棉蚜抗药性现状及治理策略
2023-10-16石丹丹
石丹丹, 张 帅, 梁 沛*
(1. 中国农业大学昆虫学系,北京 100193;2. 全国农业技术推广服务中心,北京 100125)
棉蚜 (瓜蚜)AphisgossypiiGlover (半翅目:蚜科)是一种世界性的农业昆虫[1-2],可通过直接取食或传播植物病毒为害多种寄主植物,是锦葵科 (棉花、木槿等)、葫芦科 (西瓜、黄瓜等)和茄科 (茄子、辣椒等)等多种农作物及一些观赏植物的重要害虫[3-5]。化学杀虫剂是棉蚜综合防治中不可或缺的主要防治措施,但是棉蚜较强的繁殖能力和适应能力以及杀虫剂的不合理使用,致使棉蚜对多种杀虫剂产生了抗药性,严重威胁农作物安全生产,造成巨大的经济损失[6-7]。梁彦等[6]于2013年总结了1964年至2012年国内外棉蚜对杀虫剂的抗性演化历史,本文主要对近10年来棉蚜的抗药性现状、抗性机制和抗性治理策略进行综述,意在为棉蚜的可持续治理和科学施策提供借鉴。
1 棉蚜对杀虫剂抗性的发展和现状
有机磷类、氨基甲酸酯类杀虫剂具有广泛的杀虫活性,是棉蚜早期化学防控中首选的杀虫剂[8]。随后拟除虫菊酯类杀虫剂因其高效、低毒等特点得到了广泛应用,这也导致了棉蚜对这三大类杀虫剂抗性的快速发展[6]。20世纪90年代吡虫啉出现后,新烟碱类杀虫剂迅速成为防治蚜虫最有效的替代品[9]。但很快棉蚜对新烟碱类杀虫剂也产生了抗性[7]。近年来,新型杀虫剂如磺酰亚胺类的氟啶虫胺腈、吡啶酰胺类的氟啶虫酰胺以及生物源杀虫剂双丙环虫酯等也陆续应用于棉蚜的田间防治。虽然这些药剂刚开始都具有非常好的防治效果,但由于不合理使用,棉蚜同样对其产生了抗性 (表1)。
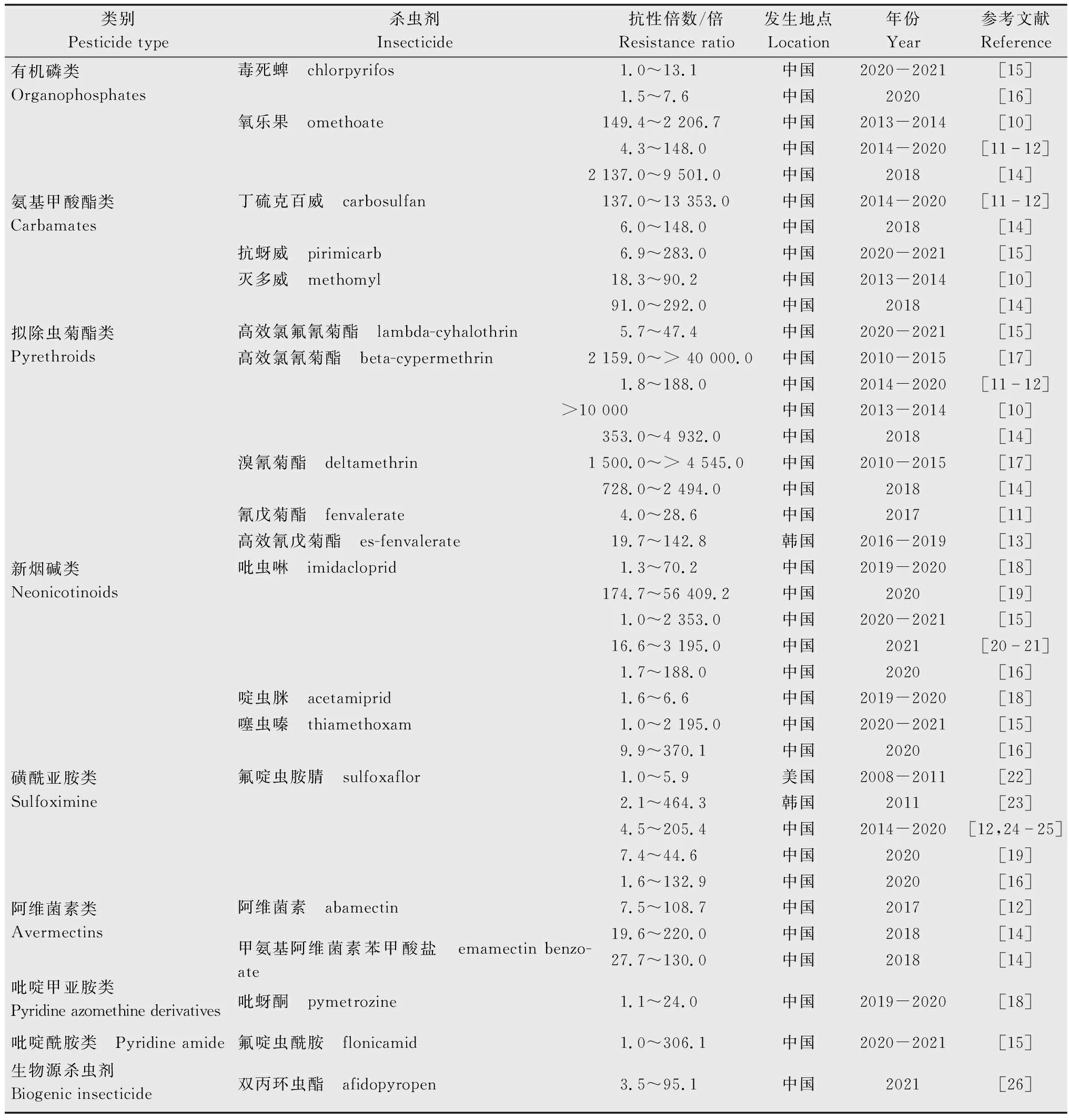
表1 棉蚜近10年的抗药性概况Table 1 Overview of insecticide resistance in Aphis gossypii in the past ten years
1.1 棉蚜对有机磷和氨基甲酸酯类杀虫剂的抗性
2013年-2014年,张帅等[10]发现山东滨州、河南西华两地棉蚜种群对有机磷类杀虫剂 (氧乐果)具有高水平抗性;河北邱县、山东滨州、河南西华三个地区棉蚜种群对灭多威处于中等水平抗性 (抗性倍数在18.3~90.2倍之间)。自2014起连续7年对河北保定、沧州和邯郸3个地区进行监测发现,棉蚜对氧乐果具有中等至高水平抗性,对丁硫克百威为高水平抗性,最高达10 000多倍[11-12]。2016年-2019年,Nam等[13]发现韩国温室辣椒上的棉蚜种群对乙酰甲胺磷处于敏感水平。2018年,新疆哈密等6个地区棉蚜对氧乐果抗性达到2 137~9 501倍,对灭多威和丁硫克百威的抗性分别处于中等至高水平抗性和低至高水平抗性[14]。2020年-2021年,河南、湖北和湖南3个省份棉蚜 (瓜蚜)对毒死蜱处于敏感至低水平抗性;对抗蚜威处于低至中等水平抗性[15]。2020年,崔丽等[16]发现山东、河北和河南7个棉蚜种群对毒死蜱处于敏感至低水平抗性。总体来看,除了新疆棉区外,其他地区棉蚜对有机磷类杀虫剂抗性呈现下降趋势,处于敏感至低水平抗性,对氨基甲酸酯类杀虫剂产生了不同水平的抗性,不同棉区之间抗性水平差异性较大,部分棉区棉蚜对丁硫克百威维持在高水平抗性。
1.2 棉蚜对拟除虫菊酯类杀虫剂的抗性
2010年-2015年,我国新疆、湖北、安徽、山西、河南、江苏、山东7个省份11个棉蚜种群对高效氯氰菊酯和溴氰菊酯均表现出极高的抗性,抗性倍数达1 500~40 000倍[17]。2013年-2014年,张帅等[10]发现河北邱县、山东滨州、河南西华3个地区棉蚜种群对高效氯氰菊酯的抗性均已达10 000倍以上。2014年-2020年,河北保定、沧州和邯郸3个地区棉蚜对高效氯氰菊酯同样处于高水平抗性[11-12]。2016年-2019年,Nam等[13]发现韩国温室辣椒上的棉蚜对高效氰戊菊酯处于中等至高等水平抗性 (19.7~142.8倍)。2017年,河北保定、沧州和邯郸3个地区棉蚜对氰戊菊酯处于低至中等水平抗性[11]。2018年,新疆哈密等6个地区棉蚜对溴氰菊酯和高效氯氰菊酯分别达到了728~2 494倍和353~4 942倍的高水平抗性[14]。2020年-2021年检测结果表明,河南、湖北和湖南3个省份棉蚜(瓜蚜)对高效氯氟氰菊酯低至中等水平抗性[15]。2020年,崔丽等[16]的检测发现,山东和河南地区棉蚜对高效氯氟氰菊酯处于低至中等水平抗性。总体来看,棉蚜对高效氯氰菊酯、溴氰菊酯等拟除虫菊酯类杀虫剂均处于高水平抗性,对于高效氯氟氰菊酯抗性水平处于上升阶段。
1.3 棉蚜对新烟碱类杀虫剂的抗性
马康生等[7]对2004年至2019年间棉蚜对新烟碱类杀虫剂的抗性相关文献进行了较为全面的梳理,发现国内外棉蚜种群对该类药剂均已产生了不同水平的抗性,本文不再赘述。最新检测表明,贵州省4个地区辣椒上的棉蚜对啶虫脒处于敏感至低水平抗性,对吡虫啉处于敏感至中等水平抗性[18]。2020年,李仁等[19]对河北、山东、山西和新疆共16个地区的棉蚜种群的检测结果表明,这些棉蚜种群对吡虫啉均处于高水平抗性。2020年-2021年的检测结果发现河南、湖北和湖南3个省份棉蚜 (瓜蚜)对吡虫啉和噻虫嗪处于敏感至高等水平抗性,最高分别达到2 353倍和2 195倍[15]。2020年,崔丽等[16]发现山东、河北和河南棉花上采集的6个棉蚜田间种群均对吡虫啉和噻虫嗪处于中等至高水平抗性;1个采自花椒上的棉蚜种群对吡虫啉敏感,对噻虫嗪具有9.9倍的低水平抗性。2021年,对新疆、河北、河南、山西及山东5省共19个棉蚜田间种群的检测结果表明,除河南南阳市和新疆图木舒克市两个种群仍然保持敏感外,其他17个种群均对吡虫啉产生了抗性 (16.6~3 195倍)[20-21]。总体来看,棉蚜对新烟碱类杀虫剂抗性水平差异较大,大部分棉区的棉蚜对吡虫啉仍处于高水平抗性。
1.4 棉蚜对磺酰亚胺类杀虫剂的抗性
该类杀虫剂主要是氟啶虫胺腈。2008年-2011年,美国中南部 (密西西比州、路易斯安那州、阿肯色州、田纳西州和德克萨斯州)的棉蚜对氟啶虫胺腈仍然保持敏感性 (1.0~5.9倍)[22],但韩国扶余、骊州和清州3个地区的棉蚜已对其产生了中等至高水平抗性,最高达464倍[23]。氟啶虫胺腈在我国最早于2013年登记用于棉蚜防治,但2014-2020年,河北棉区棉蚜对氟啶虫胺腈的抗性不断上升,从4.5倍上升至205倍[12, 24-25]。李仁等[19]2020年的检测结果表明,河北、山东、山西和新疆4省(区)16个棉蚜种群对氟啶虫胺腈维持在低至中等水平抗性。崔丽等[16]2020年的最新研究表明,山东、河北和河南棉区棉蚜对氟啶虫胺腈处于中等至高水平抗性,其中山东泰安种群抗性水平最高,抗性倍数达132.9倍。总体来看,氟啶虫胺腈对棉蚜的防治效果下降,部分棉区表现为高水平抗性。
1.5 棉蚜对其他杀虫剂的抗性
除上述几大类外,目前用于棉蚜防治的杀虫剂还有阿维菌素、吡蚜酮、双丙环虫酯和溴虫氟苯双酰胺等。2017年安静杰等[12]的结果表明,河北保定、邯郸和沧州棉区棉蚜种群对阿维菌素产生了中等至高水平抗性 (7.5~108.7倍)。2018年,新疆哈密等6个地区棉蚜种群对阿维菌素和甲维盐的抗性同样处于中等至高水平抗性,最高达220倍[14]。
虽然双丙环虫酯2019年才在我国登记用于棉蚜的防治,但本实验室2019年-2021年连续3年的监测结果表明,河北、山东、山西和新疆等重点棉区棉蚜种群对双丙环虫酯的抗性已经呈现明显上升趋势[26]。
2019年-2020年,韦治艳等[18]对采自贵州省4个地区辣椒上的棉蚜检测发现,其对吡蚜酮处于敏感至中等水平抗性。本实验室最近的检测结果表明,新疆、河北、河南、山西及山东5省(区)共19个棉蚜田间种群对新型杀虫剂环丙氟虫胺和溴虫氟苯双酰胺均保持敏感[20-21]。
2 棉蚜对杀虫剂抗性的分子机制
目前,关于棉蚜对杀虫剂抗性机制的报道主要涉及代谢抗性、靶标抗性和肠道共生菌参与的抗性3个方面。
2.1 代谢抗性
代谢抗性是指由于解毒代谢能力增强导致的抗药性,是害虫对杀虫剂产生抗性的重要机制之一。杀虫剂有效成分到达靶标的量只占进入体内总药量的很少部分,大部分都被解毒酶,如细胞色素P450多功能氧化酶、羧酸酯酶、ABC转运蛋白等降解或结合后贮存于脂肪体中或排出体外。这些酶的代谢能力增强能够使到达靶标的杀虫剂大幅度减少,从而提高害虫对杀虫剂的耐受能力,产生抗药性[27-28]。
昆虫细胞色素P450多功能氧化酶 (cytochrome P450 monooxygenases,P450s)是昆虫体内主要的解毒代谢酶,具有多种重要的生理功能,包括催化和代谢多种不同结构的内源和外源底物,在棉蚜对杀虫剂的抗性中发挥着关键作用[29]。有研究表明,P450基因表达量升高导致的酶活性增强是棉蚜对吡虫啉、噻虫嗪等新烟碱类杀虫剂产生抗性的主要原因[7]。此外,Peng等[30]和Pan等[31]研究表明P450基因CYP6A2和CYP380C6的过表达与棉蚜对螺虫乙酯的抗性密切相关。Ma等[32]在抗氟啶虫胺腈棉蚜品系中鉴定到多个过表达的P450基因,进一步利用RNA干扰技术证明CYP6CY13和CYP6CY19介导了棉蚜对氟啶虫胺腈的抗性。Zeng等[33]通过构建转基因果蝇品系证明了CYP380C6、CYP4CJ1和CYP6CY21的过表达参与了棉蚜对溴氰虫酰胺的抗性;CYP380C6、CYP6CY7和CYP6CY21的过表达还导致溴氰虫酰胺棉蚜抗性品系对顺式氯氰菊酯产生了交互抗性。
羧酸酯酶 (carboxylesterases,CarEs)是参与杀虫剂代谢的重要水解酶,很多研究表明其活性增强与棉蚜对有机磷类、氨基甲酸酯类和拟除虫菊酯类等传统杀虫剂的抗性密切相关,例如羧酸酯酶基因的过表达介导了棉蚜对马拉硫磷、氧乐果和溴氰菊酯的抗性[34-36]。但是羧酸酯酶在棉蚜对新烟碱类杀虫剂抗性中的作用尚不明确[7]。崔丽等[37]发现羧酸酯酶活力的增强与棉蚜对吡虫啉的抗性相关,但缺乏进一步的功能验证。
谷胱甘肽S-转移酶 (glutathioneS-transferases,GSTs)在昆虫对化合物的初级代谢和次级代谢中均发挥着重要作用[38],但有关GSTs与棉蚜抗药性的报道较少,有限的研究表明GSTs活力的升高与棉蚜对吡虫啉的抗性相关[37],但有待进一步验证。
UDP-葡糖基转移酶 (UDP-glycosyltransferase,UGT)是重要的Ⅱ相解毒酶。有研究表明UGT基因的过表达参与了棉蚜对吡虫啉和噻虫嗪的抗性[39-41]。Zeng等[33]通过构建转基因果蝇品系证明了UGT341A4、UGT344B4、UGT344M2的上调表达参与了棉蚜对溴氰虫酰胺的抗性,其中UGT341A4和UGT344M2 的过表达参与溴氰虫酰胺和顺式氯氰菊酯的交互抗性[33]。Ma等[42]利用RNA干扰技术证明UGT344B4、UGT344C5和UGT344A11的过表达调控棉蚜对氟啶虫胺腈的抗性。Pan等[43]发现UGT介导的糖氧化作用与棉蚜对螺虫乙酯的抗性有关,尤其是UGT344M2的过表达有助于棉蚜对螺虫乙酯的解毒代谢。
ABC转运蛋白 (ATP-binding cassette transporters,ABCs)在杀虫剂抗药性中的主要功能是参与杀虫剂及其代谢产物的跨膜运输,从而将其从细胞中转运出去[44]。有研究结果表明ABC转运蛋白除了在棉蚜对吡虫啉和噻虫嗪的抗性中发挥作用之外,包括ABCB4、ABCB5、ABCF2、MRP11和MRP12在内的多种ABC转运蛋白基因的过表达与棉蚜对螺虫乙酯的抗性有关[45-46],而ABCB5、ABCG4、ABCG7、ABCG16、ABCG17、ABCG26和MRP12的过表达则参与了棉蚜对溴氰虫酰胺的抗性[47]。
此外,有研究表明化学感觉蛋白 (chemosensory proteins,CSPs)也参与了棉蚜对杀虫剂的抗性。CSPs也是节肢动物中的一类转运蛋白,在昆虫感受外界化学信号等多个生理功能发挥着重要作用[48]。Li等[49]首次证实在黑腹果蝇中过表达棉蚜AgosCSP5基因可显著增强果蝇对氧乐果、吡虫啉和氯氰菊酯的抗性。Xu等[50]发现AgoCSP1、AgoCSP4和AgoCSP5在棉蚜抗溴氰虫酰胺品系中组成性过表达,分别抑制这3个CSP基因的表达可显著提高抗性棉蚜对溴氰虫酰胺的敏感性,证明其参与了棉蚜对溴氰虫酰胺的抗性。Xu等[51]的研究表明,化学感觉蛋白基因CSP1和CSP4的过表达参与了棉蚜对螺虫乙酯的抗性。上述研究只是证明这些CSP基因参与了棉蚜对不同杀虫剂的抗性,但对其参与抗性的分子机制均缺乏了解,有待进一步研究。
2.2 靶标抗性
靶标抗性是指由于杀虫剂分子靶标的结构或表达水平变化导致的抗药性。氨基甲酸酯类和有机磷类杀虫剂的作用靶标是昆虫乙酰胆碱酯酶 (acetylcholionesterase,AChE),董双林等[52]综述了棉蚜AChE对杀虫剂不敏感的分子机制,表明AChE基因的两个突变 (S431F和A302S)是棉蚜对多种氨基甲酸酯类和有机磷类杀虫剂不敏感的主要原因。马康生等[7]综述了棉蚜抗新烟碱类杀虫剂抗性分子机制的研究进展,认为烟碱型乙酰胆碱受体β1 亚基 (nicotinic acetylcholine receptorβ1 subunit,nAChRβ1)的氨基酸突变以及受体基因表达量下调是导致棉蚜产生抗性的重要原因。比如,已证实烟碱型乙酰胆碱受体β1 亚基的L80S、R81T、V62I 和K264E突变与棉蚜对吡虫啉的抗性有关[21, 23]。
电压门控钠离子通道 (voltage-gated sodium channels,VGSC)的基因突变是昆虫对拟除虫菊酯类杀虫剂产生抗性的主要原因,特别是其跨膜结构域ⅡS4-S6上L1014F导致击倒抗性 (knockdown resistance,kdr)及其与M918L突变共同导致的超级击倒抗性 (super-kdr) 在家蝇Muscadomestica等多种昆虫中都已经被证实[53]。棉蚜VGSC的这两个突变同样导致其对拟除虫菊酯类杀虫剂的抗性。 Williamson等[54]和Chen等[17]分别发现棉蚜钠离子通道L1014F和 M918L突变使得棉蚜田间种群对高效氯氰菊酯和溴氰菊酯具有高水平抗性。此外,Munkhbayar等[55]发现新疆棉蚜种群除了M918L突变外,还发现了一个新的氨基酸取代 (M918V)。
值得注意的是,棉蚜田间抗性种群的不同靶标蛋白上往往同时存在多个抗性相关的氨基酸突变,如新疆棉蚜种群中同时存在VGSC上的 M918L/V和nAChR上的 R81T两个位点的突变[55],山东棉蚜种群中同时存在nAChR上的R81T、K264E和VGSC上的L1014F突变[56],我国中部地区棉蚜种群同时存在nAChR的V62I和R81T、AChE的A302S和S431F及VGSC的M918L等5个突变,从而导致同一棉蚜种群多对种类型的杀虫剂产生了抗性[15]。 韩国温室中为害辣椒的棉蚜种群中同样同时存在nAChR (R81T)、VGSC (M918L)和AChE (S431F)3个靶标基因的突变[13]。
此外,靶标受体基因表达量的改变也可能在抗性发展中发挥作用。乙酰辅酶A羧化酶 (acetyl-CoA carboxylase,ACC)是一种生物素依赖酶,主要催化脂肪酸的生物合成[57]。季酮酸类杀虫剂螺虫乙酯作为ACC的抑制剂,通过抑制ACC进而干扰总脂质的生物合成从而杀死害虫[58]。有研究表明,ACC的过量表达导致棉蚜对螺虫乙酯的敏感性降低[59]。
2.3 肠道共生细菌介导的抗性
昆虫肠道共生菌可以促进宿主对植物毒素和杀虫剂的抵抗力[60]。张云骅等[61]综述了昆虫肠道共生菌与宿主营养和解毒代谢的关系,昆虫共生菌可以直接代谢有毒物质或者间接调控宿主的解毒代谢。Zhang 等[62]发现敏感和螺虫乙酯抗性品系棉蚜肠道共生菌菌群的种类和丰度均存在显著差异,且用抗生素 (氨苄青霉素和四环素)处理可增强螺虫乙酯对抗性棉蚜的毒力,说明某些肠道共生菌可能参与了棉蚜对螺虫乙酯的抗性。Guo等[63]发现取食葫芦科植物的两种不同体色 (绿色和黄色)的棉蚜 (瓜蚜)对杀虫剂抗性的差异与Buchnera属共生菌的丰度增加有关。本实验室最新研究表明,肠道中鞘氨醇单胞菌Sphingomonas的丰度增加显著增强了棉蚜对吡虫啉的抗性;进一步研究发现鞘氨醇单胞菌可通过羟基化和硝基还原直接代谢吡虫啉 (代谢效率高达 56%),从而介导了棉蚜对吡虫啉的抗性[64]。
3 棉蚜抗性治理策略
随着棉蚜对杀虫剂抗性的不断发展,对其抗性进行有效治理仍然面临巨大挑战。要有效治理棉蚜的抗药性,至少应该做到以下几个方面。
3.1 强化综合防治策略
对包括棉蚜在内的几乎所有害虫可持续控制的首选策略应该是害虫综合治理 (integrated pest management,IPM)。而在IPM中,首先应该考虑的是如何充分利用农业防治、生物防治和物理防治等非化学防治措施。化学防治虽然也是IPM策略的主要内容之一,但它是IPM的“压舱石”,只有其他措施无法有效压低害虫种群数量时才考虑使用化学杀虫剂。因为任何化学杀虫剂,只要长期使用就会导致害虫对其产生抗药性,这几乎是不可避免的。而且化学杀虫剂的大面积使用还会对田间天敌种群 (自然的或人工释放的)造成影响,显著降低其自然控害能力,加快棉蚜种群数量增长。到目前为止,还没有一种化学杀虫剂对所有天敌都安全。因此,在同样能有效控制害虫种群数量的情况下,尽可能地少用或者不用化学防治,以降低害虫产生抗药性的风险及由此带来的一系列问题。
3.2 加强棉蚜田间种群抗性监测
明确棉蚜的抗药性现状是科学选药和精准用药的关键,而加强对田间种群的抗药性监测则是了解棉蚜田间种群对杀虫剂抗性现状的最直接、有效的方法。抗药性监测的目的有二:一是了解当年不同地区棉蚜对常用的主要杀虫剂的抗性水平及抗性基因频率,为下一年科学选药、合理用药提供依据;二是了解同一地区不同年份间棉蚜抗药性变化动态及发展趋势,为制订中、长期抗性治理策略提供参考。对于刚刚登记使用的新型杀虫剂尤其要做好抗药性监测,特别是抗性基因频率的监测,在抗性基因频率明显上升但在种群水平还未表现出明显抗性时及时制订预防性抗性治理策略,避免抗性的进一步发展。
3.3 科学合理使用杀虫剂
首先要根据对本地棉蚜抗药性监测结果合理选药。对于棉蚜尚未产生明显抗药性或低水平抗性的杀虫剂可以继续使用;对于棉蚜已经产生中等水平抗性的杀虫剂应该严格限制使用,即每个生长季只使用1次,避免抗性进一步发展;对于已经产生高水平抗性、在推荐剂量下防治效果明显下降的杀虫剂应停止使用。对于可以使用的杀虫剂,最好选择与作用靶标不同且无交互抗性的杀虫剂混合使用 (可以购买商品化的混配制剂,也可以在农技人员指导下进行桶混),以达到既有效控制棉蚜,又延缓其抗性发展的目的。对于刚上市的新型杀虫剂尤其要注意与现有杀虫剂的合理混用,不仅可以有效延缓棉蚜对其抗药性的产生和发展,还能在显著降低防治成本 (新药一般都比较贵)的情况下达到理想的防治效果,可谓一举三得。
杀虫剂的轮换使用是有效治理棉蚜抗药性的另一重要措施。高希武先生指出,杀虫剂的轮用的主要依据是害虫抗药性的产生是在杀虫剂存在条件下“瞬间进化”的结果,当停止使用杀虫剂后,种群中的抗性个体在“瞬间进化”条件 (杀虫剂的选择压力)消失后,因其生物适应性比敏感个体低而逐渐被淘汰,导致其在种群中所占比例不断下降,敏感个体的比例不断增加,使得整个种群对杀虫剂的敏感性得以恢复[65]。因此,合理轮换使用作用机制不同且无交互抗性的杀虫剂,也可有效延缓棉蚜抗药性的发展。
3.4 合理使用增效剂
合理使用增效剂是延缓害虫抗药性发展的又一有效措施。增效剂可通过增加药剂对昆虫体壁的渗透性、抑制昆虫解毒酶活性及增强药剂的内吸性等多种途径显著提高杀虫剂的活性并延缓抗药性发展。目前商品化的增效剂产品比较多,在棉蚜防治中,加入商品化的增效助剂可以减少噻虫嗪的使用量达30%[66]。需要注意的是,同一增效剂对不同杀虫剂的增效作用可能存在较大差异,有的甚至会起到反作用,因此在使用前一定要通过田间试验筛选出真正有效的增效剂。
3.5 加强多学科合作、创新防控策略
加强多学科合作,不断创新害虫防控策略为有效治理棉蚜抗药性发展提供有力的技术支撑。例如,利用纳米载体同时搭载低剂量的噻虫嗪、靶向突触蛋白的双链RNA (syn-dsRNA)可实现棉蚜的高致死率[67];在棉花中表达棉蚜的P450基因的双链RNAdsAgCYP6CY3-P1不仅显著延缓了棉蚜的生长发育,降低了其种群密度,还增加了其对吡虫啉和啶虫脒的敏感性[68]。
4 结语
棉蚜的抗药性不是一朝一夕形成的,因此对棉蚜抗药性的治理也不可能一蹴而就。只有真正执行IPM策略,优先采用非化学防治措施;加强棉蚜抗药性监测,并根据监测结果及时调整防控策略;加强科普宣传,指导种植户科学选药、合理用药,并不断研发和应用新的靶向基因的绿色防控新技术,才可能实现棉蚜的可持续治理,保障农业安全生产。
