硬粒、马齿玉米单倍体自交结实株率差异研究
2023-10-13李忠南王越人马艺文李志博邬生辉曲海涛李福林张淑琴李光发
李忠南 王越人 马艺文 李志博 邬生辉 曲海涛 李福林 张淑琴 李光发
(1.吉林省农业技术推广总站 吉林 长春 130033;2.通化市农业科学研究院 吉林 梅河口 135007)
玉米单倍体育种技术是现代玉米育种的重要技术之一,以诱导系对材料进行杂交诱导为主要方法[1-3]。由于基础材料遗传背景等不同, 无论在海南自然加倍还是本地化学加倍, 单倍体自交结实株率种质间存在很大差异。 雄穗加倍率低始终是单倍体育种的瓶颈[4-6]。 近期研究表明,在1 号、3 号、4 号、5 号、7 号染色体上存在23 个雄穗高自然加倍相关QTL 位点,1 号、3 号、4 号染色体上定位到4 个共有显著QTL区段[7]。
10 余年的单倍体育种实践表明, 在相继发现玉米单倍体自交结实株率红轴对白轴、 紫叶鞘对绿叶鞘及父本血缘对母本血缘的显著优势[8-11]之后,发觉硬粒杂交材料单倍体自交结实株率极明显低于马齿。 由于硬粒×马齿为东北地区玉米主导杂优模式,研究探讨硬粒、 马齿杂交材料单倍体自交结实株率差异,对单倍体育种具有重要意义,该方面研究未见报道。
本文作者以硬粒和马齿共23 份玉米F1单倍体进行3 年的海南自然加倍试验, 并就7 份进行本地化学加倍试验, 通过研究探讨硬粒和马齿不同杂交方式单倍体自交结实株率差异,以提高育种效率。
1 材料与方法
1.1 试验材料配制及诱导
杂交配制基础材料F1的自交系或DH 系共23 份,其中, 硬粒种质8 份, 分别为PH6WC、NG7015、29、Y3011、D135-2、1258-6、B20、X826-2,马齿种质15 份(表1)。 2019 年杂交配制材料6 份:D10×PH6WC、D10×郑58、D135-2×PH4CV、29396×D1279、D1279×D752、B20×D752;2020 年 杂 交 配 制 材 料10 份:NG7015×29、PH6WC×Y3011、D1279×D956、1258-6×PH4CV、 T12111×D1279、 PH12TB×D1279、 T0322×D1279、PHB1M×D1279、A6×D1279、X826-2×D1279;2021 年杂交配制材料7 份: F1硬粒(PH6WC×29)×PH6WC、 Y3011×29、 763-1×D752、 763-1×D1279、PHB1M×D1279、D338-2×763-1、D494×PHB1M, 共计23 份F1材料。 2020 年、2021 年、2022 年进行杂交诱导,秋季挑选单倍体粒。 诱导系采用吉诱SM6278-2。试验在通化市农业科学研究院试验地进行。
1.2 海南自然加倍和本地化学加倍
2020 年、2021 年、2022 年10 月25 至11 月3 日,在海南三亚南滨农场通化市农业科学研究院试验基地,进行单倍体自然加倍试验,采用大垄双行,垄距1 m,单粒播种,株距10 cm。苗期开始去杂,授粉前查数单倍体株数。 于当年12 月中旬开始套小袋并开始授粉,月末结束,查数授粉株数;于下一年2 月中旬收获自交结实穗,晾晒并查数。 海南单倍体加倍试验圃周边作物为水稻、棉花、蔬菜等。
本地化学加倍试验采用晚播错期处理。2021 年、2022 年在通化市农业科学研究院试验地就7 份单倍体进行化学加倍试验,均于5 月10 日播种,9 月20 日收获并查数自交结实穗。 具体方法:单倍体籽粒用清水浸泡8~10 h,再用恒温箱催芽,催芽均匀;芽根长至1.5 cm 左右时,用小刀片切根尖;切完后用纱网袋装好并写好标签,用秋水仙素0.08%蒸馏水溶液浸泡8 h 后拿出,用清水浸泡、冲涮5 次,待播种。 滴灌防止土壤表层干硬,以提高出苗率。
1.3 统计分析
数据分析、系统聚类分析、回归分析和方差分析均采用DPS 17.10 进行。 数据转换方式为对数转换,聚类距离为卡方距离,聚类方法为可变类平均法;多重比较采用新复极差比较法。
自交结实株率(%)=自交结实株数/单倍体株数×100。
2 结果与分析
2.1 单倍体自交结实株率
由表2 可知, 总体上单倍体自交结实株率不符合正态分布(P<0.05),变异系数为78.63%;马齿×马齿单倍体自交结实株率高低不一,其平均值、最大值及最小值均高于其他处理,变异系数为42.97%,不符合正态分布(P<0.05);硬粒×硬粒平均值、最大值及最小值均最低, 变异系数为77.55%, 符合正态分布(P>0.05); 硬粒×马齿介于中间偏低, 变异系数为59.38%,符合正态分布(P>0.05)。表明马齿种质比硬粒种质富含优良结实性基因,杂交重组累加效应高。
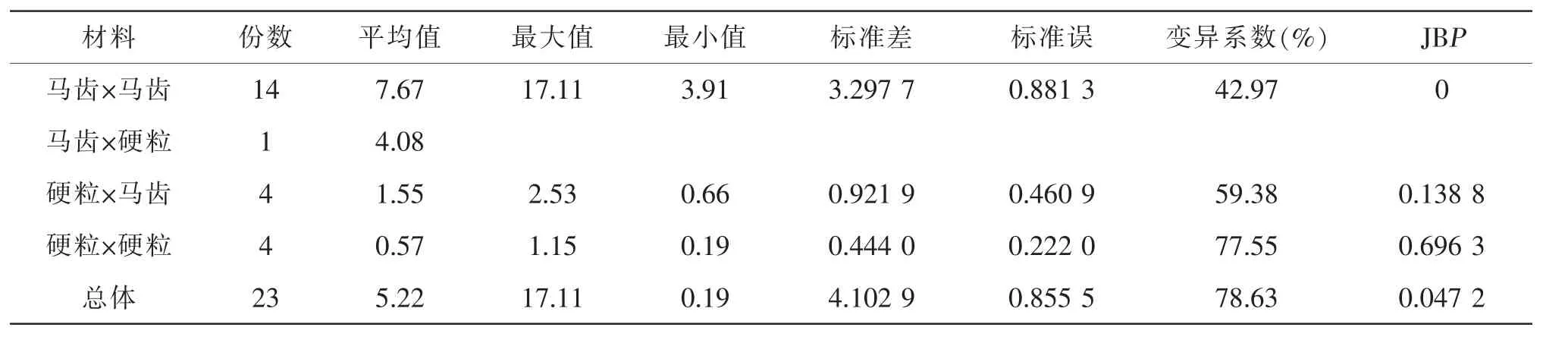
表2 单倍体自交结实株率统计参数
由表3 可知,马齿×马齿本地化学加倍比海南自然加倍自交结实株率提高2.36%~14.55%, 硬粒×硬粒提高0.67%~1.49%,马齿×硬粒提高2.49%,硬粒×马齿提高2.1%。 化学加倍对马齿×马齿效果好,说明杂交重组累加效应高。 利用自然加倍和化学加倍自交结实株率回归分析表明,相关系数0.966 6(P<0.01),决定系数0.934 3, 剩余标准差0.330 7, 截距A:1.204 2(P<0.01),斜率B:0.684 5(P<0.01),y=1.204 2+0.684 5x,种质间化学加倍与自然加倍自交结实株率呈线性关系, 说明单倍体自交结实株率高低由杂交重组累加优良结实性效应基因数量多寡决定。
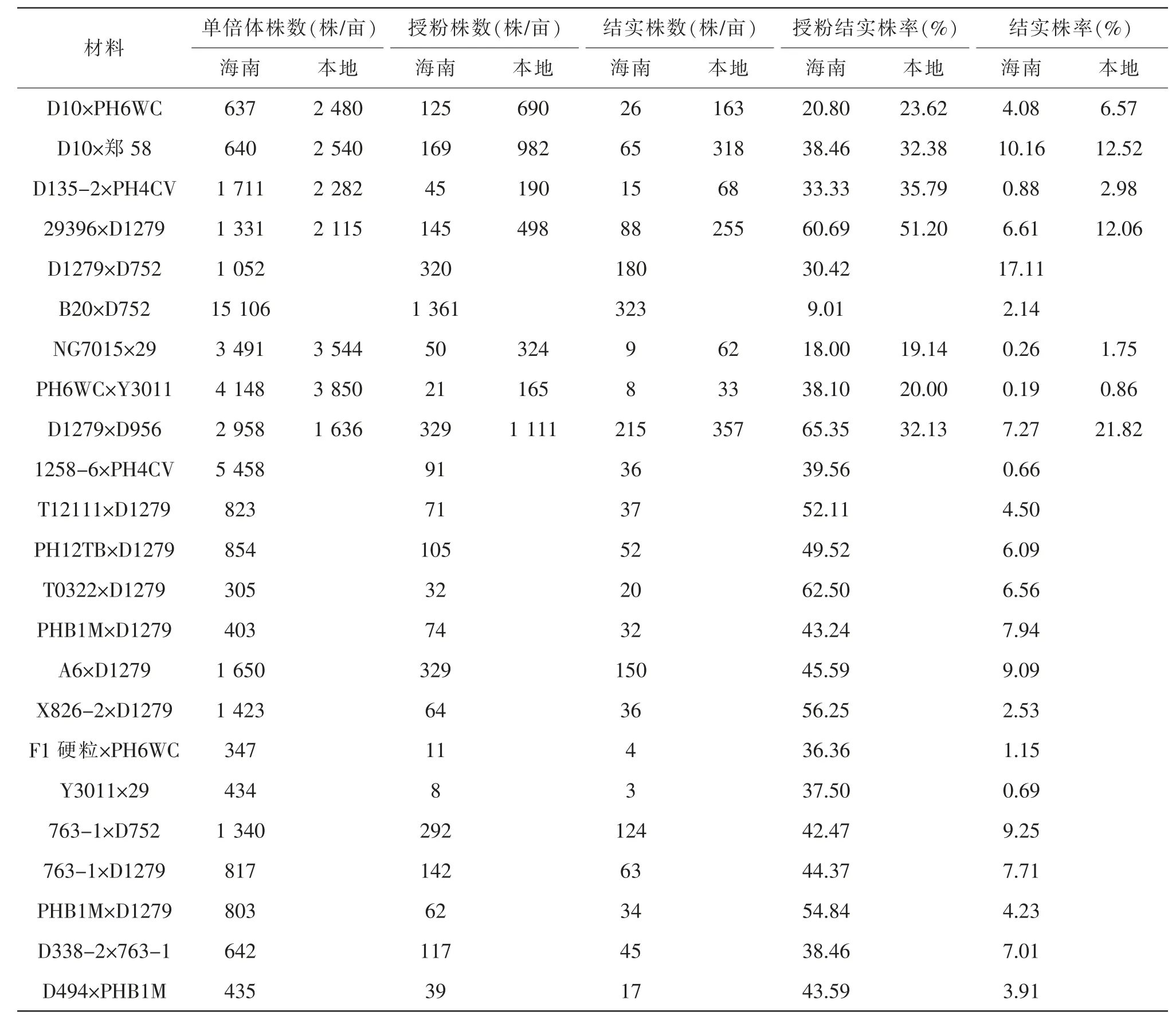
表3 玉米单倍体海南自然加倍与本地化学加倍自交结实株率
2.2 聚类分析
针对23 份材料海南单倍体株数和自交结实株率进行聚类分析,结果见图1、表4。 表4 列出了材料间卡方距离,RSQ 为残差平方和,值越大,关系越近;值越小,关系越远;PSF 为伪F 统计量层次距离中的参数;WS 为统计量,值越大,关系越远。 图1 说明卡方距离在1.50 条件下,可划分为2 个群。 群1 包括硬粒×硬粒4 份和硬粒×马齿4 份,与马齿群最大卡方距离为2.4762; 群2 包括马齿×马齿14 份和马齿×硬粒1 份。说明群1 和群2 在优良结实基因数量上和杂交重组累加效应上存在差异,存在细胞质遗传效应。

表4 卡方距离及参数
在卡方距离0.75 条件下, 群1 可划分为3 个次群:次群1、2 分别为硬粒×硬粒(各2 份),次群3 为硬粒×马齿(4 份);群2 可划分为2 个次群:次群1 为马齿×马齿(5 份),为高自交结实株率群,次群2 为马齿×马齿和马齿×硬粒(10 份)。
前期研究结果[14]:PH4CV×A6(马齿×马齿)F1单倍体自交结实株率14.70%,育成D752(马齿);B20×PH4CV(硬粒×马齿)F1单倍体自交结实株率6.90%,育成D1279(马齿),说明PH4CV 和育成的马齿DH系富含优良结实性基因, 存在细胞质遗传效应。 群2 次群1 中的D1279×D752 单倍体自交结实株率为17.11%, 说明优良结实性基因杂交重组累加效应明显。 群1 次群3 中硬粒×马齿4 份杂交材料自交结实株率:0.66%(1258-6×PH4CV)、2.14%(B20×D752)、2.53%(X826-2×D1279)、0.88%(D135-2×PH4CV),杂交材料父本均富含优良结实性基因。 可见,硬粒种质寡含优良结实性基因,杂交重组累加效应差,存在细胞质遗传效应。尤其是D135-2(红轴昌7-2×PH4CV)×PH4CV,PH4CV 占75%,说明细胞质效应明显。
群1 次群1 中的F1硬粒×PH6WC(1.15%),其F1硬粒(PH6WC×29,硬粒×硬粒)前期研究结果[9,14]单倍体自交结实株率分别为0.73%和0.60%;PH6WC 自身单倍体结实率0.20%[14];PH6WC 占75%,说明硬粒种质寡含且杂交重组累加优良加结实性基因效应微弱。 D10(马齿)选于PH6WC×郑58(硬粒×马齿),前期研究结果[14]自交结实株率1.70%~4.10%。群2 次群2 中的D10×PH6WC 为4.08%,PH6WC 占75%;D10×郑58 为10.16%,郑58 占75%,卡方距离0.240 3。 说明马齿种质富含且杂交重组累加优良结实性基因效应明显,郑58 富含优良结实性基因。
2.3 方差与差异显著性比较分析
将2020 年、2021 年海南自然加倍和2021 年、2022 年化学加倍4 份、3 份杂交材料单倍体自交结实株率进行方差和差异比较分析,结果见表5、表6。由表5 可知,区组间P<0.05、P<0.05;材料间P<0.01、P<0.05,说明本地化学加倍自交结实株率比海南自然加倍高,达显著水平;材料间自交结实株率差异达极显著水平或显著水平。 材料间的标准差及标准误已去掉区组效应,结果可用[12]。
由表6 可知, 自交结实株率D10×郑58 与29396×D1279 差异不显著, 与D10×PH6WC 差异达显著水平, 与D135-2×PH4CV 差异达极显著水平;29396×D1279 与D10×PH6WC 差 异 不 显 著, 与D135-2×PH4CV 差异达显著水平;D10×PH6WC 与D135-2×PH4CV 差异达显著水平。即硬粒×马齿<马齿×硬粒达显著水平;硬粒×马齿<马齿×马齿达极显著或显著水平。 D1279×D956 与NG7015×29 差异达显著水平,与PH6WC×Y3011 差异达极显著水平;NG7015×29 与PH6WC×Y3011 差异不显著, 即硬粒×硬粒<马齿×马齿达显著或极显著水平。
进一步研究表明, 马齿种质富含且杂交重组累加优良结实性基因效应明显, 硬粒种质寡含且杂交重组累加优良结实性基因效应微弱, 存在细胞质遗传效应。
3 讨论与结论
3.1 讨论
玉米祖先为硬粒, 经驯化和人工选择遗传演化为马齿, 硬粒分布于美洲北部, 马齿分布于美洲南部。 美洲硬粒传到欧洲,发展成欧洲硬粒。 我国老地方品种多为硬粒型。 硬粒种质具有种子耐低温、早熟、脱水快及容重高等优点[15-17],对我国北方玉米生产具有重要作用。 玉米杂种优势利用以来,硬粒×马齿一直为主导杂优模式,尤其近百余年的种质改良,马齿比硬粒具有更宽的遗传背景、 更丰富的遗传多样性,对玉米产量贡献更大[18]。 从5 000 年左右的进化时间跨度看, 马齿种质比硬粒更富含优良结实性基因。
本研究结果, 玉米单倍体自交结实株率为数量性状,马齿×马齿材料平均值最高,不符合正态分布;硬粒×硬粒材料最低,并存在细胞质遗传效应。 表明马齿种质富含且杂交重组累加优良结实性基因效应明显, 硬粒种质寡含且杂交重组累加优良结实性基因效应微弱,硬粒×马齿<马齿×硬粒。 马齿种质杂交材料, 自交结实株率双DH 系高于单DH 系表达,支持了一些研究结果[19];高自然加倍材料吉Gjb335DH3为马齿型[7],也证明了这一点。 遴选出富含优良结实性基因马齿种质PH4CV、D1279、D752、郑58。
对前期研究[14]的77 份材料进行统计表明(表7)符合本研究结论。 马齿×马齿呈更大的离散幅度,变异系数最高,不符合正态分布(P<0.01),平均值略低,由均为基础材料导致; 硬粒×马齿最大值6.90%(B20×PH4CV),PH6WC×PH4CV(先玉335,硬粒×马齿)单倍体自交结实株率6.71%[19],比较相近,应该是优势群间产量杂种优势导致,有待于进一步研究。 如果该二组合以PH4CV 为母本,其单倍体自交结实株率将会更高,有待于今后或同行验证。
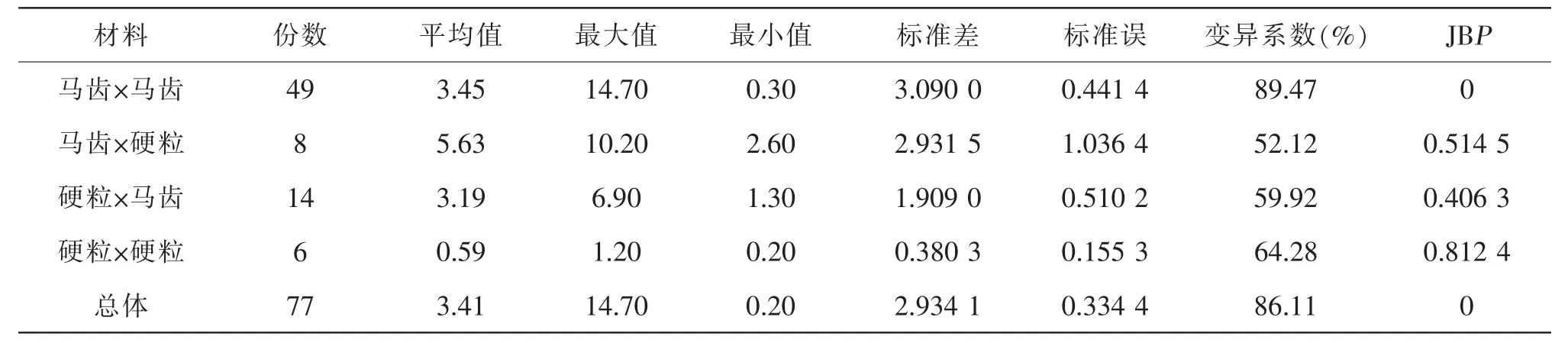
表7 自交结实株率统计参数
硬粒×马齿为东北地区主导杂优模式,选育硬粒高容重母本系是必然,利于种子耐低温、保全苗[20],同时确保单交种容重≥720 g/L(国家审定标准)。 本研究重要意义在于:在单倍体育种中,对于硬粒母本系选育应采取马齿×硬粒杂交方式, 在DH 系群中选择性状优良硬粒或近硬粒DH 系;对于硬粒×硬粒,应采用化学加倍或常规选育方法; 父本系选育应采取马齿×硬粒或马齿×马齿。如此将显著提高育种效率。随着单倍体育种轮回选择进度的加快,硬粒、马齿DH种质不同程度重组累加更多优良结实性基因, 将不同程度地表达更高自交结实株率, 极大推进种质创新工作。
3.2 结论
本研究表明, 马齿种质富含且杂交重组累加优良结实性基因效应明显,其单倍体自交结实株率高;硬粒种质寡含且杂交重组累加优良结实性基因效应微弱,其单倍体自交结实株率低。 单倍体自交结实率为数量性状, 马齿×马齿不符合正态分布; 硬粒×马齿<马齿×硬粒,存在细胞质遗传效应,硬粒×硬粒遗传效应最低。 马齿×马齿、马齿×硬粒重组累加优良结实性基因效应明显。
