黑龙江丰林国家级自然保护区红松+紫椴+硕桦群系优势树种生态位特征与种间联结分析
2023-10-18白晓航施佳颖
白晓航 施佳颖
(1.东南大学建筑学院,江苏省城乡与景观数字技术工程中心,南京 210096;2.云南省林业和草原科学院,国家林业和草原局珍稀濒特森林植物保护和繁育重点实验室,云南省森林植物培育与开发利用重点实验室,昆明 650204;3.北京师范大学地理科学学部,地表过程与资源生态国家重点实验室,北京 100875)
阔叶红松林是中国东北东部山区地带性森林植被,具有丰富的物种组成与独特的建群种,在保持水土、防风固沙、气候调节、涵养水源、维持生态平衡和生物多样性等方面具有重要意义。目前,关于阔叶红松混交林的研究主要集中在群落结构、与土壤相互作用、对环境因子的适应策略、生物多样性共存机制等宏观层面,对特定群系类型的阔叶红松混交林的生态位及其种间关系的基础研究仍然缺乏。以黑龙江丰林国家级自然保护区内紫椴+硕桦+红松群系为研究对象,通过对该区域全面踏查设置植物群落调查样地,以期揭示该群系的优势树种生态位特征与种间联结。结果表明:建群种红松、硕桦、紫椴的生态位宽度较大,群系结构相对稳定,随着优势树种重要值的增加生态位宽度逐渐增加。群系中不同树种的生态位重叠有差异、呈网状结构,优势树种间的生态位分化程度较好。群系中优势树种整体上呈现显著正联结,正联结的种对数大于负联结的种对数,优势树种间正相互作用占优势,正相互作用的强度较低,群系群落结构相对稳定、向正向演替发展。研究揭示了以红松+紫椴+硕桦群系为主的阔叶红松林内优势树种生态位特征和不同树种的资源利用能力,为该区域阔叶红松混交林保护和可持续经营管理提供理论依据。
生态位;种间联结;自然保护区;优势种;阔叶红松混交林
阔叶红松林是中国东北温带针阔混交林区地带性植被,具有丰富的物种多样性和群落组成,这片林区为中国北方木材产业提供支持,并作为中国生态战略格局的重要组成部分,在保持水土、气候调节、涵养水源以及维持区域生物多样性等方面具有不可替代性[1-2]。阔叶红松林分布区的形成与第三纪以来海面的陷落及冰期时代和间冰期时代的环境变迁有关,其分布区属于暖温带季风气候,地形复杂、物种丰富,在中国主要分布于40°45′-49°20′N,124°45′-135°01′E之间,具有林分结构复杂、物种多样性丰富等特征,林内保留着红松(Pinuskoraiensis)、水曲柳(Fraxinusmandshurica)、黄檗(Phellodendron amurense)、胡桃楸(Juglansmandshurica)、蒙古栎(Quercusmongolica)、紫椴(Tiliaamurensis)等珍贵树种,又是东北虎、远东豹、棕熊、金雕等多种濒危野生动物的栖息地[3]。多个共享相似资源的物种如何在局域群落内共存一直是学者关注的问题,经典物种共存理论认为,当稳定的种间生态位差异大于适合度差异时,才可能发生稳定的物种共存;而生态位理论认为生境过滤和竞争排斥等生态位过程是物种共存主要驱动力[4-5]。高维生态位假说假定个体之间的竞争通常发生在多维资源生态位上,但通常用少数几维生态位空间上种内和种间竞争强度的平均值来解析种内和种间的作用[6]。生态位差异是物种共存的基本条件之一,探究阔叶红松混交林的群落结构与生态特征对其保护与可持续经营具有重要意义。
群落物种生态位和种间联结性是了解群落结构变化和演替的基础,通过物种的生态位和种间联结特征,可以更好地了解不同物种在群落中对有限资源的竞争关系以及它们之间稳定共存的关系[7]。前人对小兴安岭阔叶红松林群落优势树种的生态位进行分析,结果表明阔叶红松林群落优势树种对资源的利用较为充分,乔木层优势树种种间负相互作用占优势但强度较低[8-9]。生态位互补效应认为,在多样性较高的群落中,物种以互补的方式利用有限资源,从而促进了生态系统功能,这种多样性使得群落中的物种能够在资源利用上互相补充,提高整体效率[10];生物量比率效应则认为,物种对生态系统功能的影响取决于它们的特征及在群落中的相对比率,例如,某些物种可能在生物量或个体数量上占主导地位,它们对生态系统功能产生更显著的影响[11];植被数量效应认为,生态系统功能主要取决于单位面积内森林所具有的生物量或蓄积量,而非仅仅依赖于功能多样性或功能组成,生物量的增加通常与更高的生态系统功能相关联[12]。前人对阔叶红松林功能多样性与森林碳汇功能关系研究发现生物多样性与碳汇功能的关系格局是生态位互补效应、生物量比率效应和植被数量效应等多种生态学机制共同作用的结果[13]。
紫椴+硕桦+红松群系是阔叶红松混交林典型群系代表类型之一,具有丰富的物种组成,生态功能稳定,是研究阔叶红松混交林群落结构与功能的典型群系类型。目前,关于东北阔叶红松混交林的研究主要集中在群落结构、与土壤相互作用、对环境因子的适应策略、生物多样性共存机制等宏观层面,对特定群系类型的阔叶红松混交林的生态位及其种间关系的基础研究仍然缺乏[14-17]。鉴于此,本研究以黑龙江丰林国家级自然保护区内紫椴+硕桦+红松群系为研究对象,对该区域全面踏查并设置植物群落样地,拟解决科学问题如下:以红松+紫椴+硕桦群系为主的红松阔叶混交林内优势树种的生态位和种间联结如何,该群系的资源利用能力有何特点?本研究旨在揭示以红松+紫椴+硕桦群系为主的阔叶红松林内优势树种生态位特征,分析红松种群与其他乔木种群占据空间的范围和资源利用能力,为该区域阔叶红松混交林的可持续经营与管理提供理论依据。
1 研究区域与样地设置
1.1 研究区域概况
黑龙江丰林国家级自然保护区位于黑龙江省伊春市丰林县境内,地理坐标为128°58′-129°15′E,48°02′-48°12′N。该保护区始建于1958年,主要的保护目标是以北温带针阔叶混交林生态系统,以红松为主,以及珍稀野生动植物资源,其于1997年被联合国教科文组织纳入世界生物圈保护区网络,属森林生态系统类型自然保护区,东西长20 km,南北宽16 km,总面积18 165.4 hm2,是中国北方红松林中最具代表性的国家级自然保护区[18]。该区域具有寒温带半湿润气候,年降水量650 mm,年均温-0.5℃,无霜期120 d左右。保护区内山体海拔280~683 m,坡度10°~25°,植被覆盖率达90%以上,属于中国东北区长白植物区系小兴安岭亚区,形成温带针阔叶混交林。本研究区域的阔叶红松混交林以红松为优势种,伴生有硕桦(Betulacostata)、紫椴(Tiliaamurensis)、水曲柳、白桦(Betulaplatyphylla)、色木槭(Acer pictum)等温性阔叶树种以及红皮云杉(Picea koraiensis)、臭冷杉(Abiesnephrolepis)等寒温性树种等。
1.2 样地设置
于2021年和2022年的7-9月对黑龙江丰林国家级自然保护区进行全面踏查,在研究区内选取72块红松+紫椴+硕桦群系典型样地(30 m×30 m)。记录每块样地的经度、维度、海拔高度、温度、湿度,调查样地中所有胸径大于2 cm的木本植物的树高、胸径、相对位置、冠幅和生长状态,并采集样方内凋落物及土壤,带回实验室测量相应指标。
2 数据处理与统计分析
2.1 生态位宽度与重叠
重要值能够反映植物在群落种的作用和地位,通过相对优势度、相对多度、相对高度计算乔木层重要值IV[19],计算见公式(1)。
生态位宽度是指某一物种能够利用的所有资源的总和,反映了该物种在群落中的地位和资源利用策略、与其他物种的竞争关系和资源分配情况,可以量化并衡量某一物种对水分、光照等各种资源的利用程度,以及对环境的适应能力,这些信息对于生态系统的稳定性和群落结构的理解具有重要意义。本研究选用Levins指数(BL)描述群系优势树种的生态位宽度,具体计算见公式(2)。式中,Pij是某一物种i在资源位j上的重要值与某一物种i在某一范围内所有资源位上的重要值总和之比,r是在这一范围中资源位的总数[20]。
阔叶红松林群落优势树种生态位重叠用,据Pianka指数(Oik)描述,它可以计算种对间的生态位重叠,具体计算见公式(3)。式中,Oik为第i个物种和第k个物种的生态位重叠指数,数值范围为0 ~1,数值越大表示重叠度越高,其中,pij是物种i在资源j上的重要值,pkj分别为物种k在资源j上的重要值[21]。
2.2 联结性与关联性
总体联结性是指植物群落中所有物种之间的关联程度,本研究采用Schluter的方差比(VR)检验方法来验证阔叶红松林优势树种之间的整体关联程度,并检验相关系数的显著性,具体计算见公式(4)。式中,N是研究区域中样方的总数,S是群落中物种的总数,Tj是第j个样方中的物种数,t为样方中的平均物种数,ni是i物种的样方数。在独立性假设的情况中,当VR=1,表示物种之间总体上没有关联性;当VR>1时,物种之间总体上呈现出正联结;当VR<1时,物种之间整体上呈现出负联结[22]。
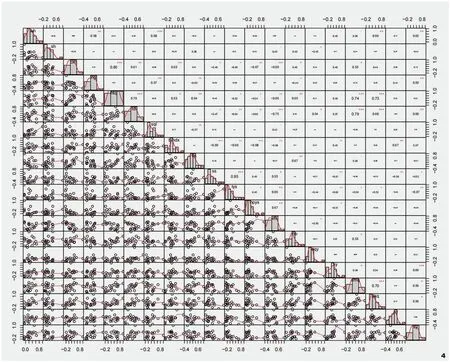
物种之间的正联结和负联结有时可能会出现相互抵消的现象,因此,需要统计W(W=N×VR),从而核验VR值与1之间的偏离程度。当X20.05(N) 种成对物种间的联结性选用红松+紫椴+硕桦群系中20个优势树种,建立种对间的2×2列联表,统计a、b、c、d的值。采用卡方检验X2(Yate’s correction formula,X2)和联结系数(AC)测定群系中成对物种间的联结性,计算见公式(5)。在一个样本总数为N的研究中,有两种物种A和B同时共存的样本数为a,有B物种存在而A物种不存在的样本数为b,有A物种存在而B物种不存在的样本数为c,同时,有A和B两种物种都不存在的样本数为d。当3.841<X2<6.635时,物种对之间存在联结;当X2>6.635时,物种对之间表现为显著联结;当ad>bc时,物种对间联结为正,联结系数AC=(ad-bc)/[(a+b)(b+d)];当ad<bc时,物种对间联结为负,联结系数AC=(ad-bc)/[(a+b)(a+c)]。当X2<3.841时,物种对表现为不联结,联结系数AC=(ad-bc)/[(b+d)(d+c)]。联结系数AC的取值范围为[-1,1],当AC的值越接近1时,表明物种间的正关联性越大;当AC的值越接近-1,表明物种间的负关联性越大;当AC=0时,表明种间无关联性[9]。 X2检验和联结系数能够定性描述种间联结的性质和显著程度,但不能描述两个种之间的数量关系,结合Pearson相关系数(rs(i,k))来判定种间的关联程度,计算见公式(6)。式中,rs(i,k)是样方中物种i与物种k之间的Pearson相关系数,其中,N是研究中样方的总数,xij和xkj分别是物种i和物种k的多度,并且由它们构成向量xi和,xi和分别是物种i在j样方中的重要值、物种i在j样方中重要值的平均值。rs(i,k)的值在(0,1]表示正相关,即当值越接近1时,表示种i和种k之间的相关性越强;rs(i,k)的值在[-1,0)表示负相关,即当值越接近-1时,表示种i和种k之间的负相关性越强;rs(i,k)的值为0时表示不相关,即种i和种k之间没有线性相关关系[23]。 综上,生态位重叠度与重要值间关系选用stats包中的lm()函数进行回归分析并探究它们之间的关联程度;计算物种之间的总体联结性选用spa包sp.assoc()函数和sp.pair()函数,并选用X2统计量来检验是否存在显著关联;物种对间的Pearson相关性选用psych包的corr.test()函数计算,并进行显著性检验;数据分析和可视化均在R 4.0.5中进行,探究阔叶红松林优势树种之间的相互作用。 在所调查的72块样地中共有32种乔木,计算32种乔木的重要值,选取重要值排名前20的物种作为主要优势树种。重要值和生态位宽度如图1所示,20种优势树种的生态位宽度由大到小的顺序依次为红松(2.03)>硕桦(1.39)>色木槭(1.31)>红皮云杉(1.27)>紫椴(1.23)>杉松(Abies holophylla)(1.22)>青楷槭(Acertegmentosum)(1.21)>辽椴(Tiliamandshurica)(1.11)=臭冷杉(1.11)>落叶松(Larixgmelinii)(1.09)>水曲柳(1.05)=春榆(UlmusdavidianaPlanch.var.japonica)(1.05)>山杨(Populusdavidiana)(1.04)>白桦(1.02)>蒙古栎(0.99)>胡桃楸(0.93)>黄檗(0.93)>暴马丁香(Syringareticulatasubsp.amurensis)(0.88)>大青杨(Populusussuriensis)(0.85)>朝鲜槐(Maackiaamurensis)(0.71)。 图1 主要优势树种的重要值和生态位宽度Fig.1 Importance values and niche width of dominant tree species 从结果看出,红松在群落中生态位宽度和重要值均达到最高,红松长势较好,硕桦和紫椴次之,它们是该群落的建群种,群系结构相对稳定。红松、硕桦、色木槭、红皮云杉、紫椴、杉松、青楷槭等树种的生态位宽度值均在1.2以上,意味着它们能够利用多样化的资源和生态条件,在该群落中表现出更强的适应能力、更有效地利用环境中的有限资源,在乔木层中占据优势地位,具有较高的生存和繁殖成功率,在群落结构和演替过程中发挥重要的作用。暴马丁香、大青杨、朝鲜槐等树种的生态位宽度较小,意味着它们在群落中对水分、光照等资源的竞争能力较弱,可能受到其他物种的竞争压力,难以在群落中占据优势地位,这些物种可能扮演较为辅助或次要的角色,对群落的整体结构和生态过程的影响相对较小。从图2中可以看出,优势树种生态位宽度与重要值大小排序并不一致,但整体上随着重要值的增加其生态位宽度逐渐增加,红松+紫椴+硕桦群系结构仍处于动态变化之中。 图2 优势树种的重要值和生态位宽度关系Fig.2 Relationship between importance values and niche width of dominant tree species 红松+紫椴+硕桦群系的190个种对中,生态位重叠>0.31的共20对,占总对数的10.52%,说明该群落中部分优势树种的生态习性具有较高的相似性(图3)。生态位重叠>0.42的9对优势树种中,部分物种对的生态位宽度较大,例如,红松、白桦、蒙古栎、水曲柳,它们是半阳性和阳性树种,它们的生态位重叠大且生态位宽度广,其适应性较强。从生态位重叠指数看,红松与白桦(0.43)、红松与蒙古栎(0.52)、红松与水曲柳(0.52)、红松与臭冷杉(0.45),在该群系中,红松与这些树种的生态位存在重叠关系,在群落发展的过程中,对相同资源的利用会产生竞争关系。水曲柳与硕桦(0.45)、水曲柳与色木槭(0.42)、色木槭与暴马丁香(0.44)、暴马丁香与辽椴(0.45)也存在高度重叠,在该群系中,生态位重叠复杂,物种间竞争呈网络结构,不同树种生态位重叠具有较大差异。生态位重叠<0.21的有145对,占总对数的76.31%,生态位重叠值较小,这些优势树种的生境差异较大,如红松与朝鲜槐、胡桃楸、辽椴、大青杨等。从整体上看,生态位重叠值较小的种对数高于生态位重叠值大的种对数,表明在该群系中不同优势树种之间的资源利用策略和生态位差异性较为明显,不同树种之间的生态位重叠有差异,使得物种能够在群落中共存,该群系的优势树种的生态位分化程度较好。 图3 红松+紫椴+硕桦群系优势树种的生态位重叠Fig.3 Ecological niche overlap of dominant tree species in Form.Pinus koraiensis + Tilia amurensis + Betula costata 方差比率法是一种用于分析物种联结性的统计方法,用于比较联结性在不同物种之间的差异,本研究中红松+紫椴+硕桦群系的VR=1.48,表明在该群系中优势树种整体上呈现正联结,这意味着优势树种之间存在着一种正向的相互作用和资源利用关系;在卡方分布表中,X20.95(72)= 53.462,X20.05(72)=92.808,用方差比率法检验统计量的W值为132.45,不在区间(53.462,92.808)范围内,VR显著偏离1,联结性显著。因此,红松+紫椴+硕桦群系优势树种总体上表现为显著正联结。 分析优势树种之间的相互作用模式和联结性,对于揭示优势树种在群落中的地位、相互作用和共存机制有所启示。Pearson相关检验展示了190对优势树种组成,表明在优势树种之间存在不同程度的联结性。正联结的优势树种对有35对,占总种对数的18.42%,其中,极显著正联结24对,显著正联结11对;负联结的优势树种对有25对,占总种对数的13.15%,其中,极显著负联结15对,显著负联结10对;无关的优势树种对有130对,占总种对数的68.42%(图4)。这些结果进一步说明在优势树种之间存在不同程度的联结性,并且在正联结和负联结的优势树种对中,还分别有极显著和显著的相关关系。在所有种对中,种间联结性较高的种对占总种对数的20.52%,这意味着这些种对之间存在较强的正向或负向相关性。相比之下,种间联结性较低的种对只占总种对数的11.05%,这些种对之间的相关性较弱。 图4 优势树种Pearson相关系数及其显著性Fig.4 Pearson correlation coefficients and significance of dominant tree species 生态位展现了物种在群落中的时空位置以及不同种群之间的功能关系,定量反映物种之间以及物种与生境之间的相互作用关系,是物种对生境中各环境因子生态适应的结果,通常用生态位宽度和生态位重叠表征[24]。生态位宽度能够反映物种在生境中的地位和分布,展现物种对生境中现有资源的利用能力和对现有环境的适应能力,在本研究中,红松生态位宽度和重要值在群系中最高,说明选定样地是以红松为主的红松阔叶混交林,红松、硕桦、紫椴为该群系的建群种,且在该群系中多个树种生态位宽度较大,在乔木层中占优势地位,群落整体结构处于动态变化之中。在前人的研究中发现,阔叶红松林中,红松、紫椴、色木槭、水曲柳、青楷槭具有较大的生态位宽度,但在不同群系中,相同树种的生态位宽度不同,明确具体群系中树种的生态位宽度极为重要[25-26]。优势树种在维持群落稳定性和生态功能等方面具有重要作用,保护和促进这些优势树种的生长和繁殖将有助于维持群落健康状态,在实际管理中,可开展优势树种自然更新培育,通过天然更新实现群落的复层林分结构。 生态位重叠用于反映群落种不同种群对环境资源利用能力的相似性及对空间位置占据程度,可以在一定程度上展现不同物种在群落中的相互作用和竞争关系[27]。本研究发现,该群系中红松与白桦、蒙古栎、水曲柳、臭冷杉、色木槭、暴马丁香等生态位重叠复杂,而与朝鲜槐、胡桃楸、辽椴、大青杨等种生态位重叠较小,不同树种生态位重叠有差异,呈网状结构,优势树种间的生态位分化程度较好。前人研究发现,随着群落的发育,生态位重叠度高的物种随着演替进行展现出竞争排斥或生态位分化,影响群落发育程度,可能会降低物种的生态位重叠度[28]。红松与朝鲜槐、胡桃楸、辽椴、大青杨等种群的生物学特性和对资源的要求上存在明显的差异,导致它们之间的生态位重叠较小,这与前人研究结论较为一致,即不同树种对资源利用存在差异、生态位重叠程度低的树种之间竞争较弱[29]。因此,本研究补充了阔叶红松混交林中红松+紫椴+硕桦群系的中优势树种中的生态位分析。 生态位和种间联结是衡量群落重要的结构指标,反映群落内不同物种间对有限资源的竞争和种间共存的情况,群落物种间的总体联结性在一定程度上与群落稳定性有关[30]。种间联结是由环境差异影响物种分布引起的现象,指群落中不同物种在空间分布上的相互关联性并描述由于群落生境差异造成的不同物种间生态习性上的差异及种间关系,是种间相互作用的结果[31]。本研究使用方差比率法分析红松+紫椴+硕桦群系中优势树种之间的关系,结果表明,该群系的优势树种整体上呈现显著正联结,优势树种之间可能相互促进生长,共同占据资源,形成一个相对稳定的共生群落;结合Pearson相关检验结果发现,在所有种对中,种间联结性较高的种对占总种对数高于种间联结性较低的种对占总种对数,且正联结的种对间表现出互补、互利的关系,而负联结的种对间则表现出竞争与排斥的关系,这些发现对于深入理解群落内物种之间的相互作用模式和竞争关系,以及群落结构和功能的形成具有重要意义。 红松、硕桦、紫椴作为建群种在群系中起着重要的作用,它们在一定程度上决定了群落的发展方向,种间联结分析有助于揭示它们在一定时期内物种的生境需求和种间的相互关系,从而推断群落的发展方向。根据本研究的结果,优势树种的正联结的种对大于负联结的种对,且大多数种对的联结性均不显著,说明群落乔木层中,优势树种之间正相互作用占主导,但这种正相互作用的强度较低,这可能意味着在当前的群落结构中,优势树种之间的竞争相对较弱,它们可能更多地是通过合作和互利来维持群落的稳定。在一些研究中发现,某些群落演替初期的物种间关联程度较低或呈现负关联,随着演替的发生,物种对资源的利用需求不断增加,竞争与互作使群落结构更加稳定,现有共存的物种之间的关系总体趋向于正联结[9,32-33]。优势树种之间的正联结关系表明它们相互促进生长、共同利用资源,在实际的林地管理中,可以采取促进树种共生的策略,如合理的间伐和树种搭配,以促进树种间资源的共享和相互支持。在本研究中虽然正联结在群落中占主导,但是部分种对之间仍然存在竞争关系,适度的调控竞争,避免某些树种过度竞争而导致资源浪费,有助于维持群落的平衡。综合以上研究结果,丰林国家级自然保护区阔叶红松混交林中的红松+硕桦+紫椴群系的群落结构相对稳定、向正向演替发展,这些研究结果对于更深入地了解阔叶红松林群落特征,以及群落中优势树种的相互作用和共存机制具有重要意义。 本文以丰林国家级自然保护区的红松+紫椴+硕桦群系为对象,深入分析了阔叶红松林内优势树种的生态位特征和种间关系。研究表明,红松、硕桦和紫椴等建群树种在群系中占据重要地位,具有广泛的资源利用能力和适应性。此外,群系中优势树种整体上呈现显著正联结,正联结的种对数大于负联结的种对数,优势树种间正相互作用占优势,正相互作用的强度较低,群系群落结构相对稳定、向正向演替发展。这些发现对于深入理解阔叶红松林内优势树种的生态适应和相互作用机制具有重要意义,为阔叶红松混交林的生态保护和可持续经营管理提供了有益的参考。未来的研究可以进一步探索不同生境条件下的优势树种生态位的变化,以及它们对群落稳定性的影响,从而更加全面地揭示阔叶红松林的生态机制和演替过程。3 结果与分析
3.1 优势树种的生态位宽度
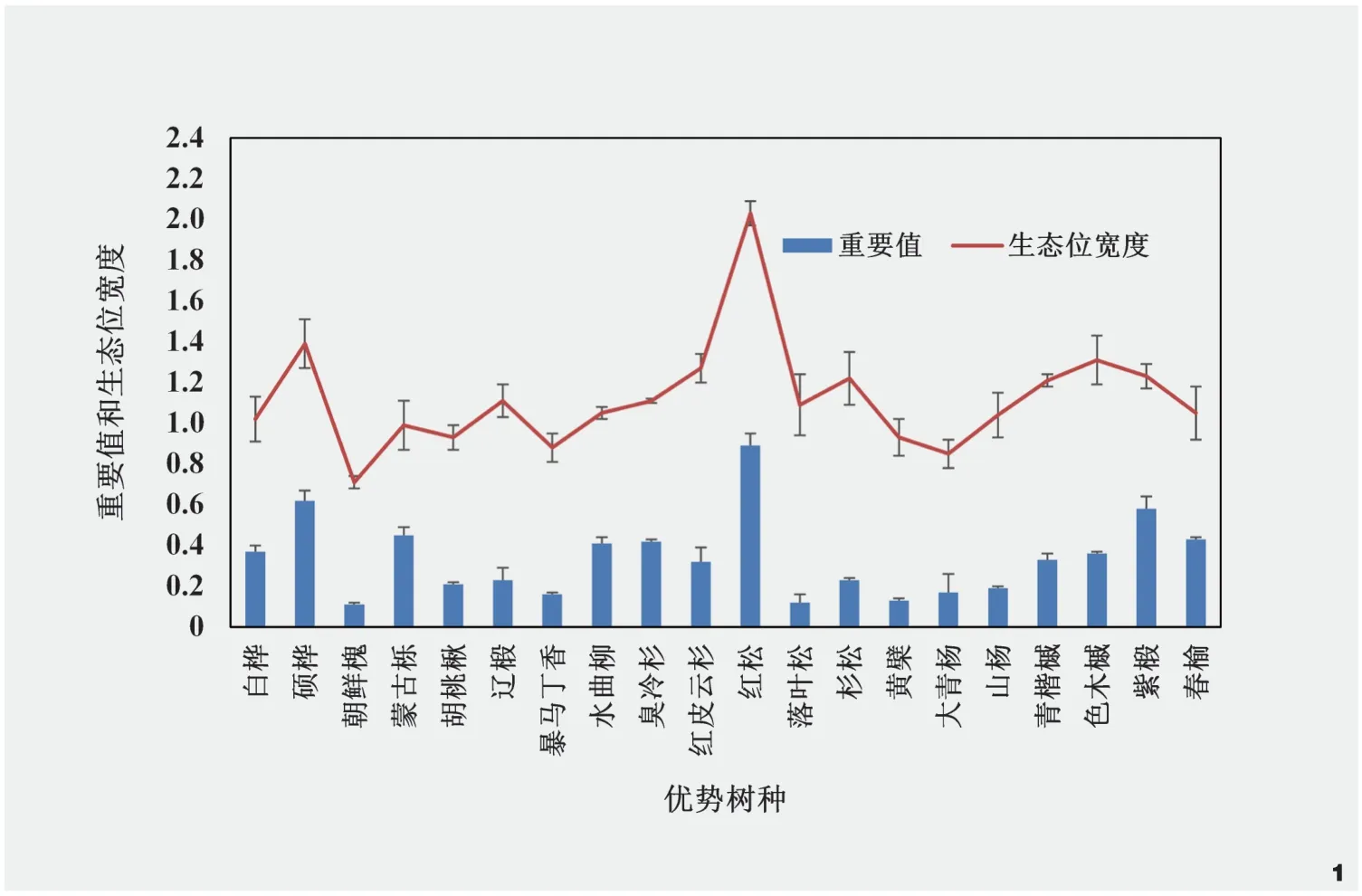
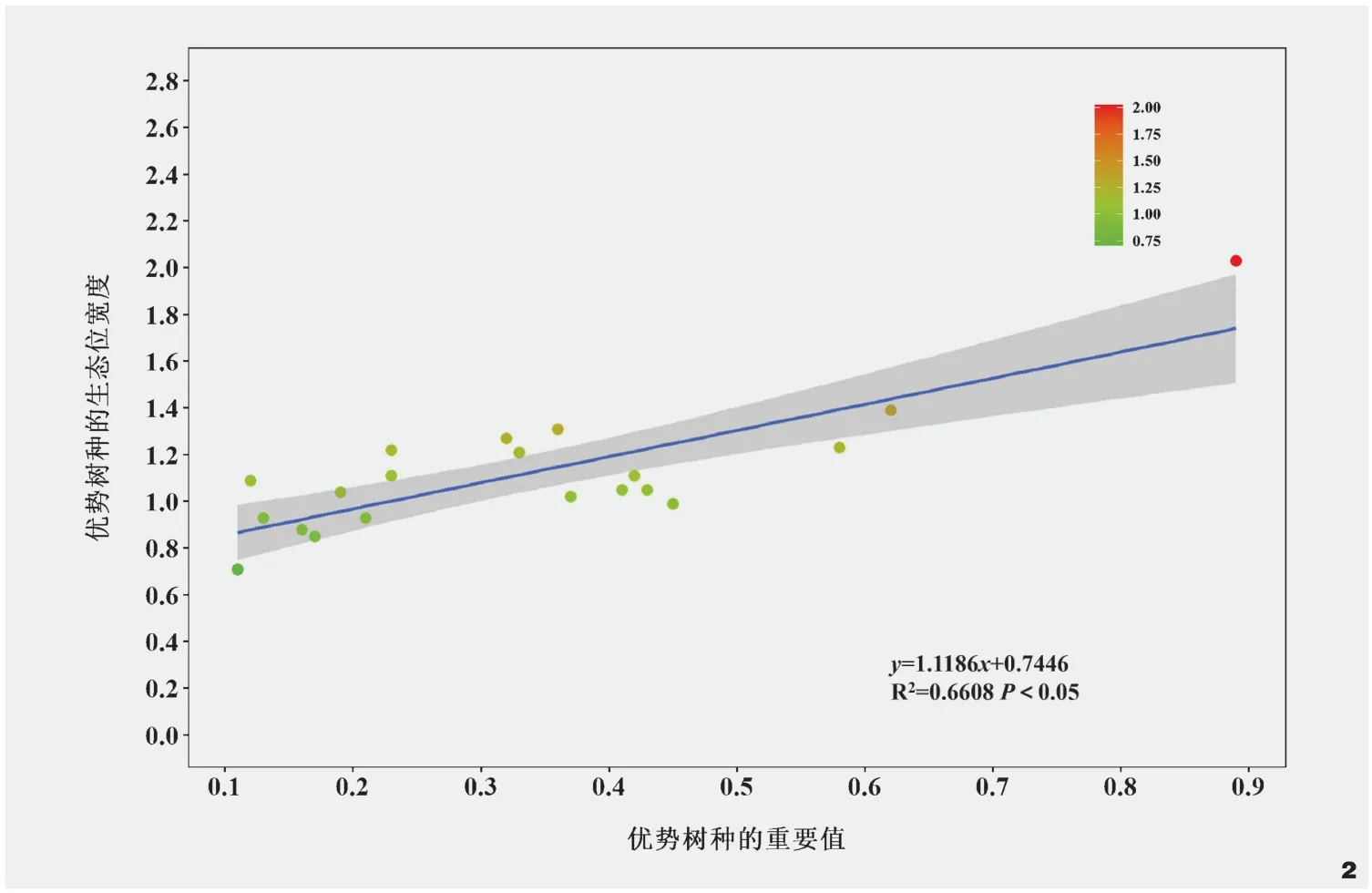
3.2 优势树种的生态位重叠

3.3 优势树种的种间联结
4 讨论
4.1 优势树种生态位特征分析
4.2 优势树种种间联结差异分析
5 结论
