独脚金内酯对甘蓝型油菜种子的引发作用及其机制研究
2023-10-17邓凤玲朝洪波李春生顾建伟
祝 四,邓凤玲,赵 光,朝洪波,李春生,顾建伟
(1.湖北工程学院生命科学技术学院,湖北 孝感 432000;2.郑州大学农业科技学院,郑州 450001)
油菜(Brassica napusL.)种子活力是一个复杂的数量性状,涉及许多生化和分子生物学过程,种子活力降低会导致出苗率低和幼苗衰弱,最终导致大田产量下降[1,2]。由此可见,高抗逆性、高生长优势和高生产潜力的高活力种子对农业生产十分重要[3,4]。
在中国许多地区,干旱胁迫是导致甘蓝型油菜种子萌发和幼苗生长受抑制和延迟的主要原因。干旱胁迫严重影响作物的种子萌发和正常发育。种子引发被认为是促进植物发芽、提高发芽率和均匀度、增强抗逆性的重要技术,并已被证实可以提高许多受胁迫作物的发芽率和出苗率[5,6]。
植物生长调节因子是植物生长发育的重要成分,且可以作为引发剂改善各种作物在种子萌发和幼苗生长方面的表现[7]。独脚金内酯(Strigolactone,SLs)作为一类新的植物生长调节因子,是一种衍生自类胡罗素的萜类化合物,能够影响豌豆、拟南芥和水稻等植物的发育过程[8-11]。目前,已经鉴定或合成了多种天然独脚金内酯或独脚金内酯类似物,其中GR24 的活性最高并已被广泛使用[12]。独脚金内酯与其他植物生长调节因子间存在复杂的相互作用并可以提高植物对胁迫的适应性,如独脚金内酯在拟南芥中可以作为正调控子参与胁迫响应并提高植物对干旱和盐胁迫的耐受性[1,13,14]。
基于此,本研究通过分析独脚金内酯处理油菜种子后其相关生理和生化指标及脱落酸(Abscisic acid,ABA)和赤霉素(Gibberellin,GA)信号途径相关基因的变化,探究了独脚金内酯在甘蓝型油菜种子引发中的作用及相关机理,以期为开发新的种子引发剂、提高油菜种子活力、扩大油菜在干旱地区种植面积提供技术支持和参考依据。
1 材料与方法
1.1 供试材料
油菜品种:阳光2009,中国农业科学院油料作物研究所提供。
试剂:独脚金内酯(GR24),购自上海源叶生物科技有限公司;聚乙二醇6000(PEG-6000),购自国药集团化学试剂有限公司;生化指标分析试剂盒均购自南京建成生物工程研究所。
1.2 试验设计
1.2.1 种子引发处理 称取0.3 mg GR24(相对分子质量为298.29),加入0.1 mL 无水乙醇溶液助溶后用超纯水定容至100 mL,配置成10.00 μmol/L 的GR24溶液,4 ℃保存用于后续试验。
油菜种子表面消毒后,使用不同浓度的GR24[T1:10.00 μmol/L;T2:1.00 μmol/L;T3:0.10 μmol/L;T4:0.01 μmol/L]在20 ℃避光浸泡24 h,同时设置去离子水浸泡处理(H2O)和未处理种子对照(CK),然后20 ℃避光干燥24 h,处理后的种子4 ℃保存备用或直接用于后续试验。
1.2.2 干旱处理 引发处理的种子和未处理的种子在10%PEG(PEG-6000)溶液浸透的滤纸上于23 ℃黑暗条件下萌发以模拟干旱胁迫。该试验设置4 个处理,分别为去离子水条件下萌发的未处理种子(CK)、10.00 μmol/L GR24 处理的种子(T1)、干旱胁迫下萌发的未处理种子(干旱胁迫对照,CK+10%PEG)、干旱胁迫下10.00 μmol/L GR24 处理的种子(T1+10%PEG)。每个处理设置3 个重复。萌发后于0、12、24、36、48 h 取样,于液氮中迅速冷冻后放于-80 ℃超低温冰箱中保存。
1.3 测定内容及方法
1.3.1 萌发分析 每个培养皿中放置40粒种子,每12 h记录一次每个培养皿的萌发情况,直至72 h萌发结束。计算种子萌发指数(GI)和萌发率(GR),计算式如下。
式中,N1至N6为每次统计时记录新萌发种子的数量。
茎长和根长均为生长到第6 天的测定值。
1.3.2 理化指标测定 丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸(Thiobarbituric acid,TBA)法检测[15]。脯氨酸(Proline,Pro)含量采用酸性茚三酮显色法测定[16]。按照试剂盒说明书依次添加检测试剂测定过氧化氢酶(Catalyse,CAT)和超氧化物歧化酶(Superoxide dismutase,SOD)的活性。
1.3.3 基因表达分析 使用高纯度总RNA 快速提取试剂盒(天根生化技术北京有限公司)提取油菜种子和幼苗(萌发0、24、48 h)的总RNA,并使用Rever-Tra Ace qPCR RT Master Mix with gDNA Remover 试剂 盒(TOYOBO,Japan)反 转 录 为cDNA,在ABI Prism® 7900HT 型荧光定量PCR 仪上进行实时荧光定量PCR 分析。以BnPPR基因为内参基因(与AT1G61780 同源)[17],采用2-ΔΔCT法计算基因的相对表达量,用于试验的引物如表1 所示。
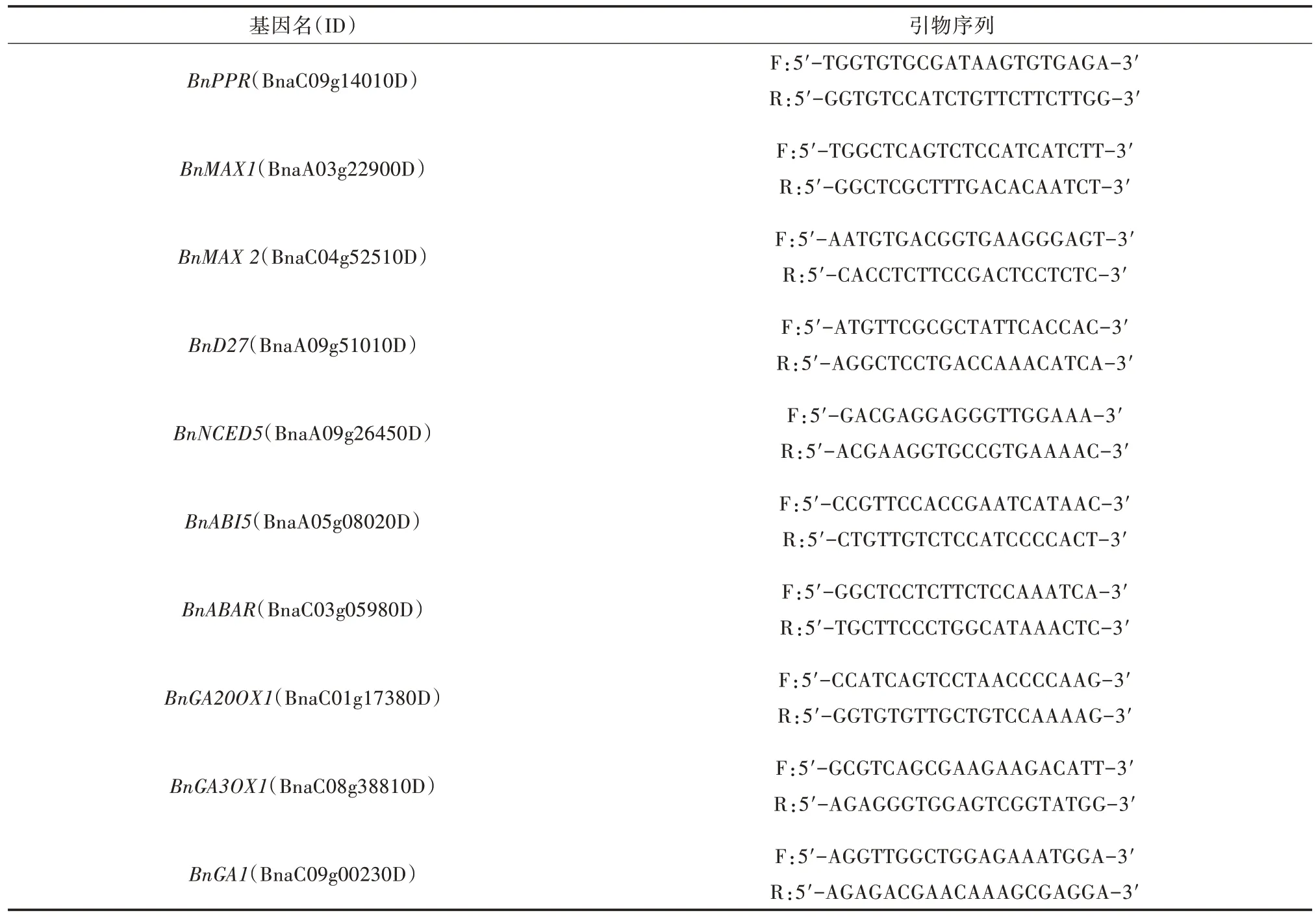
表1 荧光定量PCR 分析所用引物
1.4 统计分析
采用SPSS 22 软件对试验数据进行单因素方差分析。
2 结果与分析
2.1 GR24 处理对甘蓝型油菜种子萌发和幼苗生长的影响
用4 种不同浓度的GR24 对油菜种子进行引发处理后,在23 ℃黑暗条件下对种子萌发指数和萌发率进行统计分析。与对照相比,10.00 μmol/L 的GR24 引发处理油菜种子的萌发指数最高,其次是1.00 μmol/L 的GR24 引发处理,最低为0.01 μmol/L的GR24 引发处理(图1a)。同时,10.00 μmol/L 的GR24 引发处理种子萌发后60 h 萌发率即达100%,高于其他处理(图1b)。因此,本试验选择10.00 μmol/L 作为GR24 处理的最佳浓度。
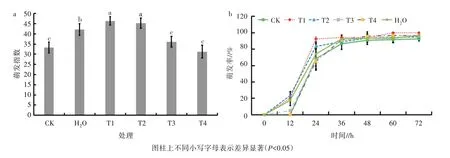
图1 不同浓度GR24 引发处理的油菜种子的萌发指数和萌发率
2.2 模拟干旱条件下GR24 引发处理对油菜种子萌发和幼苗生长的影响
以10%的PEG-6000 溶液模拟干旱,研究GR24引发处理对干旱条件下油菜种子活力的影响。GR24 处理的种子在正常(黑暗,23 ℃,无胁迫)和干旱条件(黑暗,23 ℃,10% PEG)下均比相应对照萌发更快,并且在萌发后24、36、48 h 的根长更长(图2)。在正常条件下,GR24 引发处理的种子萌发指数比CK 增加了13.08,萌发率比CK 增加了7.5 个百分点;在干旱条件下,GR24 引发处理的种子萌发指数比干旱胁迫对照增加了3.81,萌发率比干旱胁迫对照增加了5.00 个百分点(表2),表明GR24 引发处理促进了种子萌发。在正常条件下,GR24 引发处理的幼苗茎长和根长分别比CK 增加了1.48、1.60 cm;在干旱胁迫下,GR24 处理幼苗的茎长和根长分别比干旱胁迫对照增加了0.85、1.67 cm(表2)。

图2 10.00 μmol/L GR24 引发处理对油菜种子萌发的影响
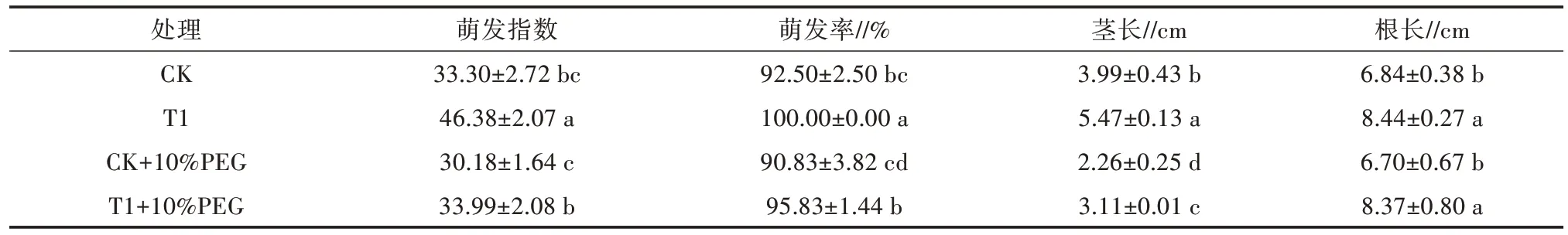
表2 10.00 μmol/L 的GR24 引发处理对油菜萌发指数、萌发率、根长和茎长的影响
2.3 模拟干旱条件下GR24 引发处理对MDA、Pro含量及主要抗氧化酶活性的影响
为了进一步分析GR24 引发处理对油菜种子萌发和幼苗生长的影响,检测了种子萌发和幼苗生长过程中的MDA、Pro 含量以及主要抗氧化酶活性的变化。结果表明,油菜种子/幼苗的MDA 含量随萌发而不断变化,CK 种子萌发后的MDA 含量均显著高于未萌发种子(图3a),说明在种子从干燥休眠状态向代谢活跃的萌发状态转变过程中脂质过氧化水平明显升高。干旱胁迫在种子萌发过程中显著加剧了脂质过氧化。在正常条件下,与CK 相比,萌发后36 h 和48 h 时,GR24 引发处理分别使种子/幼苗的MDA 的积累降低了40%和27%;干旱胁迫下,与干旱胁迫对照相比,萌发36 h 和48 h 时,GR24 处理种子中的MDA含量分别降低了55%和29%(图3a)。
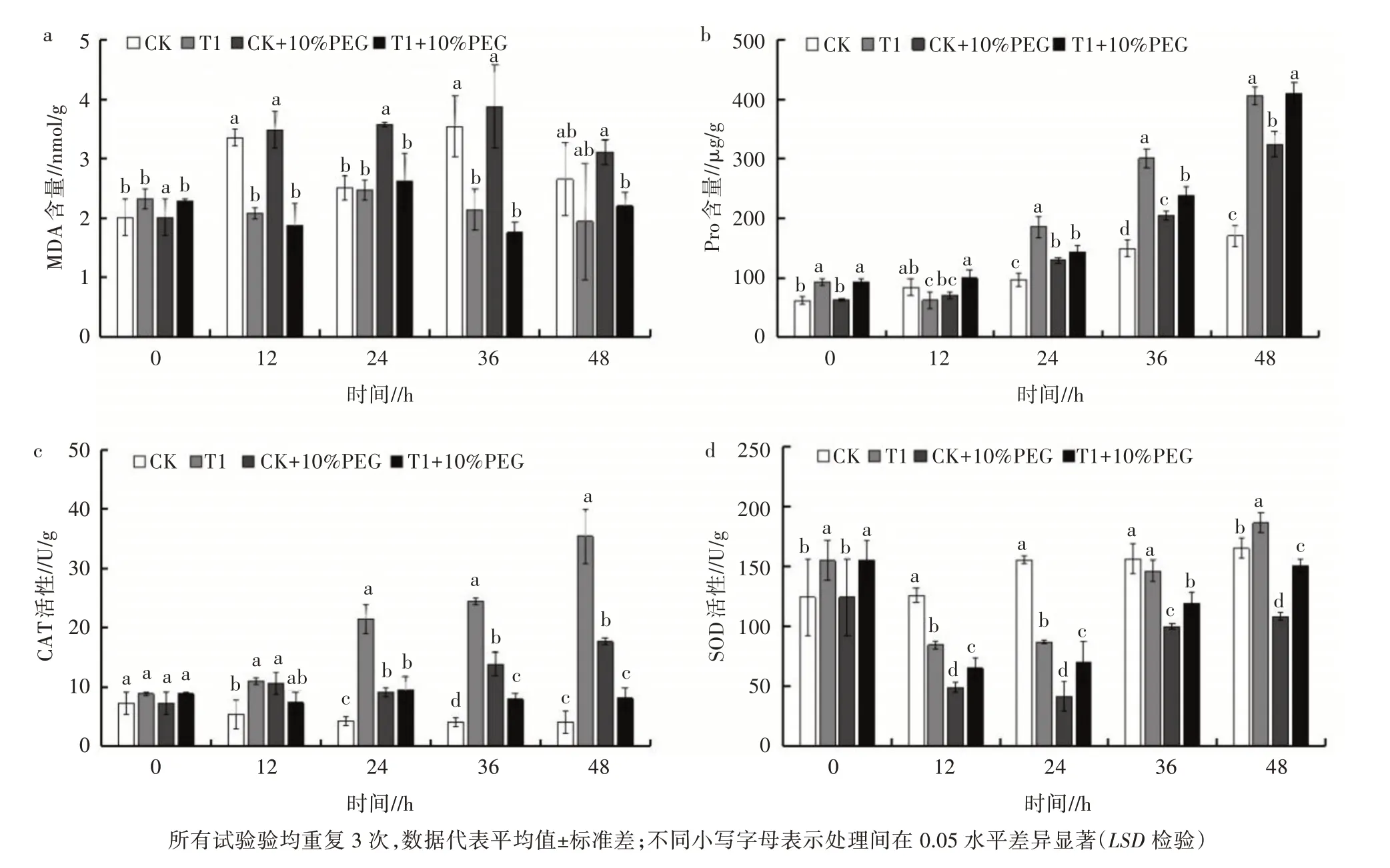
图3 油菜种子萌发过程中Pro、MDA 的含量及CAT、SOD 的活性
油菜种子/幼苗的Pro 含量在正常条件下随时间延长呈上升趋势,48 h 达最高水平(图3b)。除12 h外,GR24 引发处理增加了各萌发阶段种子/幼苗中的Pro含量。在干旱胁迫条件下,不同萌发时间的种子/幼苗中,GR24 引发处理均可提高Pro 含量,说明GR24 处理增强了种子萌发对干旱胁迫的适应性。
在正常条件下,CK 中油菜种子/幼苗的CAT 活性呈缓慢下降趋势,而经GR24 处理后,CAT 活性总体呈上升趋势,在萌发12、24、36、48 h 时均显著高于CK。在干旱胁迫下,GR24 处理和未处理种子/幼苗的CAT 活性始终高于正常条件下的CK,干旱胁迫对照的油菜种子/幼苗CAT 活力随萌发时间延长而增加,并且在36 h 和48 h 时高于GR24 处理,但始终低于正常条件下生长的GR24 处理幼苗(图3c)。在正常条件下,CK 种子/幼苗中的SOD 活性随种子萌发而逐渐升高,GR24 引发处理显著增加了0、48 h 种子/幼苗中的SOD 活性,并且在48 h 处理的幼苗中也检测到更高的SOD 活性;在干旱胁迫下,处理和未处理的种子/幼苗中SOD 活性均呈先下降后上升的趋势,但GR24 处理种子/幼苗中SOD 的活力总是显著高于未处理的种子/幼苗(图3d)。
2.4 与萌发相关的生长调节因子代谢和信号途径的主要基因表达模式
根据已有报道,筛选了SLs(BnMAX1、BnMAX2、BnD27)、ABA(BnABI5、BnNCED5、BnABAR)和GA(BnGA20OX1、BnGA3OX1、BnGA1)代谢和信号途径的几个主要基因。通过荧光定量PCR(qRT-PCR)方法,以未处理油菜种子作为对照,分析了GR24 引发处理对这些基因在正常条件下和干旱胁迫条件下表达的影响,结果如图4 所示。
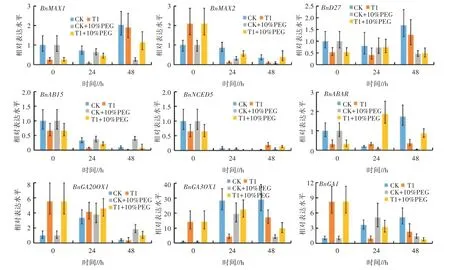
图4 植物生长调节因子相关基因在萌发种子中的表达模式
独脚金内酯生物合成的核心途径由β-胡萝卜素异构酶(D27,AT1G03055)和2 种类胡萝卜素裂解双加氧酶(CCD7 和CCD8)组成,最终形成Carlactone,即所有已知独脚金内酯的前体。BnMAX1(AT2G26170)编码细胞色素P450 蛋白,可直接转化Carlactone。BnMAX2(AT2G42620)在SL 感知中发挥重要作用。未萌发种子中,GR24 处理高度抑制了BnD27和BnMAX1的表达,但诱导BnMAX2的表达。与正常条件相比,BnD27的表达在萌发48 h 时受到干旱胁迫明显抑制,但CK 与T1 处理间没有发现明显差异。在正常条件下,BnMAX1的表达在CK 和GR24 处理下均随种子萌发时间的延长呈先降低后升高的趋势,且该基因在GR24 处理中的表达始终低于CK;但在干旱条件下,BnMAX1的表达在干旱胁迫对照下呈下降趋势,在GR24 处理下呈升高趋势。在种子萌发过程中,各处理BnMAX2的表达随种子的萌发持续降低,但与正常条件下不同的是,在干旱条件下GR24 处理中BnMAX2的表达始终高于干旱胁迫对照。
BnNCED5(AT1G30100)编码ABA 生物合成中的关键调控酶。BnNCED5的表达在GR24 处理后也呈下降趋势。BnABI5(AT2G36270)参与种子成熟和萌发过程中ABA 的信号传导。与CK 相比,GR24 引发处理后BnABI5的相对表达水平在萌发过程中明显降低,干旱胁迫没有改变GR24 处理对该基因表达的影响。BnABAR(AT5G13630)调节从质体到细胞核的ABA 信号转导。在正常条件下,GR24 引发处理后,BnABAR的表达在0、48 h 时均比CK 有所降低,而在干旱条件下,GR24 引发处理在24 h 和48 h时提高了BnABAR的表达。
在未萌发种子中,GR24 引发高度诱导了3 个GA 代谢和信号通路相关基因,但在萌发过程中表现出不同的表达模式。BnGA1(AT4G02780)是催化GA生物合成途径中的第一个关键步骤。与BnGA20OX1(AT4G25420)和BnGA3OX1(AT1G15550)不同的是,在24 h 和48 h 时,GR24 处理中BnGA1的表达始终低于相应对照。BnGA20OX1和BnGA3OX1参与种子中GA 生物合成的后期步骤。在正常条件下,GR24 处理的BnGA20OX1在0 h 和24 h 表达水平分别为CK的5.54 倍和1.23 倍;在干旱胁迫条件下,GR24 处理的BnGA20OX1的表达水平在24 h 时为干旱胁迫对照的1.22 倍,而在48 h 时2 个处理的BnGA20OX1表达水平均呈急剧下降趋势,且GR24处理明显低于干旱胁迫对照。在正常条件下,GR24处理中BnGA3OX1的表达呈先降低后升高的趋势,且在24、48 h 时低于CK;在干旱胁迫下,GR24 处理的BnGA3OX1表达在0、24、48 h 时均高于干旱胁迫对照。
3 讨论与小结
3.1 SLs 引发处理改善种子萌发、幼苗生长和干旱胁迫耐受性
SLs 参与调节植物与丛枝菌根真菌的共生,并且在根部结构调控和非生物胁迫反应中也具有重要作用[18,19]。为验证SLs 在种子萌发中的作用,用不同浓度的GR24 对油菜种子进行引发处理。结果表明,油菜种子的萌发指数、萌发率及幼苗茎长和根长在正常和干旱胁迫条件下均显著高于相应对照,表明GR24 处理促进了种子萌发和幼苗生长,并可以提高其干旱胁迫耐受性。
3.2 SLs 引发处理提高了抗氧化系统的活力
MDA 是判断植物在胁迫下氧化损伤程度的指标[20]。本研究表明,GR24 处理显著降低了种子萌发过程中MDA 的积累,表明GR24 处理对膜胁迫损伤发挥了强大的积极预防作用,从而确保了萌发过程中膜系统的正常功能[15]。
脯氨酸是植物在逆境胁迫下产生的一种重要的渗透调节物质,在保护细胞膜结构和功能、维持细胞内外渗透压、调节体内酸碱平衡等方面具有重要作用[21]。在正常条件和干旱胁迫下,Pro 含量随着油菜种子萌发呈不断升高的趋势,除种子萌发12 h 外,对照的Pro 含量始终低于GR24 处理,说明GR24 引发能促进Pro 的积累,进一步调节氧化还原平衡以抵御外部不利条件。
在植物受到环境胁迫时,体内活性氧含量上升,因此植物必须要维持SOD 和CAT 在内的保护酶系统以清除活性氧的过多积累,保护植物正常生长[20,22]。本研究表明,在正常条件下,GR24 处理可以增加种子及幼苗中CAT 的积累量,但在干旱条件下,GR24 处理CAT 活性呈下降趋势,可能是由于GR24 引发处理显著提高了SOD 含量,改变了保护酶系统对环境胁迫的响应模式造成的。这表明GR24 引发处理可以调节种子萌发和幼苗生长过程中的抗氧化酶活性,提高油菜种子在干旱胁迫下的抗氧化能力,与前人的结果是一致的[23,24]。
3.3 GR24 引发处理通过调节不同途径相关基因的表达来改善油菜种子萌发和幼苗生长
已有研究证明SLs 代谢和信号相关基因在成熟的种子和幼苗中表达[19]。植物生长调节因子间存在广泛的相互作用,SLs 和ABA 在其他生理过程中的的相互作用已被证实[14,24,25]。ABA 和GA 在种子萌发调控中的重要作用也被大量文献报道[20]。因此,推测SLs 引发处理可以通过调节ABA、GA 代谢和信号通路来促进种子萌发和幼苗生长。为了验证这一假设,分析了几个与SLs、ABA、GA 代谢和信号通路相关的重要基因在萌发过程中的表达。
GR24 处理使未萌发种子中BnMAX1和BnD27表达显著降低,而使BnMAX2表达显著升高,表明GR24 处理可能会抑制种子内源SLs 的合成,但是提前启动了SLs 的信号转导途径。未处理种子中的BnMAX1和BnD27表达量在萌发早期较低,但在萌发48 h 时明显升高,而BnMAX2的表达一直呈下降趋势,类似的趋势在GR24 处理种子萌发过程中也被观察到,表明SLs在萌发早期(0~24 h)和进入幼苗生长阶段(48 h)可能具有不同的作用机制。拟南芥中的研究证明D27作用于BnMAX1上游从而通过SLs 控制植物的生长发育[26],而BnMAX2作用于Bn-MAX1下游参与SLs 信号转导,可以提高植物对干旱胁迫的耐受性[27]。本研究结果表明,干旱胁迫明显抑制了SLs 相关基因的表达,但是GR24 处理可以改善这种情况,GR24 在不同阶段可能是通过增强内源SLs 的合成或信号转导提高了萌发油菜种子和幼苗对干旱胁迫的耐受性。
ABA 和GA 是控制种子萌发的核心调节因子[28,29]。本研究结果表明,ABA 途径相关基因的表达受到GR24 处理的抑制,而GA 途径相关基因的表达受到GR24 处理的诱导。同时,在种子萌发阶段,GR24 处理增强了SLs 信号基因BnMAX2的表达。BnMAX2不仅是SLs 通路的重要组成部分,还受多种不同生长调节因子信号通路调控,参与植物生长发育的诸多方面,影响植物体内多种生长调节因子的代谢和信号转导[30-32]。BnMAX2已被证明在BnABI5的上游发挥作用,参与调控种子萌发和萌发后幼苗的早期发育。BnMAX2在ABA 信号通路中的作用可能随发育阶段的不同而变化,它可能在ABA 和渗透胁迫诱导的种子萌发和幼苗生长早期抑制中发挥负调控作用[33]。这些结果表明,GR24 处理负调控ABA 途径并正调控GA 途径,最终导致休眠解除,加速萌发和早期幼苗生长。
ABA 在介导植物对非生物胁迫的防御反应中起主要作用,但同时也与其他生长调节因子间存在复杂的相互作用[34]。本试验中,干旱胁迫在48 h 时诱导了GR24 处理油菜种子/幼苗BnNCED5和BnABAR的 表达。同 时,BnMAX1和BnMAX2在GR24 处理也表现出较高的表达。这些结果表明,SLs 引发处理可能通过与ABA 的协同作用提高油菜幼苗对干旱胁迫的耐受性。
3.4 小结
本研究表明,独脚金内酯引发处理对种子萌发和幼苗生长具有显著的促进作用。独脚金内酯引发处理的这种促生长作用可能是通过调节ABA 和GA代谢及信号通路相关基因的表达来实现的。这些结果证明了独脚金内酯作为种子活力的有效促进剂的应用潜力,但相关机制需要进一步研究,也并未排除独脚金内酯直接调节种子萌发的可能性。
