甜菜碱对高温胁迫下茄子幼苗生理特性的影响
2023-10-12高晓萍牛天航常有麟刘思恬颉建明
高晓萍,张 婧,牛天航,刘 阳,常有麟,刘思恬,颉建明
(甘肃农业大学 园艺学院,甘肃 兰州 730070)
茄子(SolanummelongenaL.)起源于古印度[1],因其果实富含花青素、维生素和各类矿物质营养,深受大众喜爱[2]。茄子适宜生长温度为22~30 ℃,超过35 ℃会导致花芽分化不良、果皮失去光泽、产生僵果等[3],影响茄子产量和品质。在日光温室生产中,晴天正午常有超过40 ℃的现象,对茄子植株造成高温伤害。已有研究表明,外源喷施2,4-表油菜素内酯[4]、褪黑素[5]等物质可以减轻高温胁迫对茄子植株的伤害,但因其投入较高,在生产中应用较少。
甜菜碱(glycine betaine, GB)是存在于植物、细菌和动物中的一类季铵类水溶性生物碱,最早在甜菜中发现[6],在稳定蛋白质的四级结构方面有重要作用[7]。外源甜菜碱可以促进作物生长[8],改善植物水分状态[9],提高脯氨酸积累[10]、水分利用效率与气孔导度(stomatal conductance,Gs)[11],还可以保护光化学反应中心PSⅡ免受热诱导失活的影响[12]。作为一种渗透保护剂,甜菜碱可改善植物在逆境胁迫条件下的生长和生理状况。甜菜碱参与一系列非生物胁迫,如干旱胁迫、高温胁迫、低温胁迫、水分胁迫、金属胁迫、盐胁迫等。在非生物胁迫下,一定浓度的外源甜菜碱通过提高过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)等多种抗氧化酶活性来激活植物的抗氧化系统,保护植物免受氧化伤害,减少逆境对植物造成的伤害[13]。研究表明,当外源甜菜碱浓度为0.5~2.0 mmol·L-1时可以显著缓解高温胁迫对菜豆造成的伤害[14];喷施0.5 mmol·L-1甜菜碱能够提高葡萄幼苗的抗热性[15];高温胁迫下万寿菊叶面施用0.5、1.0 mmol·L-1的甜菜碱减轻了光抑制,缓解了热应激诱导的万寿菊CO2同化率、Gs、相对含水量和蒸腾速率(transpiration rate,Tr)的降低[16]。外源施用甜菜碱能增强番茄种子萌发和幼苗生长过程中对高温的耐受性,可能与甜菜碱介导的热休克基因表达增强有关[17]。热应激下大麦种子吸收的甜菜碱易位到幼苗后,通过增加净光合速率(net photosynthesis rate,Pn)、降低细胞膜通透性和热胁迫下重要离子的泄漏,增强它们保持更高水分含量和更高幼苗活力的能力[18]。甜菜碱在转基因番茄植株中的积累通过增强热反应基因的表达和热激蛋白的积累,从而提高番茄对热应激的耐受性[19]。叶面喷施20 mmol·L-1甜菜碱可提高芥菜的品质和产量[20]。
研究外源甜菜碱对茄子热损伤的缓解机制,有助于避免或减轻高温对茄子生长的影响。本研究对高温胁迫下茄子幼苗叶面喷施不同浓度甜菜碱,分析茄子幼苗生物量积累、光合特性、卡尔文循环关键酶活性、抗氧化酶活性、生物膜伤害与渗透调节物质的变化,旨在探讨甜菜碱对高温胁迫下茄子幼苗生长生理的缓解作用,以期为茄子抵御高温危害提供理论与技术依据。
1 材料与方法
1.1 试验地点与材料
本试验在培养室和人工气候箱(RDN-400E-4,浙江宁波)进行。供试茄子品种为新娘(长茄,甘肃主栽品种),购自酒泉夏禾种业有限责任公司;甜菜碱(纯度>99%,CAS:107-43-7)购自上海源叶生物有限责任公司。
1.2 试验设计
挑选大小均一、质地饱满的茄子种子,于55 ℃温水中浸种后,播种于9 cm×9 cm的营养钵中,栽培基质中草炭、蛭石、珍珠岩体积比为3∶1∶1,每钵播1粒种子。出芽后,置于(28±1)℃(昼)/(18±1)℃(夜)的气候箱内培养,光照强度为20 000 lx。待幼苗长至四叶一心时,叶面喷施不同浓度的外源甜菜碱。试验共设置4个处理,分别为CK(对照,清水处理,含0.1%吐温20)、T1(20 mmol·L-1甜菜碱)、T2(40 mmol·L-1甜菜碱)、T3(60 mmol·L-1甜菜碱)。连续处理3 d后,将气候箱温度持续性升至(40±2)℃,高温处理48 h。各处理随机挑选9株测定生长指标,测定幼苗生长点向下第2片功能叶的生理指标。
1.3 指标测定
1.3.1 生长指标测定
茄子幼苗高温胁迫48 h后,测量植株高度,以及全株、地上部分、地下部分鲜重。于105 ℃烘箱中杀青30 min后,调至80 ℃烘至质量不变,测定各部分干重。
1.3.2 根系形态参数测定
将茄子幼苗根系从根原基处剪断,用根系扫描仪(型号Epson Expression 11000XL)扫描根系图片,用WinRHIZO软件分析根长、根尖数、根体积和根表面积。
1.3.3 叶片气孔形态观察
茄子叶片气孔观察。在高温胁迫48 h后,采集功能叶,每处理设3个重复。参照Min等[21]的方法对样品进行预处理;避开主叶脉将叶片切成5 mm×5 mm的正方形,用4%戊二醛固定液在室温下浸泡2 h,然后用磷酸缓冲液(0.10 mol·L-1PBS,pH值6.8)冲洗4次,每次间隔10 min。反复冲洗后,用一系列不同体积分数的乙醇(30%、40%、50%、60%、70%、75%)依次对叶片脱水,每个梯度冲洗20 min,然后置于75%乙醇中过夜;第2天,采用不同体积分数(75%、80%、85%、90%、95%)乙醇对样本进行脱水,每次20 min,然后转入100%乙醇中脱水3次,每次30 min。最后用一系列不同体积分数(30%、50%、70%、80%、85%、90%、95%、100%、100%、100%)的叔丁醇洗脱乙醇,每次30 min。样品干燥后喷涂一层薄薄的金粉,使用扫描电子显微镜(SEM,Hitachi-S3400N)观察气孔形态并拍照。
1.3.4 叶绿素含量与叶绿素荧光参数测定
叶绿素含量测定参照李静[22]的方法。高温处理48 h后,选取完全展开的功能叶片,剪碎混匀,称取0.1 g于试管中,用10 mL 80%丙酮提取叶绿素。测定663 nm、645 nm波长的吸光度,根据以下公式算出叶绿素a(Chl a)含量、叶绿素b(Chl b)含量、总叶绿素(Chl a+b)含量和Chla/Chlb的值。
ca=[12.71×D663-2.59×D645]×(V/1 000m);
(1)
cb=[22.88×D645-4.67×D663]×(V/1 000m);
(2)
ca+b=[20.29×D645+8.04×D663]×(V/1 000m);
(3)
式(1)~(3)中:ca、cb、ca+b分别代表叶绿素a含量、叶绿素b含量、总叶绿素含量的值;D663、D645分别代表663、645 nm处的吸光度;V和m分别代表提取物总体积(mL)和样品鲜重(g)。
叶绿素荧光参数测定参照胡琳莉[23]的方法。各处理随机选择3株幼苗,经30 min暗适应后,剪下第3片完全展开的功能叶置于荧光仪测定台上,使用调制叶绿素荧光成像仪IMAGIN-PAM(Heinz WaItz, Germany)测定。测量光光量子通量密度为0.1 μmol·m-2·s-1,光化光量子通量密度为81 μmol·m-2·s-1,饱和脉冲光量子通量密度为2 700 μmol·m-2·s-1;脉冲光时间0.8 s,每20 s打1次饱和脉冲光,共打光15次。暗反应下的初始荧光(original fluorescence,Fo)、最大荧光产量(maximal fluorescence,Fm)通过打饱和脉冲光获得,计算出最大光化学效率(maximum quantum efficiency of PSⅡ,Fv/Fm);在光化光持续供给300 s后,在光化光下获得稳态荧光Fs;在饱和脉冲光发出0.8 s后得到光下最大荧光产量Fm′,计算实际光化学效率ΦPSⅡ、光化学猝灭系数(photochemical quenching coefficient,qP)、非光化学猝灭系数(non-photochemical quenching coefficient,NPQ)。
1.3.5 光合参数的测定
茄子幼苗植株处理后,选择植株功能叶测定茄子幼苗光合气体交换参数。采用CIRAS-2(PP SYSTEM Inc, Amesbury, MA 01913, USA)便携式光合仪测定,参数包括Pn、Gs、胞间CO2浓度(intercellular CO2concentration,Ci)、蒸腾速率(transpiration rate,Tr)。
1.3.6 卡尔文循环酶活性测定
选取展开的功能叶用液氮速冻后存储于-80 ℃冰箱中待测。果糖-1,6-二磷酸酶(FBPase)、1,5-二磷酸核酮糖羧化酶(Rubisco)、果糖1,6-二磷酸醛缩酶(FBA)、转酮醇酶(TK)和3-甘油醛-磷酸脱氢酶(GAPDH)活性采用ELISA(酶联免疫法)试剂盒测定(上海远慕,中国)。
1.3.7 抗氧化酶活性测定
超氧化物歧化酶(SOD)活性使用SOD活性检测试剂盒测定;过氧化物酶(POD)活性使用POD活性检测试剂盒测定;过氧化氢酶(CAT)活性使用CAT活性检测试剂盒测定。检测试剂盒均购于索莱宝生物科技有限公司。
1.3.8 脯氨酸和丙二醛(MDA)含量测定
脯氨酸含量测定采用磺基水杨酸法;MDA含量测定用硫代巴比妥酸法,均参照陈刚等[24]的方法。
1.3.9 超氧阴离子组织化学(NBT)染色
茄子叶片NBT染色参照夏凌君[25]的方法。取甜菜碱处理后的茄子幼苗叶片,置于锥形瓶中,量取NBT染液(0.5 mg·mL-1,用pH值7.8的PBS溶解)加入其中,真空泵抽真空30 min;25 ℃暗置孵育6 h;孵育完成后,倒出染液,加入脱色液(乳酸、甘油、无水乙醇体积比为1∶1∶3),沸水浴5 min,直至叶片中绿色完全褪去为止。用扫描仪扫描染色叶片并观察。
1.4 数据分析
采用IBM SPSS Statistics 26.0软件,用LSD和Duncan’s检验法对有显著差异(P<0.05)的数据进行多重比较,利用Excel 2021软件制图和统计数据。
2 结果与分析
2.1 外源甜菜碱对高温胁迫下茄子幼苗生长的影响
由图1可知,高温胁迫下CK的茄子幼苗长势较弱,甜菜碱处理后茄子幼苗长势增强,其中T2处理长势最好。在高温胁迫下,不同浓度的甜菜碱处理均能够不同程度地促进茄子幼苗的生长(表1)。随着甜菜碱浓度的增加,高温胁迫下茄子幼苗的株高、全株干重、全株鲜重、地上部干重和地上部鲜重均呈先增加后降低趋势。与CK相比,T2处理显著提高了茄子幼苗株高、全株鲜重、全株干重、地上部鲜重、地上部干重,分别提高32.97%、34.17%、11.11%、23.71%、15.49%。
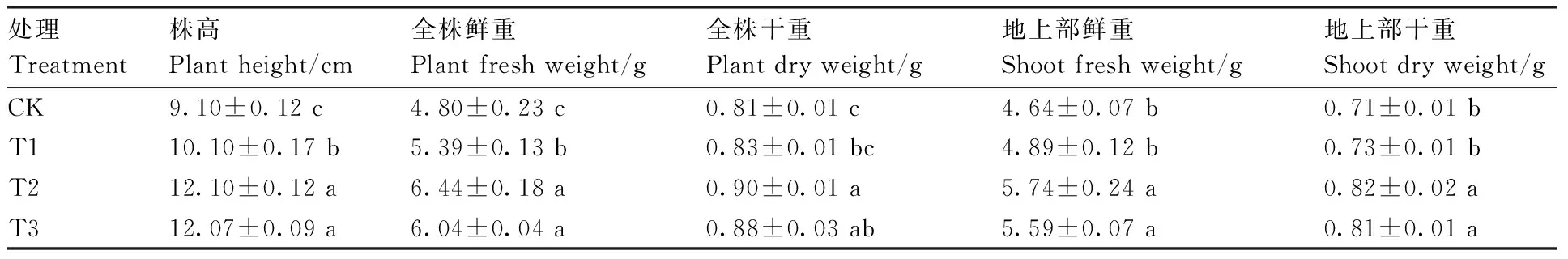
表1 高温胁迫下甜菜碱对茄子幼苗生长的影响

CK、T1、T2、T3处理的甜菜碱浓度分别为0、20、40、60 mmol·L-1。下同。The glycine betaine concentrations of CK, T1, T2 and T3 were 0, 20, 40 and 60 mmol·L-1, respectively. The same as below.图1 高温胁迫下不同浓度甜菜碱处理的茄子幼苗表型Fig.1 Phenotype of eggplant seedlings treated with different concentrations of glycine betaine under high temperature stress
2.2 外源甜菜碱对高温胁迫下茄子幼苗根系形态结构的影响
与CK相比,高温胁迫下甜菜碱处理后的茄子幼苗根系生长旺盛,其中,T2处理的根系生长最好(图2)。高温胁迫下,不同浓度甜菜碱处理可不同程度地促进茄子幼苗根长、根表面积、根体积和根尖数的增加(表2)。随着甜菜碱浓度的增加,高温胁迫下茄子幼苗根长、根表面积、根体积、根尖数呈先增加后降低趋势。与CK相比,T2处理的根长、根表面积、根体积和根尖数均显著提高,分别提高19.47%、16.55%、32.20%和25.29%。
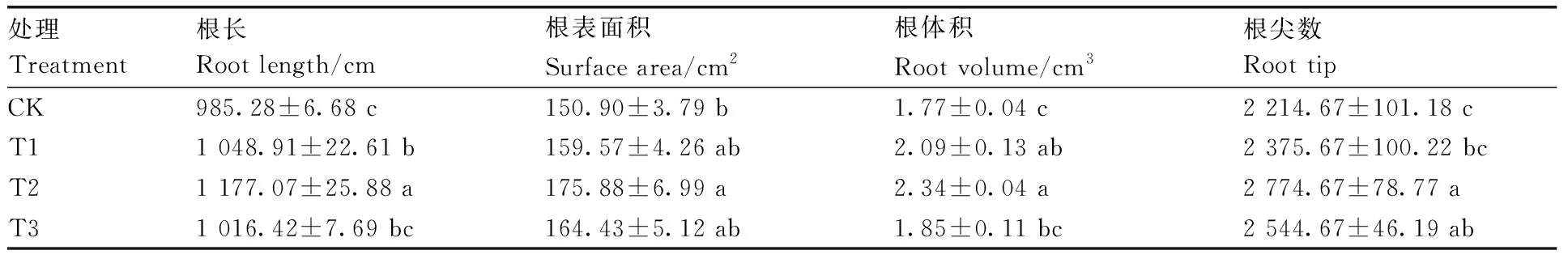
表2 甜菜碱对高温胁迫下茄子幼苗根系的影响
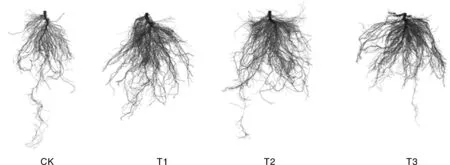
图2 高温胁迫下不同浓度甜菜碱处理的茄子幼苗根系形态Fig.2 Root morphology of eggplant seedlings treated with different concentrations of glycine betaine under high temperature stress
2.3 外源甜菜碱对高温胁迫下茄子光合色素和光合作用的影响
2.3.1 光合色素含量
由表3可知,随着甜菜碱浓度的增加,茄子幼苗叶绿素含量呈先增加后降低趋势。与CK相比,T2处理的茄子幼苗叶绿素a含量、叶绿素b含量、总叶绿素含量和叶绿素a/b均显著增加,分别增加35.26%、12.00%、28.68%和20.87%。

表3 高温胁迫下不同浓度甜菜碱处理的茄子幼苗光合色素含量
2.3.2 光合气体交换参数
由表4可知:T2和T3处理较CK均可显著提高茄子幼苗净光合速率,分别提高51.26%和31.03%;与CK相比,T1、T2和T3处理的Gs和Tr均显著增加,Gs分别增加25.51%、40.24%和33.29%,Tr分别提高23.92%、49.86%和39.19%;T1、T2、T3处理均显著降低了叶片的Ci,与CK相比,分别降低4.54%、14.70%和3.22%。

表4 高温胁迫下不同浓度甜菜碱处理的茄子幼苗光合特性
2.4 外源甜菜碱对高温胁迫下茄子幼苗卡尔文循环关键酶活性的影响
不同浓度的外源甜菜碱均可促进高温胁迫下茄子幼苗卡尔文循环相关酶活性(图3)。随着甜菜碱浓度的增加,高温胁迫下茄子幼苗叶片的FBPase、Rubisco、FBA、GAPDH和TK活性呈现先升高后下降的趋势。与CK相比较,T2处理茄子幼苗叶片的FBPase、Rubisco、FBA、GAPDH和TK活性均显著提高,分别提高31.83%、35.64%、34.50%、18.20%、46.61%。
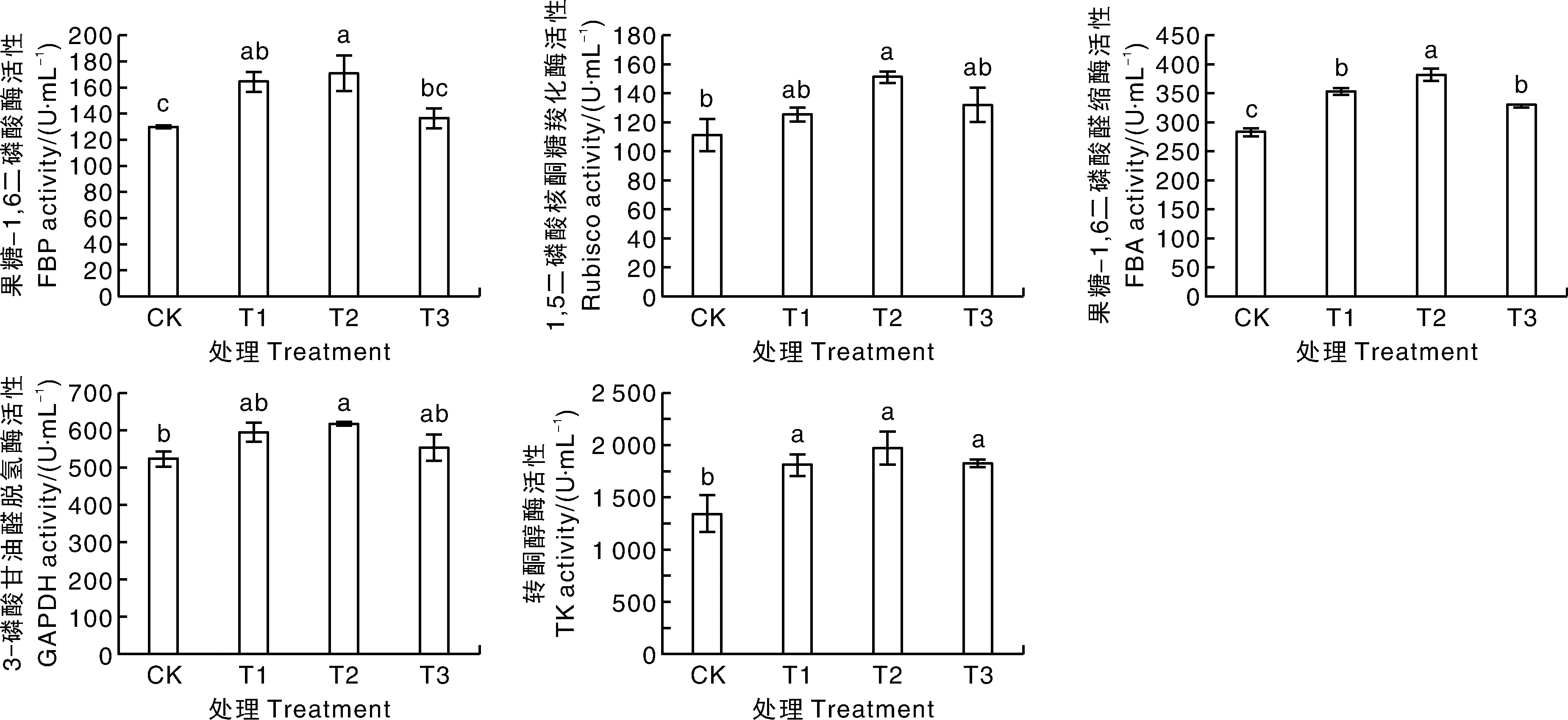
不同处理间没有相同小写字母表示差异显著(P<0.05)。下同。The bars of different treatments without the same lowercase letters indicated significant difference (P<0.05). The same as below.图3 高温胁迫下甜菜碱处理的茄子幼苗光合作用关键酶活性Fig.3 Key enzyme activity of photosynthesis in eggplant seedlings treated with glycine betaine under high temperature stress
2.5 外源甜菜碱对高温胁迫下茄子幼苗叶片气孔形态特性的影响
外源甜菜碱对高温胁迫下茄子幼苗叶片气孔形态特性的影响如图4所示。CK的茄子幼苗叶片保卫细胞失水皱缩、气孔关闭,喷施适宜浓度的甜菜碱后保卫细胞恢复正常,T1、T2、T3处理的叶片气孔呈不同程度张开状态。

图4 高温胁迫下甜菜碱处理的茄子幼苗气孔形态Fig.4 Stomatal morphology of eggplant seedlings treated with glycine betaine under high temperature stress
2.6 外源甜菜碱对高温胁迫下茄子幼苗叶片叶绿素荧光的影响
图5为高温胁迫下不同浓度甜菜碱处理后茄子幼苗叶片Fv/Fm的荧光成像,表明高温胁迫降低了Fv/Fm,但对叶脉影响较小,叶面喷施甜菜碱后减轻了PSⅡ的光抑制程度。由表5可知,随着甜菜碱浓度的增加,茄子幼苗叶片的叶绿素荧光参数Fv/Fm、ΦPSⅡ、qP均有不同程度的提高,NPQ则呈降低趋势。与CK相比,T2处理叶片的Fv/Fm、ΦPSⅡ、qP均显著提高,分别提高16.69%、13.18%、7.15%,NPQ降低了15.73%。
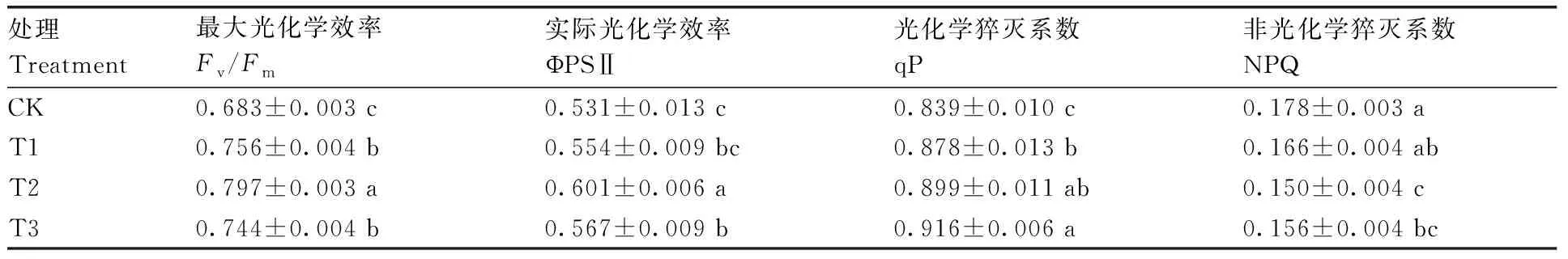
表5 高温胁迫下不同浓度甜菜碱处理的茄子幼苗叶绿素荧光参数

图5 高温胁迫下甜菜碱处理的茄子幼苗叶片Fv/Fm的荧光成像Fig.5 Fluorescence imaging of Fv/Fm in leaves of eggplant seedlings treated with glycine betaine under high temperature stress
2.7 外源甜菜碱对高温胁迫下茄子抗氧化酶活性的影响
由图6可知,在高温胁迫下喷施甜菜碱均能够显著提高茄子幼苗的SOD、POD和CAT活性。与CK相比,T1、T2、T3处理下幼苗叶片SOD活性分别显著提高26.46%、35.62%和29.65%。与CK相比,T2处理茄子幼苗POD和CAT活性分别提高109.65%和72.99%。
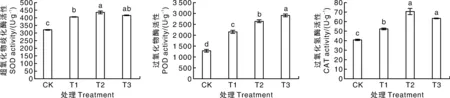
图6 高温胁迫下甜菜碱处理的茄子幼苗抗氧化酶活性Fig.6 Antioxidant enzyme activities of glycine betaine treated eggplant seedlings under high temperature stress
2.8 外源甜菜碱对高温胁迫下茄子MDA、脯氨酸含量和超氧阴离子含量的影响
喷施不同浓度甜菜碱均降低了高温胁迫下茄子幼苗叶片的MDA含量(图7-A)。与CK相比,T1和T2处理的叶片MDA含量显著降低,分别降低22.1%和43.08%;T3处理的叶片MDA含量与CK无显著差异。T1、T2、T3处理的茄子幼苗叶片脯氨酸含量较CK均显著增加(图7-B),分别增加16.92%、58.13%、48.68%。叶片组织化学染色结果显示,与CK相比,T2、T3处理叶片颜色明显变浅,其中,T2处理的叶片染色最浅,说明叶片超氧阴离子含量最低(图7-C)。
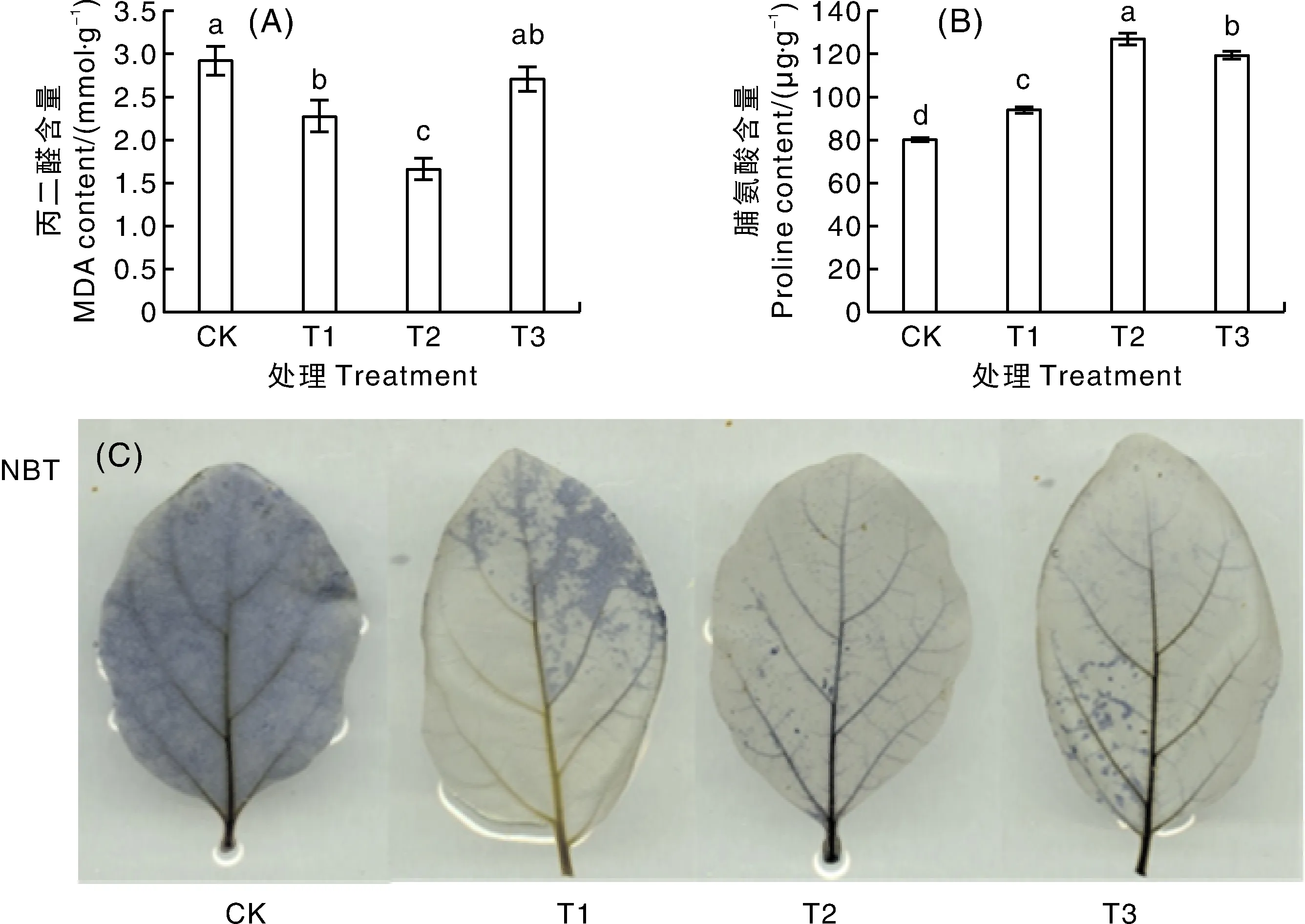
A,丙二醛含量;B,脯氨酸含量;C,茄子叶片组织化学染色结果。A, Results of malondialdehyde (MDA) content; B, Results of proline content; C, Results of histochemical staining of eggplant leaves.图7 高温胁迫下甜菜碱对茄子MDA含量、脯氨酸含量和超氧阴离子的影响Fig.7 Effects of glycine betaine on MDA content, proline content and superoxide anion in eggplant under high temperature stress
3 讨论
高温胁迫对幼苗的生长造成伤害,抑制了茄子幼苗株高、全株鲜重、全株干重、地上部分鲜重、地上部分干重[26]。高温胁迫下喷施不同浓度甜菜碱后,茄子幼苗的株高和生物量明显增加,表明甜菜碱有效缓解了高温对茄子幼苗生长的抑制作用。Tisarum等[27]研究发现,外源施用甜菜碱可维持缺水胁迫下2个品种水稻的总体生长性能和产量性状。Islam等[20]研究结果表明,在正常生长条件下叶面喷施20 mmol·L-1甜菜碱,芥菜植株地上部生物量有一定程度的增加。Zulfiqar等[28]研究表明,甜菜碱可以稳定热胁迫下植物的光合作用,促进热胁迫下植物的生长。本研究结果表明,高温胁迫抑制了茄子幼苗的生长,降低了植株干鲜重;40 mmol·L-1甜菜碱能够显著促进高温胁迫下茄子幼苗株高和生物量的积累,这与Islam等[20]的研究结果一致。根系不仅是植株吸收养分、运输营养物质和水分的主要结构器官,还起到固定植株、合成生命物质的作用。因此,根系的生长状况和结构可以作为反映植物抗性强弱的一个重要生理指标[29]。本研究结果表明,高温胁迫下,外源喷施甜菜碱能使茄子幼苗的根系保持良好的生长状况,40 mmol·L-1外源甜菜碱能够显著促进幼苗根长、根表面积、根体积和根尖数。说明甜菜碱通过促进根系的生长发育,进而促进植株生长,提高生物量的积累。
叶绿素是植物光合作用的主要色素,其含量直接影响植物的正常光合作用,甚至影响植物的正常新陈代谢[30]。高温胁迫会损伤植物叶绿体结构,降解植物体叶绿素含量。赵新西等[31]认为,甜菜碱能保护逆境下的蛋白质和色素,维持正常的光合作用。本研究结果表明,高温胁迫下喷施40 mmol·L-1甜菜碱能够显著减轻茄子幼苗叶片光合色素的降解,这与王继玥等[32]的研究结果一致。
光合作用是植物体重要的生理生化过程,高温胁迫下植物光合作用的下降有气孔因素和非气孔因素,非气孔因素主要是光合相关酶活性下降和光合器官损伤[33]。Mäkelä等[34]研究表明,逆境胁迫下甜菜碱可通过提高CO2的同化率来维持类囊体膜的稳定性;在逆境胁迫下,甜菜碱可通过保护D1蛋白来维持光系统的正常运行。余雪娜等[35]研究表明,喷施低浓度的外源物质后Tr升高,表明植株通过提高蒸腾作用降低叶表温度,保持体内较高的酶活性,从而间接提高光合速率。Gs升高而Ci却降低,表明低浓度的外源物质通过增大Gs来减小气孔因素对光合作用的抑制,同时增强了CO2的固定和转化效率,降低非气孔因素影响。喷施40 mmol·L-1甜菜碱维持了茄子幼苗叶片的正常生长,提高了茄子幼苗光合作用,增强了光合作用相关酶活性。喷施60 mmol·L-1甜菜碱时,茄子叶片中的Ci与对照差异不显著,可能是由于过高浓度的甜菜碱对茄子幼苗叶片有毒害作用,使CO2的固定和转化受到抑制。
植物中CO2固定途径是卡尔文循环,植物的光合作用受卡尔文循环相关酶调节[36]。Rubisco是卡尔文循环的关键酶,是所有光合生物进行光合碳同化的关键性酶;TK在很大程度上控制着光合碳固定和RuBP再生;GAPDH是卡尔文循环中的关键酶,在糖代谢和能量代谢过程中起着重要的作用;FBPase加速卡尔文循环有利于提高植物的光合效率;FBA在叶绿体光合碳化阶段起重要作用[37]。本研究结果表明,甜菜碱提高了高温胁迫下茄子幼苗RuBisco酶活性,促进了CO2的固定,提高了光合作用的碳同化速率。甜菜碱处理显著提高了高温胁迫下茄子幼苗的FBPase、FBA、GAPDH和TK的活性,从而促进了碳代谢途径,提高了光合作用。叶绿素荧光动力学及其参数是利用叶绿素荧光探针快速、灵敏、无害化检测逆境对植物光合作用和生理状态影响的理想方法[38]。有研究认为,高温胁迫下植物的光系统受到破坏,Fv/Fm、ΦPSⅡ、qP会降低,而NPQ会升高[39]。有学者研究认为,在高温光抑制处理下,PSⅡ可能在转基因植物中快速有效地修复,从而减轻PSⅡ的光抑制[40]。在本试验中,高温胁迫下喷施40 mmol·L-1甜菜碱后,茄子幼苗Fv/Fm、ΦPSⅡ、qP升高,NPQ降低,表明甜菜碱可通过抑制PSⅡ潜在活性的下降,提高热耗能力,减轻高温对PSⅡ的伤害程度,使茄子叶片吸收的光能最大程度地用于光合电子传递,提高光合作用效率,最终缓解高温对茄子幼苗生长的抑制。
高温胁迫下茄子幼苗的SOD、POD和CAT活性显著增加,表明它们在高温信号下能够通过增强自身抗氧化酶活性来适应高温[41]。有学者认为,甜菜碱具有很强的清除自由基的能力,可通过诱导抗氧化酶活性、降低氧化胁迫有效缓解胁迫[13]。本研究结果表明,喷施40 mmol·L-1甜菜碱可以增加茄子幼苗抗氧化酶活性,增强高温胁迫下幼苗抗氧化能力和自由基清除能力,进而提高茄子幼苗对高温胁迫的抵御能力,这与呼彧等[14]在高温胁迫下对菜豆幼苗喷施甜菜碱的研究结果一致。
在高温胁迫下,植物器官常发生膜脂过氧化作用,MDA就是产物之一,通常被用来作为细胞膜脂质过氧化的指标,用于表示植物对逆境条件反应的强弱[42]。在抗旱、抗寒等逆境中的研究表明,喷施一定浓度的甜菜碱溶液能够减轻逆境胁迫对番茄[43]、水稻[44]的影响,可减轻逆境胁迫对质膜造成的伤害,抑制MDA积累。本研究结果表明,与对照相比,喷施40 mmol·L-1甜菜碱显著抑制了MDA的积累,减轻了膜脂过氧化程度,这与柴文臣等[45]的研究结果相一致。渗透调节是植株抵抗高温损伤的生理因素之一,脯氨酸是植物体内重要的渗透调节物,在植物受到胁迫时,脯氨酸的积累对防止水分的流失和提高原生胶体稳定性有一定关系,大量的脯氨酸缓解高温加剧蒸腾作用的损害,减少细胞质膜损伤,增强抗逆性[5]。本研究结果表明,高温胁迫下喷施40 mmol·L-1甜菜碱使茄子幼苗叶片的脯氨酸含量增加,表明甜菜碱能增加茄子幼苗的脯氨酸含量,保持渗透平衡,减少细胞质膜损伤,这与刘术均等[46]的研究结果一致。说明在胁迫条件下外源甜菜碱可提高茄子叶片脯氨酸含量,并能保持膜系统和膜结构的稳定性,使植株进行正常的生理代谢功能。
4 结论
外源甜菜碱(40 mmol·L-1)能够提高抗氧化酶活性,降低茄子幼苗MDA含量,进而缓解高温胁迫对茄子幼苗造成的氧化损伤和细胞膜的伤害,提高茄子对高温逆境的耐受能力,促进茄子幼苗在高温胁迫下生物量和叶绿素的积累,提高净光合速率和光系统Ⅱ的实际光化学效率,增强卡尔文循环相关酶活性,进而提高茄子幼苗在高温下的光合作用。
