不同树种对森林土壤可溶性有机碳空间异质性的影响
2023-10-15孔令迁刘宪斌
孔令迁,刘宪斌,2,李 云
(1玉溪师范学院化学生物与环境学院,云南玉溪 653100;2玉溪师范学院生物与环境工程研究院,云南玉溪 653100)
0 引言
由于森林生态系统地上部植被和地下部土壤共同固定了大量碳元素,且不断通过各种物理过程、化学作用和生物化学合成与分解过程与周围环境进行着大通量碳元素交换,使其成为全球陆地生态系统的重要组成部分[1-2]。土壤体作为森林生态系统中各种物质交换和能量流动的源与库,是森林生态系统物种进化、结构组成、功能调整和生命演变的重要控制因子之一,尤其是作为减缓全球温度升高对森林碳库产生的消极影响的缓冲媒介,土壤生态系统的结构和组成一直是相关研究领域的热点[1,3]。土壤有机碳是土壤有机质的核心组成部分,是碳元素在土壤剖面中的主要存在状态,对全球气候变化较为敏感,其动态变化对森林生态系统的物种组成、功能调整和发展演化方向产生显著影响[4]。从内容物组成角度讲,土壤有机碳是由不同分解速率和分解程度的土壤碳组分组成的复杂混合体,主要来源是生态系统地上部植物有机体的凋落和地下部植物根系的分泌物和脱落物,其组成和贮量主要受植被类型、气候条件、土壤环境和人类活动的干扰[4-5]。土壤可溶性有机碳属于土壤有机碳复合体中比较活跃的部分,其作为土壤微生物活动的能源和土壤肥力的标志物,可直接参与土壤生物化学转化过程,对土壤碳库平衡、土壤物理化学性质、土壤肥力保持等方面都具有重要作用[4,6]。因此,研究森林生态系统土壤可溶性有机碳的组成、分布、贮量和相关环境控制因子不仅对了解和学习森林生态系统碳循环和养分循环有很大的帮助,而且对研究森林生态系统植被组成和功能多样性同样具有重要意义。
从宏观角度分析,不同植物群落或森林植物分布带由于所处地理位置各种环境因子组合不同,不同种类森林生态系统之间土壤可溶性有机碳组成和贮量有显著性差异;同一种类植物群落或森林植物分布带在不同年龄段由于其固碳能力、生长量和功能多样性不同,造成其在产生和保存土壤活性有机碳方面的能力也有所不同;同一种植物群落或森林植物分布带、同一个生长年龄阶段、但是在一年中不同生长季节期间,由于受到不同季节间植被生长周期和环境因子组合不同的影响,其土壤可溶性有机碳的组成和贮量也不相同[4,6]。从微观角度分析,在同一个森林生态系统的同一个山谷流域中,不同坡位、方向和植被种类组合造成植物生长环境中的水分、光照和温度等环境因子的差异,从而导致土壤深度、质地组成、微生物和动物含量与种类的不同,使不同微环境条件下的土壤可溶性碳含量也有明显差异,从而造成明显的土壤可溶性有机碳的林间土壤空间异质性[7]。刘涛泽等[8]在广西环江县南部的喀斯特地区坡地土壤研究中发现不同坡向和坡地表层土壤可溶性有机碳含量不同,主要受到有机质输入、地形和土壤质量等因素的综合影响;袁知洋等[9]在江西萍乡武功山草甸区植被小群落下取样调查,发现乔木、灌木、高草和地被植物4 种不同植物群落下土壤可溶性有机碳含量互相之间均达到显著差异性水平。这些研究成果只能证明不同类型植物带或植物聚集区域土壤活性有机碳的分布规律,但对于受人为干扰因子较小、植物种类多样的原生林生态系统说服力较小。本项目以云南哀牢山中山湿性常绿阔叶原生林生态系统为试验场所,选择3 个具有代表性的山谷流域,在每个山谷流域上游森林生态系统原生土区域选择7 个森林生态系统主要树种作为调查对象,在其根际范围内采集0~10 cm 土壤样品,测定土壤可溶性有机碳和土壤有机碳的含量,观察不同树种对土壤可溶性有机碳和土壤有机碳空间异质性的影响。本项目的主要研究内容包括3个方面:(1)调查不同树种根际范围内土壤可溶性有机碳和土壤有机碳的含量;(2)分别对比不同山谷样地中同一个树种个体根际范围内土壤可溶性有机碳含量和土壤有机碳含量之间的差异;(3)分析不同树种间和不同山谷样地中土壤可溶性有机碳和土壤有机碳的比例变化规律。
1 材料与方法
1.1 样地信息
本项目土壤样品野外采集样地设置在云南省哀牢山北段上部徐家坝地区(24°32′ N,101°01′ E,海拔2400~2600 m,隶属于普洱市景东县太忠乡)的亚热带中山湿性常绿阔叶原生林生态系统,该森林生态系统位于哀牢山国家级自然保护区的核心位置,也是中国科学院哀牢山亚热带森林生态系统研究站和云南哀牢山森林生态系统国家野外科学观测研究站的位置所在地[10-11]。该区域主要受西南季风气候和垂直山地气候的影响,年均降雨量1931 mm,蒸发量1485 mm,空气相对湿度85%,林间常年有地表径流且随着季节和降雨量的变化而改变径流量,在山谷间地势较平坦的地方累积了大量淤积土并形成小面积草地生态系统[11-12]。年内干湿季节分明,雨热同期,5—10 月为雨季,降雨量大且集中,常常有大雨天气,降雨量占全年总降雨量的85%左右;11月—次年4月为干季,降雨量占全年总降雨量的15%左右[11-12]。年均温度11.3℃,7月份平均气温最高,为16.4℃;1月份平均气温最低,为5.4℃,冬季常常受冷空气和大雪的干扰,全年无霜期200 d左右[11,13]。
该区域森林生态系统主要包括山地上游海拔较高的原生林生态系统和山谷中间海拔较低的草地生态系统2 种[10,14]。山地上游原生林生态系统土壤类型为原生土,森林植被主要由壳斗科、茶科、樟科和木兰科等树种组成,其代表树种有硬壳柯[Lithocarpus hancei(Bentham) Rehd.]、木 果 柯[Lithocarpus xylocarpus(Kurz) Markg.]、变色锥[Castanopsis rufescens(Hook.f.et Thoms.) Huang et Y.T.Chang]、南洋木荷(Schima noronhaeReinw.ex Bl.Bijdr)、翅 柄 紫 茎(Stewartia pteropetiolataW.C.Cheng)、云南连蕊茶[Camellia forrestii(Diels) Coh.St.]、黄 心 树(Machilus gambleiKing ex Hook.f.)、黄丹木姜子[Litsea elongata(Wall.ex Nees)Benth.et Hook.f.]、红花木莲[Manglietia insignis(Wall.) Blume]、多花含笑(Michelia floribundaFinet et Gagn.)等;山谷中间海拔较低的草地生态系统土壤类型为淤积土,由于常年积水和上游原生土的累积,不利于高大乔木的生长,植被类型主要包括生长速度较快的草本植物和适于积水条件的矮小灌木植物,其植被代表种类主要有弯蕊开口箭[Tupistra wattii(C.B.Clarke) Hook.f.]、纤细冷水花(Pilea gracilisHand.-Mazz.)、长柄异药花(Fordiophyton longipesY.C.Huang ex C.Chen)、紫茎泽兰[Ageratina adenophora(Spreng.R.M.King et H.Rob.)]、珍 珠 花(Lyonia ovalifoliaHort.Ex Gard)、金 丝 桃(Hypericum uralumBuch.)、岩 子 果(Gaultheria forrestiiDiels)、朝 天 罐(Osbeckia opiparaC.Y.Wu et C.Chen)等[14-16]。原生林生态系统林间原生土土壤母质由古生代板岩、微晶片岩、绿泥片岩、石英片岩、片麻岩、闪长岩和石英岩等组成,风化发育后形成现在典型的山地黄壤、棕壤和黄棕壤,风化程度高,土层深厚,土层深度可达120~180 cm[10,17]。森林生态系统植被物种丰富,年生长量大,林间地表长年覆盖有大量凋落物(平均3~7 cm厚),土壤疏松而肥沃,土壤孔隙度较高,土壤微生物种类丰富;土壤中含有大量有机质和矿质养分,0~30 cm土层中pH 4.2~4.5,有机质含量为5%~9%,土壤有机碳、氮、磷和钾的含量分别为5.03%、0.35%、0.07%、0.37%;0~30 cm 土层的C/N 比为14.4,30~50 cm 土层的C/N比为15.3[10,18-19]。
1.2 采样方法
本项目的野外土壤采集样地选择以徐家坝地区杜鹃湖为中心,在其东、南和北3个方向分别选取一个可以充分代表当地原生林生态系统植被类型和环境条件的山谷作为土壤样品采集样地,所选取的3 个山谷分别为:老君山神(样地中心地理位置为:24°32′49.68″N,101°01′58.50″ E;海拔2513 m;坡向朝西南;坡度22.3°)、三棵树(样地中心地理位置为:24°32′25.28″N,101°01′40.29″ E;海拔2514 m;坡向朝东北;坡度38.3°)和山门口(样地中心地理位置为:24°32′48.38″N,101°01′21.37″ E;海拔2511 m;坡向朝东;坡度26.6°)[10]。在对该3 个山谷样地前期野外调查工作的基础上,根据森林生态系统中每个树种的相对优势度和重要值,确定7 个树种作为本项目的研究对象,包括:硬壳柯、变色锥、云南连蕊茶、云南越桔[Vaccinium duclouxii(Levl.) Hand.-Mazz.]、木果柯、多花山矾(Symplocos ramosissimaWall.ex G.Don)和南洋木荷[11-12]。在每个山谷中,在1 hm2(100 m×100 m)的面积范围内分别选择上述每一种研究对象个体10株,选择标准是保证同一个树种所选择的10 株个体生长情况相对一致,能够代表3 个山谷样地绝大多数本种树种的个体生长情况,便于保证后期研究结果同一个树种测定结果的相对一致性和3个不同山谷样地之间同一个树种测定结果的可比性(表1)。

表1 云南哀牢山中山湿性常绿阔叶原生林生态系统7个采样树种不同个体生物学特征
由于野外试验样地地表覆盖有较厚的植被凋落物(主要是落叶和细小树枝),凋落物和矿质土壤接触面土壤湿度较高,便于土壤微生物和土壤动物的生存和繁衍,被分解的有机物较多,在表层土壤中富集有大量土壤有机质和矿质营养,前期的研究结果也证明该森林生态系统林间表层土的土壤有机碳、活性有机碳、微生物量碳、可溶性有机碳和水溶性有机碳在整个土壤剖面中含量均最高且有一致的规律可循,林间表层土壤样品的碳和矿质养分状况可以代表土壤的整体状况进行调查和样本间互相对比,所以本项目中所采集的土壤样品均为0~10 cm 的表层土[10,20]。野外采集土壤样品时,先对每一棵选定的调查树种个体进行标号,以其树干为中心、以树干和树冠边缘1/2 的距离为半径画圆,在圆圈的边线上,在树干东、南、西、北4 个方向分别用环刀采集0~10 cm 的表层土土壤样品,充分混合均匀后装入自封袋进行标号,并及时送回野外试验站进行保存,用以后期实验室内测定土壤可溶性有机碳和土壤有机碳的含量。
1.3 指标测定
本项目中土壤有机碳数据采用的是重铬酸钾氧化-外加热方法测定,土壤可溶性有机碳数据用TOC自动分析仪(Vario TOC cube)测定[8,21-22]。
1.4 数据分析
数据用Excel 2017 进行前期分析处理和后期作图,用SPSS 20.0软件进行统计学单因素方差分析。
2 结果与分析
2.1 土壤可溶性有机碳
土壤可溶性有机碳数据在3 个山谷样地之间、每个山谷样地中的7 个树种之间差异显著性比较明显(图1和表2)。在老君山神山谷样地中,硬壳柯和南洋木荷2 个树种根际范围内土壤可溶性有机碳含量最高,为80.00 g/kg 左右;其次为木果柯,其根际范围内土壤可溶性有机碳含量为(60.33±4.87)g/kg;然后是变色锥和多花山矾2个树种;云南连蕊茶和云南越桔2个灌木树种根际范围内的土壤可溶性有机碳含量最低,为30.00 g/kg左右(图1a)。在三棵树山谷样地中,7个树种根际范围内土壤可溶性有机碳含量的排序从大到小依次为:硬壳柯≈南洋木荷>变色锥≈木果柯≈多花山矾>云南越桔≈云南连蕊茶(图1b)。在山门口山谷样地中,硬壳柯、南洋木荷和木果柯3个树种根际范围内的土壤活性有机碳含量最高,为60.00 g/kg左右;其次为变色锥和多花山矾,其根际范围内土壤可溶性有机碳含量为46.00 g/kg左右;云南连蕊茶和云南越桔2个灌木树种根际范围内的土壤可溶性有机碳含量最低,为30.00 g/kg左右(图1c和表2)。
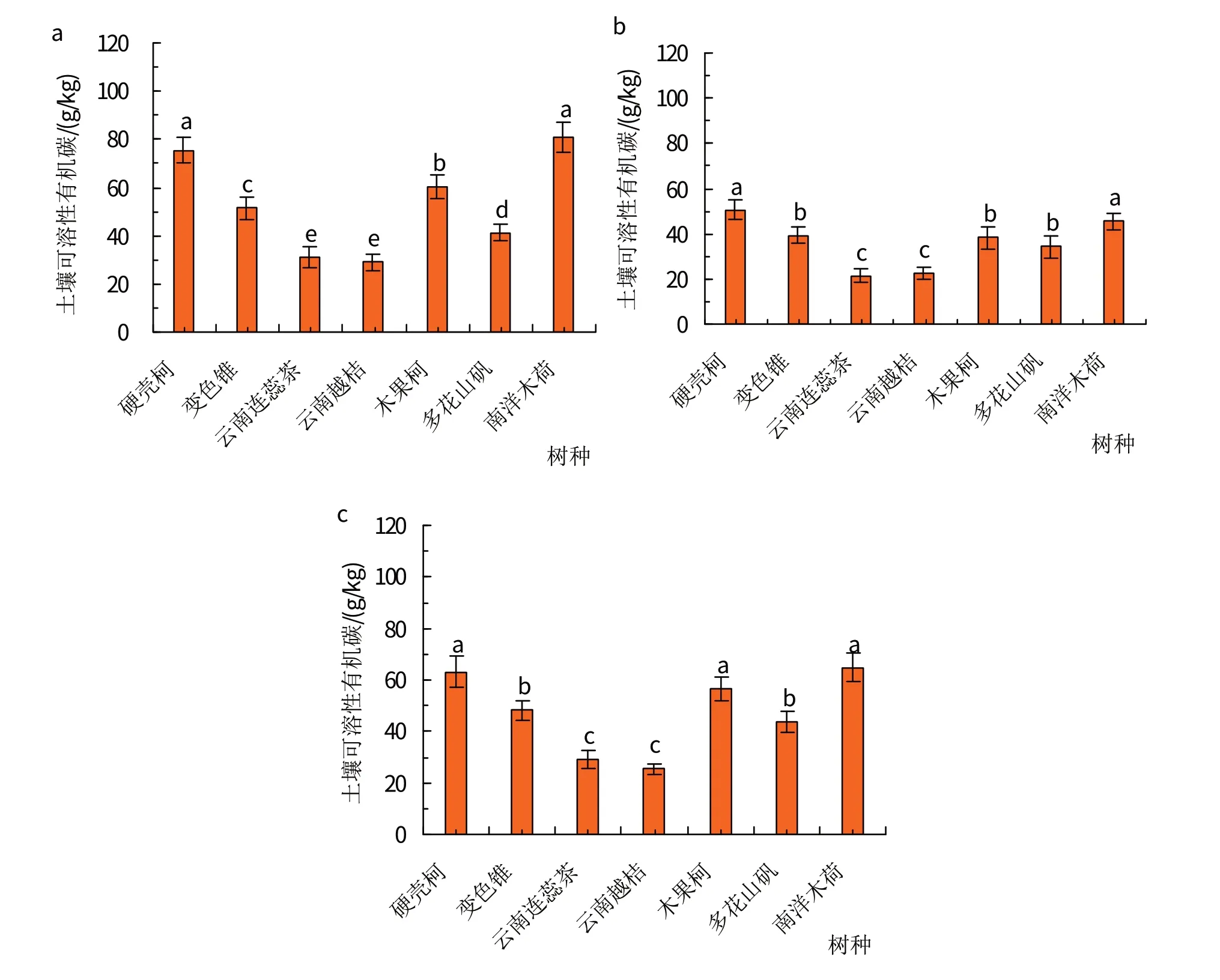
图1 云南哀牢山中山湿性常绿阔叶原生林生态系统老君山神(a)、三棵树(b)和山门口(c)3个样地中7个不同树种根际土壤可溶性有机碳含量显著性分析
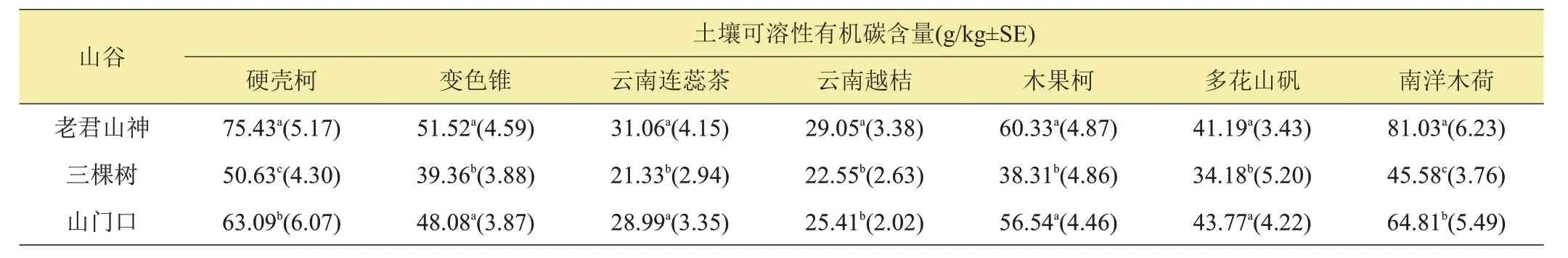
表2 云南哀牢山中山湿性常绿阔叶原生林生态系统7个不同树种根际土壤可溶性有机碳含量在老君山神、三棵树和山门口3个山谷样地中的显著性分析表
从同一个树种在3个不同山谷样地中土壤可溶性有机碳含量角度比较,硬壳柯和南洋木荷2 个树种根际范围内的土壤可溶性有机碳在老君山神山谷样地中含量最高,其次为山门口山谷样地,三棵树山谷样地中的土壤可溶性有机碳含量最低,分别为(50.63±4.30) g/kg 和(45.58±3.76) g/kg;变色锥、云南连蕊茶、木果柯和多花山矾等4 个树种根际范围内土壤可溶性有机碳在老君山神和山门口2 个山谷样地中的含量差异不显著,但在三棵树山谷样地中含量均降低到最低值;云南越桔树种根际范围内土壤可溶性有机碳含量在老君山神山谷样地中达到最大值,为(29.05±3.38)g/kg,但在三棵树和山门口2 个山谷样地中差异不明显,为24.00 g/kg左右(表2)。
2.2 土壤有机碳
与上述土壤活性有机碳的林下土壤空间异质性调查结果相似,土壤有机碳数据调查结果在3 个山谷样地、7 个不同树种根际范围内的变化也比较明显(图2和表3)。在老君山神山谷样地中,硬壳柯、木果柯和南洋木荷3 个树种根际范围内土壤有机碳的含量最高,高达120 g/kg左右;其次为变色锥和多花山矾2个树种,其根际范围内土壤有机碳含量为90 g/kg 左右;云南越桔根际范围内的土壤有机碳含量最低,为(78.52±6.63)g/kg;云南连蕊茶树种根际范围内土壤有机碳含量为(85.03±6.55)g/kg,低于变色锥和多花山矾但高于云南越桔,只是与这3 个树种根际范围内土壤有机碳的含量差异还没有达到统计学意义上的显著性(图2a)。在三棵树山谷样地中,7 个调查树种根际范围内土壤有机碳含量的排序从高到低依次为:硬壳柯≈木果柯≈南洋木荷>变色锥≈多花山矾>云南越桔,云南连蕊茶根际范围内土壤有机碳含量为(76.93±6.49)g/kg,介于变色锥和多花山矾2 个树种与云南越桔之间,只是均没有达到统计学意义上的差异显著性(图2b)。在山门口山谷样地中,硬壳柯、木果柯、多花山矾和南洋木荷等4个树种根际范围内土壤有机碳的含量显著性差异不明显,为108 g/kg左右;云南越桔根际范围内土壤有机碳含量最低,为(75.09±7.12)g/kg;变色锥和云南连蕊茶2个树种根际范围内的土壤全碳分别为(96.48±6.41)g/kg 和(85.91±5.72)g/kg(图2c 和表3)。

图2 云南哀牢山中山湿性常绿阔叶原生林生态系统老君山神(a)、三棵树(b)和山门口(c)3个样地中7个不同树种根际土壤有机碳含量显著性分析

表3 云南哀牢山中山湿性常绿阔叶原生林生态系统7个不同树种根际土壤有机碳含量在老君山神、三棵树和山门口3个山谷样地中的显著性分析表
在3 个山谷样地中,硬壳柯和南洋木荷2 个树种根际范围内土壤有机碳含量在老君山神山谷样地中达到最大值,分别为(122.24±15.47) g/kg 和(120.53±12.16) g/kg,分别显著高于三棵树山谷样地中的(97.29±8.97) g/kg 和(91.19±7.72) g/kg,山门口山谷样地中2 个树种根际范围内土壤有机碳含量介于老君山神和山门口2 个山谷样地之间,但含量差异从统计学意义上分析不显著;变色锥、云南连蕊茶和木果柯等3 个树种根际范围内的土壤有机碳含量分别在老君山神和山门口2 个山谷样地中含量差异不明显,但均显著高于其在三棵树山谷样地中根际范围内土壤有机碳的含量;云南越桔根际范围内土壤有机碳含量在3 个山谷样地中差异均不显著,为75.00 g/kg 左右;然而,多花山矾根际范围内土壤有机碳含量在山门口山谷样地中达到最大值,为(101.73±8.10) g/kg,在三棵树山谷样地中降低为最小值,为(82.86±8.34) g/kg(表3)。
2.3 土壤可溶性碳所占全碳比例
相对于7个研究树种根际范围内土壤可溶性有机碳含量和土壤有机碳含量在3个山谷样地中变化的多样性,其根际范围内可溶性有机碳和土壤有机碳的比例在3 个山谷样地中的变化规律较为一致(表4)。硬壳柯、变色锥、云南连蕊茶、云南越桔和木果柯等5 个树种根际范围内土壤可溶性有机碳含量占土壤有机碳含量的比例在老君山神山谷样地中分别达到最大值,分别为:62.32%±3.95%、55.76%±3.03%、36.52%±5.31%、36.84%±3.70%和54.22%±2.51%,且在三棵树山谷样地中分别降低至最小值;多花山矾根际范围内土壤可溶性有机碳含量占土壤有机碳含量的比例在3个山谷样地中差异不显著,为43.00%左右;南洋木荷根际范围内土壤可溶性有机碳含量占土壤有机碳含量的比值在3 个山谷样地中的排序从大到小依次为:老君山神>山门口>三棵树。

表4 云南哀牢山中山湿性常绿阔叶原生林生态系统老君山神、三棵树和山门口3个山谷样地中7个不同树种根际土壤可溶性有机碳含量占土壤有机碳含量比例显著性分析表
3 讨论
作为土壤有机碳的重要组成部分,土壤可溶性有机碳是土壤有机碳所有组成成分中最活跃的成分,是由一系列来自于植物有机体、被土壤微生物和土壤动物不同程度分解的有机物组成,从最简单的各种小分子有机酸到复杂的大分子物质,如单糖和多糖、氨基酸、氨基糖、胡敏酸和富里酸等[5,23]。作为土壤轻组碳的一部分,土壤可溶性有机碳组成成分从其来源分析比较复杂,可能含有的成分包括:部分死亡微生物释放出的碳、处于不同腐烂程度的植物和土壤动物残留物及其分解产物,和易分解的非腐殖有机物质,如碳水化合物、多糖、蛋白质、有机酸、氨基酸、蜡、脂肪酸和其他非特异性含碳化合物[5,24-25]。从土壤可溶性有机碳的定义、范畴和组成成分可以判断其来源,其去向则包括:随重力作用或土壤微生物和土壤动物的活动进入土壤剖面的下层土壤、随降雨或地下水淋溶流失、随着地表径流或泥石流迁移到下游地区、被土壤微生物和土壤动物消化利用、被植物根系吸收利用、被土壤黏粒包裹或被土壤团聚体固定发生土壤矿化作用而长久固定[26-28]。土壤可溶性有机碳含量是土壤生态系统有机碳输入、分解和输出动态发展过程的平衡结果,从而影响土壤有机碳输入和输出的所有环境因子都会影响土壤可溶性有机碳的含量。森林生态系统土壤有机碳输入来源主要包括地上部植被凋落物、地下部根系分泌物和根系死亡脱落物,其中地上部植被凋落物的贡献量最大。据研究,森林生态系统每年通过地上部植被凋落物层向地下矿质土壤剖面输送可溶性有机碳量为115~500 kg/hm2,占生态系统年凋落物总量的35%左右[29]。进一步研究发现,新近或新鲜凋落物和土壤腐殖质是森林生态系统土壤可溶性有机碳的主要来源,主要原因是新近或新鲜凋落物中易于被分解的内容物多为植物细胞内容物和简单细胞组织,这一部分有机物也是可溶性物质,而土壤腐殖质中绝大部分有机物被完全分解,其中可溶性有机碳易于被溶解而进入矿质土壤[30-31]。而影响土壤可溶性有机碳在土壤有机碳中的比例的因素除了输入物本身的质量和可溶性有机碳含量以外,能够影响输入物分解的环境因子还包括:土壤温度、土壤含水量、土壤pH、降雨强度与降雨量、土壤矿质元素含量、土壤微生物量和土壤动物数量等[5,24-25]。因此,一个自然生态系统土壤可溶性有机碳含量的研究要综合考虑生态系统地上部和地下部各种生物和非生物环境因子,才能充分解释在试验过程中所观察到的现象。
在本项目的3个野外土壤样品采集样地中,7个采样树种中作为乔木树种的硬壳柯、变色锥、木果柯、多花山矾和南洋木荷等5个树种个体根际范围内表层土壤可溶性有机碳含量和土壤有机碳含量高于云南连蕊茶和云南越桔2种灌木植物,可能的原因包括:(1)5种乔木树种的年凋落物输入量远远大于2 种灌木植物,导致乔木树种根际范围内通过凋落物分解进入表层土壤的土壤有机碳和土壤可溶性有机碳含量超过灌木植物[32];(2)5 种乔木植物的地下植物根系比2 种灌木植物发达,乔木植物根系生长量较大,通过乔木植物根系输入土壤中的根系分泌物和脱落的死亡根系量比灌木植物多[33];(3)乔木作为林冠上层树种,其叶片较林下灌木树种叶片日均接触光照时间长、光照强度大、进行光合作用时间长、合成有机物量大,作为凋落物累积到林间土壤表面后,释放出的可溶性有机物含量比灌木植物叶片量大[30];(4)乔木植物蒸腾量较林下灌木植物大,地下根系较林下灌木植物发达,由地上部叶片产生的蒸腾拉力促使地下部植物根系从土壤中吸收的各种矿质养分含量较林下灌木植物大,使林下灌木植物长期处于生长受抑制状态,土壤根系与土壤微生物形成对养分的竞争关系,两者处于一种此消彼长的关系,不利于灌木树种根际范围内地表凋落物的分解和可溶性有机物的释放[34]。而在5 种乔木树种中,硬壳柯和南洋木荷2个树种的土壤可溶性有机碳和土壤有机碳含量较高,分析其原因,硬壳柯在3个山谷样地中相对优势度高,重要值大,树种在3 个山谷样地中生长优势大;南洋木荷树种虽然个体数在3个山谷样地中少,但是单株个体生长量大,林冠下凋落物较多,通过凋落物进入土壤中的可溶性有机碳和有机碳多[11]。3 个山谷样地之间进行比较,老君山神山谷样地平均坡度为22.3°,林下有大量草本植物,地表累积大量植被凋落物,平均厚度为5~10 cm,凋落物和表层矿质土壤接触面土壤含水量较高,土壤微生物量和土壤动物数量大,便于凋落物分解;三棵树山谷样地平均坡度为38.3°,局部区域甚至超过50°,致使植被凋落物难以在陡峭坡面上累积,大部分凋落物受重力作用和雨季地表径流冲刷作用而累积到下游山谷中间平坦地带的淤积土壤中,致使样地内多有裸露矿质土壤,导致表层土壤含水量低,地被草本植物少,地表碳输入量少;而山门口山谷样地环境条件介于老君山神山谷样地和三棵树山谷样地之间,导致其样地内各个树种根际范围内土壤可溶性有机碳含量、土壤有机碳含量介于老君山神山谷样地和三棵树山谷样地同种树种之间。
云南哀牢山国家级自然保护区内的森林生态系统属于亚热带中山湿性常绿阔叶原生林生态系统,其主要特征是森林面积大、历史时间长、受人为干扰因素小、保存完好、以云南当地特有植物种为优势种和建群种[19,35]。此森林生态系统植物物种数量多、森林结构功能稳定,在保持水土、保护生物多样性、维护生态环境、提供生态系统服务、调节地区局部气候条件、促进人类社会经济发展、固定碳元素和平衡地区与全球碳循环的功能等方面发挥着重要的作用[14]。在哀牢山森林生态系统研究站的支持下,来自中国科学院西双版纳热带植物园、中国科学院昆明植物研究所、云南大学和云南农业大学等高校和科研院所相关研究人员以此森林生态系统为研究平台,发表了一系列关于地上部森林生态系统结构功能和物种组成、地质地貌和历史演变、气候变化与环境条件改变、地下部土壤生态系统组成和对环境的响应等方向和领域的研究成果[14,36]。然而,在山地森林生态系统中,陡峭的地势地貌和复杂的地质环境条件对植被种类分布、土壤理化性质和物质循环也常常产生显著影响。本项目聚焦土壤可溶性有机碳和土壤有机碳的空间异质性,结合7 个具有代表性的不同树种和3 个不同地势地貌的山谷样地,探讨和分析不同树种对土壤可溶性有机碳和土壤有机碳空间异质性的影响,为哀牢山中山湿性常绿阔叶原生林生态系统土壤不同属性碳和土壤养分空间异质性的研究提供新的研究方向和研究思路,为该森林生态系统相关研究领域提供新的研究内容,为全球同种类型森林生态系统的相关领域研究也有着重要的提示作用。
4 结论
(1)土壤可溶性有机碳和土壤有机碳在不同树种根际范围内表层土壤中的含量具有明显空间异质性。按照不同树种角度分析,乔木植物种类的硬壳柯、变色锥、木果柯、多花山矾和南洋木荷等5个树种根际范围内表层土壤可溶性有机碳含量和土壤有机碳含量明显高于灌木植物种类的云南连蕊茶和云南越桔2个树种根际范围内土壤可溶性有机碳含量和土壤有机碳含量;在乔木植物中,硬壳柯、南洋木荷和木果柯3 个树种根际范围内的土壤可溶性有机碳和土壤有机碳含量又明显高于其他2 个树种;说明不仅乔木植物和灌木植物两种不同类型树种影响林间表层土壤可溶性有机碳和土壤有机碳的空间异质性,而且同样为乔木种类的不同树种对土壤可溶性有机碳和土壤有机碳的含量也有影响。7个树种根际范围内土壤可溶性有机碳和土壤有机碳在老君山神和山门口2个山谷样地中的含量和比例明显高于三棵树山谷样地,说明不同山谷样地中的环境条件不仅对植被根际范围内土壤可溶性有机碳和土壤有机碳含量和分布有显著影响,而且还能够影响土壤可溶性有机碳在有机碳含量中的比值。
(2)本项目中,不同树种对林间表层土壤可溶性有机碳空间异质性的影响和不同采集土壤样地对土壤可溶性有机碳和土壤有机碳含量的影响结果与前人研究结论相符,说明本项目中测定的数据和相关研究结果可信。在同一个受人为干扰因素较小的原生林生态系统不同树种根际范围内采集土壤样品测定可溶性有机碳和土壤有机碳含量、讨论不同树种和采集土壤样地对土壤可溶性有机碳和土壤有机碳空间异质性的影响,在云南哀牢山中山湿性常绿阔叶原生林生态系统中尚属首次,也是对同种类型和其他类型森林生态系统中相关研究领域的有效补充,为全球热带和亚热带各种不同类型森林生态系统中不同属性土壤碳的空间异质性研究提供新的研究思路和方向。
(3)本项目只采集了调查树种0~10 cm 的表层土壤样品进行土壤可溶性有机碳含量和土壤有机碳含量的测定,并没有涉及到2 种不同属性土壤碳沿着土壤整体剖面的含量和垂直分布规律。考虑到乔木种类植物和灌木种类植物、不同种类乔木植物之间植物根系在土壤剖面中的分布规律和分泌物的量和组成的不同,不同树种对土壤剖面中各个土层可溶性有机碳和土壤有机碳的含量和分布规律应该产生不同的影响,这是本项目的不足之处,也是下一步的工作内容。
