲨;PTH—浅海长尾鲨;SPZ—锤头双髻鲨;FAL—镰状真鲨。BTH—Alopias superciliosus;PTH—Alopias pelagicus;SPZ—Sphyrna zygaena;FAL—Carcharhinus falciformis.图2 营养富集因子校正后的稳定同位素混合空间Fig.2 Stable isotope mixing space corrected by trophic enrichment factors
2.3 食源贡献率
2.3.1 种间摄食差异 MixSIAR模型结果显示:大眼长尾鲨和浅海长尾鲨主要捕食头足类,其贡献率分别为72.6%和59.0%,大型硬骨鱼的贡献率均较低,分别为9.7%和2.8%;对于锤头双髻鲨,头足类贡献率最高(43.5%),其次是大型硬骨鱼(35.2%),小型硬骨鱼贡献率最低(21.3%);对于镰状真鲨,头足类的贡献率最高(59.3%),其次是大型硬骨鱼(22.1%),小型硬骨鱼最低(18.6%)(图3)。
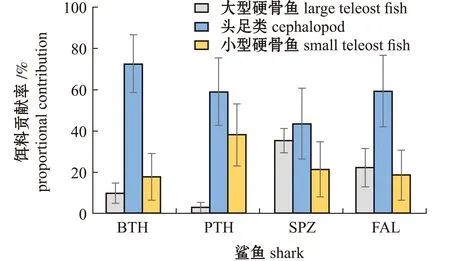
BTH—大眼长尾鲨;PTH—浅海长尾鲨;SPZ—锤头双髻鲨;FAL—镰状真鲨。BTH—Alopias superciliosu;PTH—Alopias pelagicu;SPZ—Sphyrna zygaena;FAL—Carcharhinus falciformi.图3 热带东太平洋4种鲨鱼的食源贡献率Fig.3 Diet proportion of four shark species in the tropical eastern Pacific Ocean
2.3.2 性别间摄食差异 MixSIAR模型结果显示,头足类对于大眼长尾鲨雌鲨和雄鲨的贡献率均最高,分别为63.1%和59.0%;对于浅海长尾鲨,头足类和小型硬骨鱼对雄鲨的贡献率相对较高,均为47%,头足类对雌鲨的贡献率最高(55.7%),其次为小型硬骨鱼(39.3%)和大型硬骨鱼(5.0%);对于锤头双髻鲨,大型硬骨鱼对雌鲨的贡献率最高(45.3%),头足类对雄鲨的贡献率最高(46.1%),小型硬骨鱼对雌鲨和雄鲨的贡献率均最低,分别为17.3%和16.2%;镰状真鲨性别间饵料贡献率存在较大差异,大型硬骨鱼对雌鲨和雄鲨的贡献率分别为19.0%和26.4%,头足类贡献率分别为59.9%和39.9%,小型硬骨鱼贡献率分别为21.1%和33.7%(表4)。

表4 热带东太平洋4种鲨鱼不同性别的食源贡献率
2.3.3 不同生长阶段摄食差异 大眼长尾鲨性成熟前后的饵料贡献率差异较小;锤头双髻鲨性成熟前头足类贡献率最高(46.1%),性成熟后大型硬骨鱼贡献率最高(38.9%);浅海长尾鲨性成熟前后头足类的贡献率均大于小型硬骨鱼和大型硬骨鱼,性成熟前后大型硬骨鱼贡献率均较低;镰状真鲨性成熟前后存在明显的摄食转变,性成熟前大型硬骨鱼和小型硬骨鱼的贡献率均为45%左右,头足类贡献率较小,而性成熟后头足类贡献率升高(56.1%),其次是小型硬骨鱼贡献率为26.1%,大型硬骨鱼的贡献率最低(表5)。

表5 热带东太平洋4种鲨鱼性成熟前后的食源贡献率
3 讨论
3.1 鲨鱼种间摄食差异
本研究中,利用MixSIAR贝叶斯混合模型分析了热带东太平洋4种鲨鱼的食物来源贡献比,镰状真鲨和锤头双髻鲨的饵料贡献率从高到低依次为头足类>大型硬骨鱼>小型硬骨鱼,大眼长尾鲨和浅海长尾鲨的饵料贡献率依次为头足类>小型硬骨鱼>大型硬骨鱼,但头足类对浅海长尾鲨的饵料贡献率低于大眼长尾鲨(图3)。尽管大洋性鲨鱼均广泛分布于热带和亚热带海域[15],但不同鲨鱼物种可能具有各自的栖息地偏好。锤头双髻鲨更偏好栖息于热带和温带的沿岸和远洋水域,常于近岸及浅海底层等水域摄食[16];镰状真鲨则广泛分布于大陆架边缘,相较于锤头双髻鲨更具有大洋性特征[17]。浅海长尾鲨白天更偏好栖息于200~300 m水层,夜晚洄游至50~150 m的上层水域[18];大眼长尾鲨栖息水层更深,白天多栖息于超过300 m深的水层,夜晚洄游至10~130 m的水域[18]。两种长尾鲨显著的昼夜垂直迁移习性相较于锤头双髻鲨和镰状真鲨栖息水深变化范围更大,栖息水层的重合使其摄食头足类和小型硬骨鱼类等饵料生物更多。此外,大眼长尾鲨具有发育良好的奇网(rete mirabile)系统[19],可以维持颅内温度和提升视力,提高其在冷水中的觅食能力,从而可捕食更多糙乌贼、菱鳍乌贼等深海猎物。
研究表明,两种长尾鲨和两种真鲨的捕食方式不同[20-21]。长尾鲨游泳速度较慢,常利用尾鳍击晕鱼群以完成捕食行为[20],其饵料生物多以体型较小的猎物为主,而镰状真鲨利用高速移动的游泳能力捕捉猎物[21],比长尾鲨更易捕食游泳能力较强的大型硬骨鱼。González-Pestana等[22]对浅海长尾鲨进行胃含物分析发现,茎柔鱼相对多样性指数(IRI)最高,其次是小型硬骨鱼[沙丁鱼(Sardinepilchardus)、智利无须鳕(Merlucciusbilinearis)等],大型硬骨鱼较少。Preti等[19]也发现,大眼长尾鲨饵料鱼以太平洋无须鳕和手钩鱿属(Gonatussp.)等小型硬骨鱼和头足类为主。而镰状真鲨猎物饵料鱼则以头足类和大型硬骨鱼居多,如Galván-Magaa等[17]发现,太平洋海域的镰状真鲨捕食头足类最多,其次为大型硬骨鱼,小型硬骨鱼最少。此外,大型硬骨鱼对长尾鲨的贡献率较低,也可能是由于本研究中两种长尾鲨多为中小型个体,故捕食大型硬骨鱼的成功率较低[23]。
同种鲨鱼对猎物的喜好程度因海域而异,在沿岸海域,甲壳类也可作为镰状真鲨的优势饵料种类[24]。如加利福尼亚西海岸镰状真鲨的胃含物中红蟹(Pleuroncodesplanipes)的IRI值高达83%,远高于头足类和硬骨鱼类[24]。秘鲁沿岸海域锤头双髻鲨以摄食头足类为主(IRI>80%),其次捕食硬骨鱼类(IRI<10%)[25],与本研究中结果存在一定差异。这可能是由于胃含物分析的结果反映其短期的摄食情况[26],受环境的影响较大,而肌肉稳定同位素则反映较长时期的摄食信息[27]。鲨鱼作为洄游性物种,在所栖息海域利用丰度较高的饵料生物,导致胃含物分析与稳定同位素混合模型结果出现差异。受厄尔尼诺-南方涛动(ENSO)影响,饵料生物的种类、数量和分布情况会发生变化,鲨鱼由于其自身代谢和能量需求也会进行相应的生理和行为调整,因此,造成两种方法的评价结果出现差异[28]。
3.2 鲨鱼性别间摄食差异
鲨鱼的优势饵料群体存在性别差异[21],本研究结果也证实了这一观点。Estupián-Montao等[29]发现,厄瓜多尔海域头足类对锤头双髻鲨雄鲨的相对重要性指数大于雌鲨,雄鲨更偏好捕食远洋猎物,而雌鲨生产前会洄游至沿岸海域。Xu等[30]发现,雌性浅海长尾鲨肝脏中的C22:0+C24:0等特征脂肪酸较高,表明雌鲨具有较高的陆源食物来源和近岸特征,而雄鲨肝脏中C18:1n9含量较高,表明其通常栖息于较深层水域[31]。在身体结构方面,一般来说雌鲨体型相对雄鲨较大,故需要更多能量来维持自身的代谢活动[27]。雌性镰状真鲨在怀孕期间所需能量增加,在怀孕前会储存更多繁殖所需的能量[26,30],因此,多种原因造成鲨鱼性别间的摄食差异。
3.3 鲨鱼不同生长阶段摄食差异
对多数鲨鱼物种的研究证实,其捕获猎物的数量和多样性与体型有关[32],捕获猎物的能力与感觉器官的发育程度也有关,随着鲨鱼体型的增加,其口裂逐渐发育完备,咬合能力显著增强,对较大猎物的捕食成功率增加[23]。此外,随着鲨鱼的生长发育,大型鲨鱼需摄食更高能量的猎物以满足自身代谢、繁殖和洄游等生命活动的需求[23],从而造成个体间摄食差异。有研究表明,鲨鱼在不同的生长阶段所偏好的栖息环境不同[30],多种鲨鱼性成熟后偏好生活在远洋水域,繁殖期洄游至沿海水域进行交配。Gonzalez-Pestana等[25]通过胃含物分析发现,东太平洋锤头双髻鲨幼鲨以近岸物种如鞭乌贼属(Mastigoteuthis)、巴塔哥尼亚枪乌贼(Doryteuthisgahi)和低鳍鲳属(Peprilussp.)等为食,而成年鲨鱼多捕食具有大洋性的茎柔鱼和鱼钩乌贼等。本研究中,镰状真鲨性成熟后饵料生物大型硬骨鱼的贡献率显著降低,也可能与其独特的15N富集机制有关。本研究中,稳定同位素结果显示,镰状真鲨肌肉的δ15N值在性成熟后显著降低,由于镰状真鲨幼鲨偏好栖息于海洋中层或深层海域,依赖中层悬浮颗粒物为基线的食物网,因此,所处生态系统的基线δ15N值较高[33]。Li等[33]亦发现,随镰状真鲨的生长发育其摄食区域发生变化,体型较大的镰状真鲨肌肉组织的δ15N值较低。这可能是本研究中模型输出结果表现为镰状真鲨性成熟后大型硬骨鱼贡献率降低的原因之一。
4 结论
1)利用MixSIAR贝叶斯混合模型计算了3类主要饵料群体对4种鲨鱼的饵料贡献率,初步分析了4种大洋性鲨鱼的摄食习性。4种鲨鱼的主要饵料生物均为头足类,小型硬骨鱼和大型硬骨鱼的贡献率存在种间差异,表明同海域鲨鱼的资源利用模式存在差异。
2)除大眼长尾鲨外,浅海长尾鲨、锤头双髻鲨和镰状真鲨在性别间和性成熟前后饵料贡献率存在差异,表明大洋性鲨鱼在个体发育过程中存在食性转变,主要与栖息环境和个体能量需求等有关。
3)为进一步了解大洋性鲨鱼在同海域内的资源分配模式,更具准确性地分析大洋性鲨鱼的摄食习性,建议将胃含物分析与稳定同位素分析方法相结合以探究大洋性鲨鱼的摄食生态。
