原发性脑干损伤致死大鼠的脑干组织代谢组学分析
2023-10-15苏秦陈倩玲吴伟斌向青青杨成梁乔东访李志刚
苏秦,陈倩玲,吴伟斌,向青青,杨成梁,乔东访,李志刚
1.广州市刑事科学技术研究所 法医病理学公安部重点实验室,广东 广州 510442;2.中山大学中山医学院法医系,广东 广州 510080;3.南方医科大学法医学院,广东 广州 510515
脑干损伤是指对中脑、脑桥或延髓造成的损伤,是颅脑损伤中最危重的类型之一,起病急,病情进展快,预后差。脑干网状结构是维持意识的重要结构,因此,脑干损伤后常常发生意识障碍甚至死亡。原发性脑干损伤(primary brain stem injury,PBSI)在所有记录的颅脑损伤中约占2%,死亡率却高达70%[1]。PBSI包括脑干横断、挫伤、弥漫性轴索损伤和水肿[2]。在法医实际工作中,常遇到头面部遭受暴力后迅速死亡的案例,尸体检验(包括心传导系统检查)时,通常找不到或仅在显微镜下发现脑干有轻微、散在的小灶性出血,这给暴力致脑干损伤死亡的诊断带来困难[3-4]。此外,在PBSI 致死过程中,原发病灶和后续脑外伤并存,相互影响,对死因鉴定造成干扰,直接影响案件的定性,给刑事办案带来困难[5]。PBSI 的死因鉴定尚未建立明确、客观的诊断依据,目前主要借助于形态学方法,结合外伤史,在排除其他死亡原因的前提下得出鉴定意见[6]。因此,PBSI 的死因鉴别诊断仍是法医病理鉴定实践中的重点和难点。
随着社会法律意识与证据意识的提高,法医病理学者必须从新的切入角度来进行PBSI 的死因鉴别诊断。代谢组学分析被运用于死因鉴别和死亡时间鉴定中,并取得了较好的效果[7-9]。本研究拟对比PBSI与其他死因,如非脑干脑损伤致死或剪头处死的大鼠尸体脑干代谢组的变化规律,寻找代谢标志物,提出可用于诊断脑干损伤的生理病理学依据。
1 材料与方法
1.1 PBSI 大鼠模型的构建与评价
将72 只7 周龄、体质量约250 g 的雄性SD 大鼠(广东至远生物医药科技有限公司)随机分为PBSI组、非脑干脑损伤组和对照组(剪头处死),每组24 只。
PBSI 组大鼠于麻醉诱导箱中以3%异氟烷诱导麻醉,至角膜反射消失后取出,2%异氟烷维持麻醉。待大鼠头顶部备皮后进行碘附消毒,采用改良的Marmarou 模型进行动物建模(图1A)[10],新模型舍弃了缓冲垫,以张力极小的锡箔纸保持鼠头与身体水平,保证打击后头部绕矢状面旋转90°做角加速运动,更符合脑干损伤(创伤性轴索损伤)的损伤机制。具体步骤:沿中线切开头皮,暴露骨膜,将不锈钢垫片(直径1 cm、厚2 mm)用组织胶水固定于顶骨矢状缝中间(图1B),随后撤离麻醉剂,待大鼠稍微清醒后将其俯卧固定于海绵垫上,将质量为250 g 的不锈钢圆柱体自2 m 高度自由下落,撞击大鼠颅顶部不锈钢垫片使其头部产生快速过屈运动,且圆柱体打击后不产生反弹。大鼠遭到打击后迅速进入昏迷状态,相继于1 h内死亡。对于非脑干脑损伤组大鼠,除了将不锈钢垫片固定于额骨中间(图1C)外,其他操作同PBSI 组,经处理后的大鼠也相继于1 h 内死亡。对照组大鼠在麻醉后仅进行备皮消毒和切开头皮的处理,并于1 h 后剪头处死,剪断位置为颈椎处。
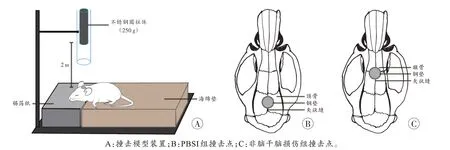
图1 大鼠模型的构建Fig.1 Construction of rat models
将3 组大鼠尸体全部放置在室温条件下(温度25~27 ℃,湿度50%~55%),并于死后6 h 从每组随机取8 只大鼠提取脑干组织。将剩余48 只大鼠尸体转移至0~4 ℃环境中,分别于死后1 d 和3 d 提取脑干组织,每组每个时间点8 只。
为评价构建的大鼠模型,取出大鼠全脑,沿正中矢状面对半切开后,取一半脑干组织置于4%多聚甲醛溶液中固定24 h,之后以切面为包埋面进行常规梯度乙醇脱水、二甲苯透明、浸蜡、包埋,并将蜡块连续切片(5 µm),分别进行常规HE 染色和甘氨酸银染色。
本研究涉及动物的实验均通过广东至远生物医药科技有限公司动物伦理学委员会的审查(审批文号:IAEC-2022022301)。
1.2 样本收集与处理
将切取的另一半脑干组织分装于无菌袋(4 cm×4 cm)中,经液氮速冻后转移至-80 ℃冰箱中保存。每份样本取120 mg组织进行液氮研磨,加入120 µL 50%甲醇溶液后充分混匀,提取样品中的代谢物。混匀物常温静置10 min 后转移至-20 ℃条件下过夜以沉淀样品中的蛋白质。4 000×g离心20 min,取上清液代谢物提取液至96 孔板用于上机检测,其中每个样品等量取出10 µL 稀释液进行混合,形成质量控制样品,用于检测检验方法的稳定性。
1.3 LC-MS 检测
用UltiMate 3000 超高效液相色谱仪(美国Thermo Fisher Scientific公司)分离色谱。采用ACQUITY UPLC BEH C18色谱柱(100 mm×2.1 mm,1.8 µm;英国Waters公司)反相分离,柱温35 ℃,流速0.4 mL/min;流动相A 为水溶液(含0.1%甲酸),流动相B 为乙腈(含0.1%甲酸)。梯度洗脱条件设置为:0~0.5 min 5%B,0.5~7.0 min 5%B~100%B,7.0~8.0 min 100%B,8.0~8.1 min 100%B~5%B,8.1~10 min 5%B。上样量为4 µL。
1.4 数据前处理
使用MSConvert 软件(美国ProteoWizard 公司)将下机的原始数据转换成可读数据mzXML 文件,再利用R 4.1.2软件(https://www.r-project.org/)和XCMS 3.4.1 包对代谢物离子峰进行提取和质量控制。通过R 软件和CAMERA 3.4.1 包以及Compound Discoverer 3.1.0 软件(美国Thermo Fisher Scientific 公司)分别对提取到的物质进行加和离子分析和物质注释(HMDB数据库,https://hmdb.ca),将二级质谱信息与代谢物标准品本地数据库(杭州联川生物技术股份有限公司)进行匹配。经K-最近邻(K-nearest neighbor,KNN)算法补充缺失值,概率商归一化(probabilistic quotient normalization,PQN)算法对样品数据归一化处理,利用质量控制样品进行质量控制-鲁棒样条校正(quality control-robust spline correction,QC-RSC),去掉不稳定代谢物。
1.5 统计分析
采用R 软件MetaX 1.4.16 包进行偏最小二乘-判别分析法(partial least square-discriminant analysis,PLS-DA),计算R2(表示模型解释能力)和Q2(表示模型预测能力),若均高于0.5,表明模型的解释率和预测率均较好[11]。同时利用MetaX 1.4.16 包以变量投影重要性(variable important in projection,VIP)评分≥1.0作为筛选条件筛选组间变化显著的代谢标志物[12],同时进行交叉验证,并绘制置换检验图。通过R 软件VennDiagram 1.7.1包绘制韦恩图。利用R软件random-Forest 4.7-1 包进行随机森林建模,首先采用Bootstrap方法对数据集进行抽样,每次抽取每组总体样本的75%作为训练集对样本进行分类,另外25%的数据(称为袋外数据)作为测试集用于估计随机森林的分类误差和计算变量的重要性。随机森林建模时将树的个数和每棵树包含的特征个数分别设置为500 和3。随后利用R 软件randomForest 4.7-1 包建立的模型对测试集进行预测。利用R 软件pROC 1.18.0 包进行受试者操作特征(receiver operator characteristic,ROC)曲线绘制。
2 结果
2.1 PBSI 模型评估
HE 染色结果显示:对照组大鼠额叶、脑室和脑干组织形态结构正常,未见明显病理改变;非脑干脑损伤组大鼠可见全脑蛛网膜下腔出血,实质内神经元及血管周围间隙扩大,侧脑室和第三脑室出血,额叶实质内散在点片状出血;PSBI 组大鼠可见程度不一的蛛网膜下腔出血,实质内神经元及血管周围间隙增宽,侧脑室、第三脑室和第四脑室出血,延髓实质内见多处灶状出血(图2)。甘氨酸银染色结果显示:对照组和非脑干脑损伤组大鼠脑干组织轴索排列整齐,走行正常,而PSBI 组大鼠脑干轴索排列紊乱,弥漫性断裂、肿胀、扭曲呈蚯蚓状,间隙增宽(图3)。上述结果表明,PBSI 模型构建成功。
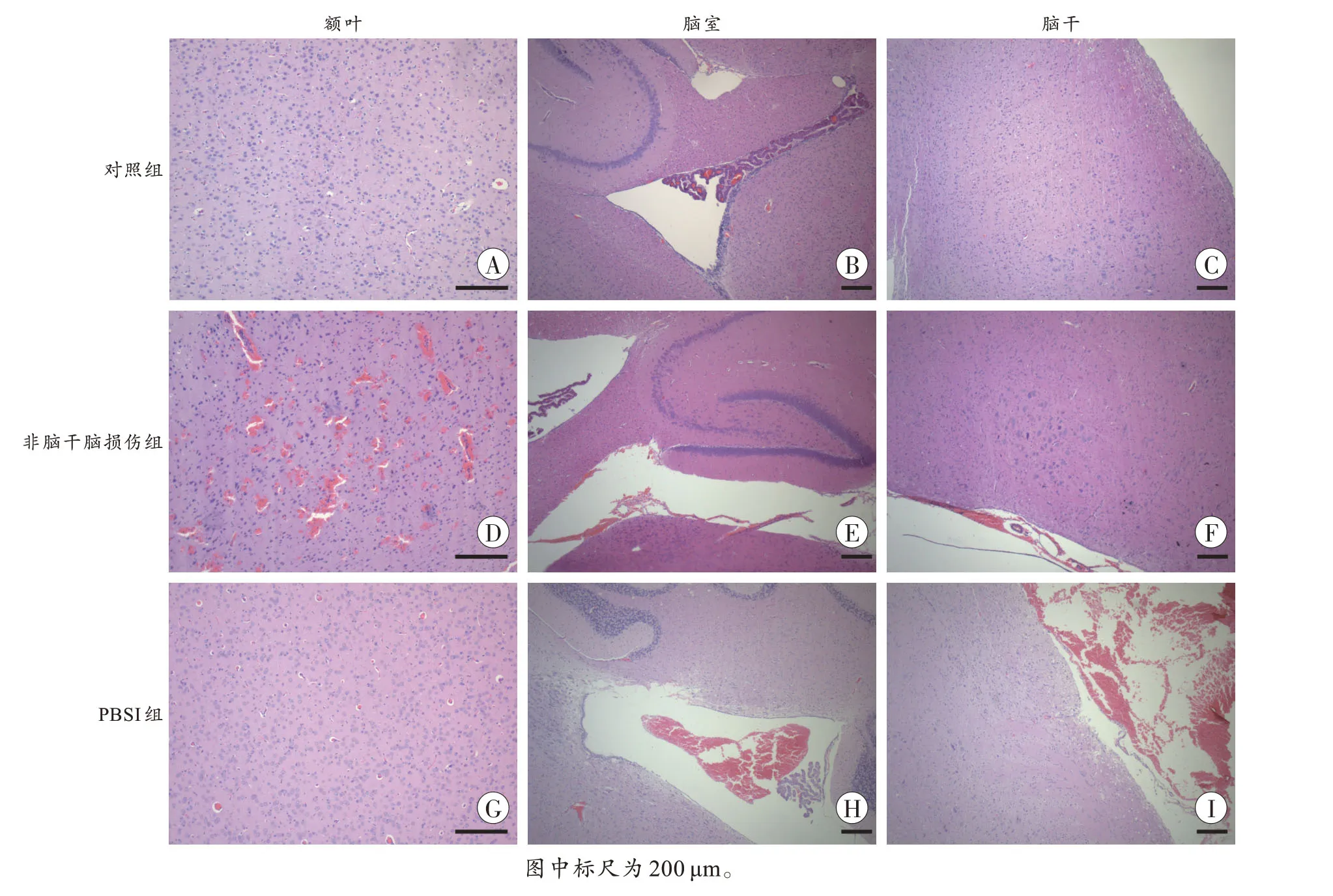
图2 不同死因大鼠脑组织的病理学改变(HE 染色)Fig.2 Pathological changes of rat brain tissues with different causes of death(HE staining)

图3 不同死因大鼠脑干组织的病理学改变(甘氨酸银染色)Fig.3 Pathological changes of rat brain stem tissues with different causes of death(Silver staining)
2.2 不同死因大鼠脑干组织的代谢物鉴定
3 组大鼠共检测到1 597 种代谢物,包括有机酸及其衍生物(29.34%),脂质和类脂分子(25.55%),杂环化合物(13.08%),苯类化合物(9.17%),有机氧化物(8.19%),苯丙酸类和聚酮化合物(4.77%),核苷(酸)及其类似物(3.55%),有机氮化物(2.81%)及其他(3.55%)。
2.3 基于PLS-DA 的差异代谢物分析
PLS-DA 被用于确定区分3 个死因模型中脑干的差异代谢物,利用1 597种代谢物中能被注释到HMDB数据库的818 种代谢标志物进行分析。死后6 h,3 种死因大鼠的脑干组织PLS-DA 结果显示,R2和Q2分别为0.94 和0.81,死后1 d 大鼠脑干组织的R2和Q2分别为0.92 和0.65,死后3 d 大鼠脑干组织的R2和Q2分别为0.92 和0.69(图4),且同一死因的样本紧密聚合在一起,表明同一方式处理的样本重复性良好。在3 个时间点的样本中,PBSI 组和对照组均能明显分离,而PBSI组和非脑干脑损伤组仅在死后6 h能有效分离,而在死后1 d和3 d存在部分重叠。对模型的质量进行交叉验证,置换检验结果显示Q2均<0,表明模型不存在过拟合的情况,基于此获得的差异代谢物准确(图5)。
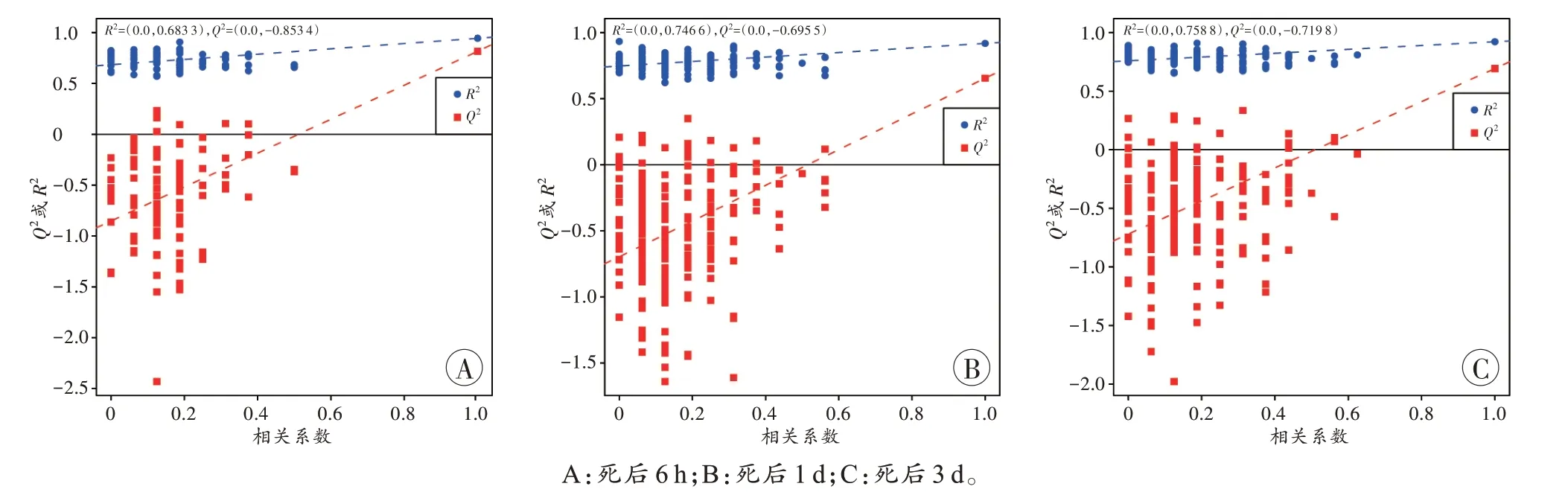
图5 不同死因大鼠死后不同时间点脑干组织OPLS-DA 的置换检验(200 次)结果Fig.5 Permutation test results(200 times)of OPLS-DA of rat brain stem tissues with different causes of death at different time points after death
同时通过计算VIP 衡量各代谢物表达模式对各组样本分类判别的影响强度和解释能力,从而辅助代谢标志物的筛选。结果显示,死后6 h、1 d 和3 d 大鼠脑干组织的代谢标志物分别为285、281 和301 种。通过韦恩图分析发现,死后3 个时间点大鼠脑干组织共有的代谢标志物为86 种(图6)。
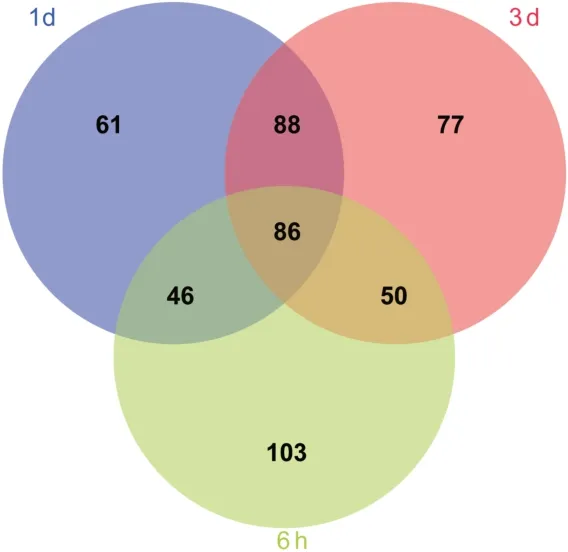
图6 死后6 h、1 d 和3 d 大鼠脑干组织代谢标志物的韦恩图Fig.6 Venn diagram of metabolic markers in rat brain stem tissues at 6 h,1 d and 3 d after death
2.4 基于随机森林的差异代谢物分析
为了从死后3个时间点大鼠脑干组织共有的86种代谢标志物中找到最具有组别区分效应的代谢物,将72 个样本的86 种代谢标志物的丰度信息构建数据集,从各组各时间点大鼠样本中随机抽取6 个样本构成训练集(共54个样本),其余样本构成测试集(共18个样本),进行随机森林模型构建和预测。通过对测试集数据进行预测,发现其准确率为83.3%(图7A,表1)。其中:对照组均预测正确;非脑干脑损伤组存在2 个预测错误,其中1 个被预测成对照组,另1 个被预测成PBSI 组;PBSI 组中有1 个被错误预测成非脑干脑损伤组。
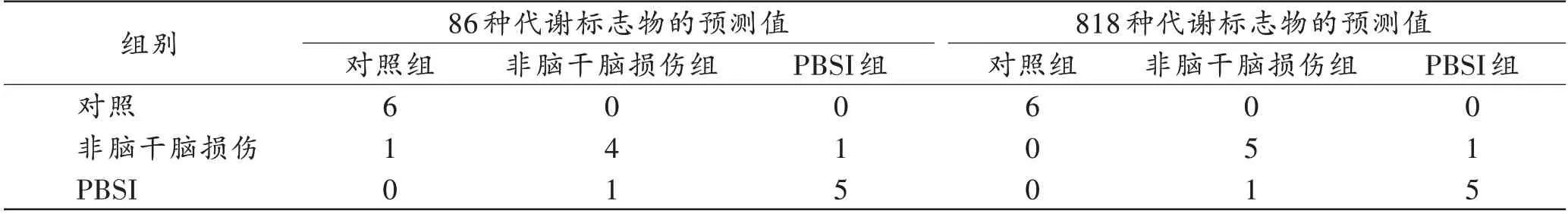
表1 86 种代谢标志物和818 种代谢标志物进行随机森林建模的预测结果Tab.1 Prediction results of random forest modeling using 86 metabolic markers and 818 metabolic markers(个)

图7 86 种代谢标志物的随机森林模型构建结果Fig.7 Construction results of random forest model for 86 metabolic markers
对随机森林中变量的重要性进行计算,结果(图7B)显示,占据前10 的分别为HMDB0038126[京尼平苷酸(genipinic acid,GA)]、HMDB0037386{(S)-薄荷酮-8-硫代乙酸酯[(S)-menthone 8-thioacetate]}、HMDB0059773[S-3-氧代癸酰半胱胺(S-3-oxodecanoyl cysteamine)]、HMDB0013272[N-月桂酰甘氨酸(N-lauroylglycine)]、HMDB0029365[阿魏酰酪胺(moupinamide)]、HMDB0040551[L-α-氨基-1H-吡咯-1-己 酸(L-alpha-amino-1H-pyrrole-1-hexanoic acid)]、HMDB0029427[L-次甘氨酸A(L-hypoglycin A)]、HMDB0005199{(R)-去甲猪毛菜碱[(R)-salsolinol]}、HMDB0013645[N,N-二甲基鞘氨醇(N,Ndimethylsphingosine)]和HMDB0001008[胆绿素(biliverdin)]。其中,与对照组和非脑干脑损伤组相比,在PBSI 组显著上调的代谢标志物分别为HMDB0038126(京尼平苷酸)、HMDB0013272(N-月桂酰甘氨酸)、HMDB0005199[(R)-去甲猪毛菜碱]和HMDB0013645(N,N-二甲基鞘氨醇),见图8。
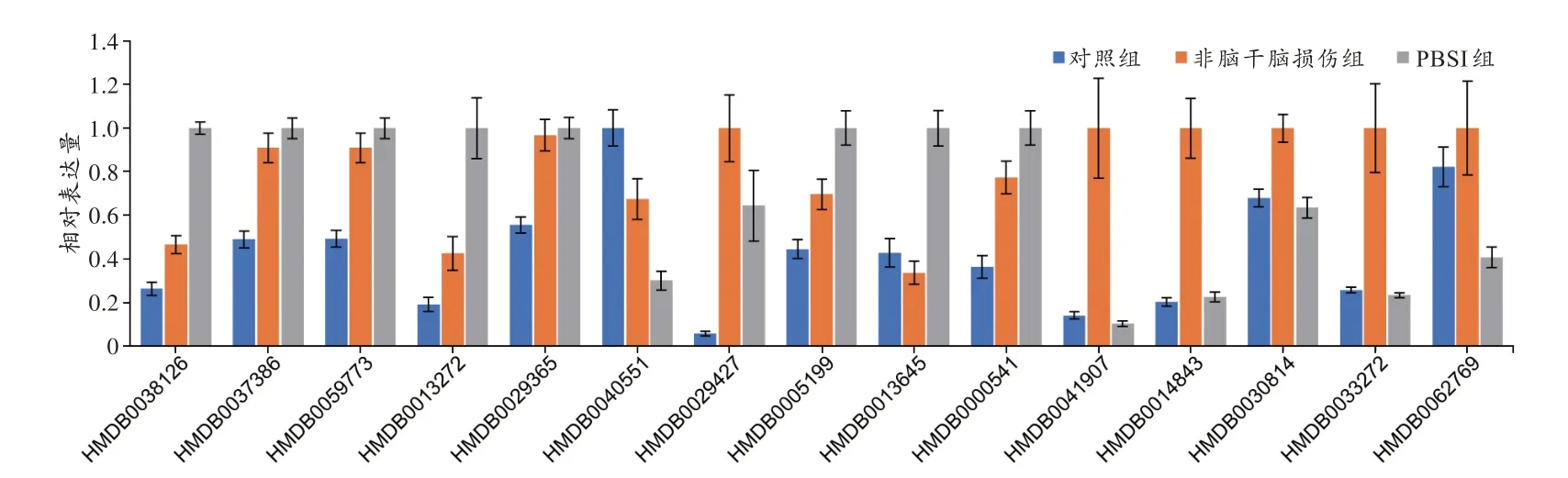
图8 15 种代谢标志物在不同死因大鼠脑干组织中的相对表达量Fig.8 Relative expression levels of 15 metabolic markers in the rat brain stem tissues with different causes of death
此外,为了排除PLS-DA 筛选结果对随机森林模型构建的影响,将72 个样本的818 种代谢标志物的丰度信息构建数据集,进行随机森林模型构建和预测(训练集为54 个样本,测试集为18 个样本)。模型对测试集数据的预测准确率为88.9%(图9A,表1)。变量的重要性排序中,占据前10 的为HMDB0038126(京尼平苷酸)、HMDB0041907[咪达普利(imidapril)]、HMDB0014843[地拉 韦啶(delavirdine)]、HMDB0040551(L-α-氨基-1H-吡咯-1-己酸)、HMDB0030814(Mammea E/BB)、HMDB0033272(dihydroclusin)、HMDB0029365(阿魏酰酪胺)、HMDB0013645(N,N-二甲基鞘氨醇)、HMDB0037386[(S)-薄荷酮-8-硫代乙酸酯]和HMDB0062769[ε-己内酰胺(epsilon-caprolactam)],见图9B。其中HMDB0041907(咪达普利)、HMDB0014843(地拉韦啶)、HMDB0030814(Mammea E/BB)、HMDB0033272(dihydroclusin)和HMDB0062769(ε-己内酰胺)均在非脑干脑损伤组中高表达(图8),另外5 种代谢标志物与利用PLS-DA 筛选出来的86 种代谢标志物进行随机森林建模变量的重要性占据前10 的代谢标志物重叠。
图9 818 种代谢标志物的随机森林模型构建结果Fig.9 Construction results of random forest model for 818 metabolic markers
3 讨论
本研究采用LC-MS 检测了大鼠3 种死因所致的脑干代谢组变化,基于PLS-DA 分析筛选出86 个VIP值≥1.0 的代谢标志物或直接用818 种代谢标志物进行数据分析,基于随机森林算法构建最佳的死因鉴别模型,并对这些代谢标志物在模型中的重要程度进行排序,最终发现HMDB0038126(京尼平苷酸)、HMDB0037386[(S)-薄荷酮-8-硫代乙酸酯]、HMDB0041907(咪达普利)等15 种代谢标志物起到重要的作用。
HMDB0038126(京尼平苷酸)最早于1964 年被发现[13],通常在饮料和水果中被检测到。京尼平苷酸属于环烯醚萜类,可有效抑制小胶质细胞释放一氧化氮,减少细胞中白细胞介素1β、肿瘤坏死因子α、前列腺素E2 的产生,从而抑制细胞炎症、保护神经元[14-15]。CHEN 等[16]研究发现,京尼平苷酸可上调神经突生长标志物,包括微管相关蛋白2(microtubule associated protein 2,MAP2)和生长相关蛋白43(growth associated protein 43,GAP43),从而促进神经突生长。此外,京尼平苷酸还可以作为降压活性成分发挥作用[17]。在PBSI 中,脑干组织损伤所引起的局部反应主要表现为细胞膜破坏,活性氧类物质和胞质内游离钙增多,缺氧,化学毒害和遗传物质变异等几方面,而京尼平苷酸可以保护神经元不受氧化应激等诱导的细胞毒性的影响,抑制细胞炎症,促进小胶质细胞降解和清除异常蛋白并促进神经元轴突再生[18]。因此,京尼平苷酸的生物学功能与本研究观察到的结果即PBSI 致脑干组织中出现京尼平苷酸上调是一致的。但由于京尼平苷酸可能是大鼠通过食物摄取进入体内的物质,不属于体内固有的代谢产物,其重要性有所削弱,在后续甄别应用中应加以注意。
HMDB0013272(N-月桂酰甘氨酸)被发现可作为诊断肝硬化的代谢组学生物标志物[19],在肝硬化患者体内其丰度显著提高[20]。HMDB0005199[(R)-去甲猪毛菜碱]是一种儿茶酚异喹啉类化合物,是多巴胺神经毒素{N-甲基(R)-去甲猪毛菜碱[N-methyl-(R)-salsolinol]}的前体[21],其合成水平受脑组织中多巴胺和乙醛浓度的影响。研究[22]表明,(R)-去甲猪毛菜碱可激活蛋白激酶B-哺乳动物雷帕霉素靶蛋白(protein kinase B-mammalian target of rapamycin,AktmTOR)信号通路来调节细胞内的自噬水平,从而引发神经毒性。HMDB0013645(N,N-二甲基鞘氨醇)是一种神经性疼痛诱导剂,可通过上调神经生长因子活性诱发神经性疼痛样行为[23]。结合本研究的结果,表明(R)-去甲猪毛菜碱和N,N-二甲基鞘氨醇的上调可能是由于PBSI 的应激作用引起。在代谢物重要性排序前10 中还有其他代谢物,目前虽暂不清楚其在PBSI 致死过程中的具体作用,但也提示其与PBSI致死相关。
此外,本研究利用随机森林算法进行死因识别建模,模型准确率最高达88.9%,这可能是由于每个死因组别中包含了死后6 h、1 d 和3 d 的样本,而样本中的代谢物水平会随着时间的变化而发生变化。若有足够的样本量来进行单个时间点的建模,相信预测准确率将有所提高。此外,两种随机森林模型的结果表明,基于所有代谢物建模的效果更好,这可能是由于PLS-DA 与随机森林算法对变量的筛选过程存在差异,利用更多变量进行随机森林建模,可提高模型预测的准确性。当然,本研究主要是基于大鼠模型,若在实际案例中,则需要使用人类尸体样本重新建模。
综上所述,本研究对PBSI、非脑干脑损伤和对照(剪头处死)组死后6 h、1 d 和3 d 的大鼠脑干组织样本进行了代谢组学分析,基于PLS-DA 和随机森林算法,成功建立了死因识别模型,并鉴定出与PBSI 相关的代谢标志物——京尼平苷酸、N-月桂酰甘氨酸、(R)-去甲猪毛菜碱和N,N-二甲基鞘氨醇,为法医学实践中脑干相关的死因鉴别提供了一定的参考。
