不同砧木对吐鲁番地区‘克瑞森无核’葡萄生长特性及果实品质的影响
2023-10-10白世践户金鸽吴久赟张雯谢辉赵荣华陈光蔡军社
白世践, 户金鸽, 吴久赟, 张雯, 谢辉, 赵荣华, 陈光, 蔡军社*
(1.新疆维吾尔自治区葡萄瓜果研究所,新疆 吐鲁番 838200; 2.新疆农业科学院吐鲁番农业科学研究所,新疆 吐鲁番 838000; 3.新疆农业科学院园艺作物研究所,乌鲁木齐 830091)
‘克瑞森无核’葡萄是优良无核晚熟葡萄的代表性品种,于1998年从美国引入吐鲁番地区[1],该品种具有无核、大粒、肉脆、糖酸比适中、耐贮运等优点,但该品种在吐鲁番地区特殊生态条件下存在果实着色困难、易萎蔫、自根树生长势不易控制、花芽分化差等问题,导致商品果率及产量偏低[2],严重制约了‘克瑞森无核’葡萄在吐鲁番地区乃至全疆的发展。为解决‘克瑞森无核’葡萄栽培上存在的问题,从植物生长调节剂使用[2-3]、叶面肥喷施[4]、乙醇与氯化钙处理[5]、水肥管理[6]等方面进行了研究,并在改善其栽培性状及果实品质方面取得了一定的效果。砧化栽培即使用抗性砧木进行嫁接栽培,砧化栽培与这些措施相比,具有长效、稳定的优势,优良的砧木不仅能够提高葡萄的生态抗逆性,还可以调控葡萄生长势、提高果实品质及产量。
葡萄砧化栽培最初被用于欧洲葡萄抵抗根瘤蚜危害,随后葡萄砧木与接穗之间的互作影响被广泛研究。葡萄砧木主要包括河岸葡萄、沙地葡萄、冬葡萄等野生葡萄自交选育的砧木品种以及以河岸葡萄×沙地葡萄、冬葡萄×河岸葡萄和冬葡萄×沙地葡萄杂交选育的优良砧木品种。河岸葡萄具有极抗根瘤蚜、易生根、易嫁接,但生长势偏弱的特点;沙地葡萄具有抗根瘤蚜、抗旱、生长势强的特点;冬葡萄具有耐高钙质和碱性土壤、抗根瘤蚜的特点[7]。大量研究证实,砧木不仅能够提高接穗品种对生物胁迫(根瘤蚜和线虫)和非生物胁迫(干旱、低温及盐碱)的抗性[8],还会对接穗品种的生长[9-11]、光合特性[12]、产量[10]和果实品质[13]等栽培性状产生明显影响。Frioni等[8]研究发现,M4砧木在干旱胁迫下能够通过推迟气孔关闭改善气体交换和水分状况,提高‘Grechetto Gentile’葡萄抗旱性。牛锐敏等[9]研究认为,砧木110R 能够明显增强‘霞多丽’葡萄生长势,促进主干粗度、叶面积及叶片SPAD值增加。沈碧薇等[10]研究发现,在长江以南地区采用华佳8 号砧木不仅能够增大‘瑞都红玉’葡萄新梢节间粗,促进新梢生长势,还能够提高单株产量,促进果实着色,进而改善果实品质,而采用抗砧1 号和225Ru 砧木的嫁接树新梢节间显著变短,新梢生长势减弱;南方湿热地区采用Macadams、101-14、Riparia Gloire 和Dogridge砧木能够显著增大‘天工翠玉’葡萄新梢节间粗,增强植株生长势[11]。韩晓等[12]研究表明,3309C、1103P 能够提高设施‘87-1’葡萄表观量子效率、降低光补偿点和暗呼吸速率从而提高其耐弱光能力,SO4、华葡1 号能够提高‘87-1’葡萄耐高温能力。Cheng 等[13]研究认为,101-14 砧木能够增加‘霞多丽’葡萄可滴定酸含量,延迟葡萄成熟,而1103 能够显著增加‘霞多丽’浆果中游离和结合态挥发性化合物总量。此外,砧木接穗栽培特性及品质还受品种特性、地域气候条件等多种因素的影响,亦会表现出不同地域同种砧木对同一接穗品种的影响结果不一致的现象,如高展等[14]研究表明,在玛纳斯河流域,5BB、SO4 砧木能显著提高‘马瑟兰’葡萄叶片的光合指标;而李敏敏等[15]研究发现,SO4 对河北昌黎产区‘马瑟兰’葡萄叶片光合速率无明显影响,5BB 砧木则会降低‘马瑟兰’葡萄叶片净光合速率。
不同产区生态条件不同,葡萄品种存在的问题及砧木适应性不尽相同,砧木对接穗栽培性状的影响亦不尽相同。研究砧木对‘克瑞森无核’葡萄生长特性及果实品质的影响对解决该品种在吐鲁番地区生态条件下生长势过强、产量低及果实着色困难、易萎蔫等栽培问题具有重要意义。为此,本研究以新疆吐鲁番产区表现较好的7 个砧木与‘克瑞森无核’葡萄进行嫁接组成砧穗组合,以‘克瑞森无核’葡萄自根树为对照,分析不同砧穗组合及自根树在生长势、光合特性、产量及果实品质等方面的差异,旨在筛选对本产区‘克瑞森无核’葡萄生长势调控效果好,并能够显著改善其丰产性及果实品质的优良砧木,促进本产区葡萄产业健康发展。
1 材料与方法
1.1 试验地概况
试验于2020 年在新疆维吾尔自治区葡萄瓜果研究所葡萄栽培示范基地(42°91′N,90°30′E)进行。试验地海拔419 m,年降雨量25.3 mm,年蒸发量2 751 mm,全年日照时数3 122.8 h,10 ℃以上有效积温4 525 ℃以上,无霜期192 d,属于典型的大陆性暖温带荒漠气候,为极端干旱区气候类型。土壤质地为碱性砾石沙壤土,pH 8.0左右。
1.2 试验材料与设计
供试葡萄品种为欧亚种‘克瑞森无核’(Vitis viniferaL. cv.Crimson Seedless, CS),嫁接树砧木分别为5BB、SO4、3309M、110R、山河3 号(Shanhe 3)、河岸7 号(Hean 7)、贝达(Beta),以自根树(CS rooting)为对照,砧木品种和‘克瑞森无核’葡萄均由新疆维吾尔自治区葡萄瓜果研究所葡萄资源圃提供。砧木和‘克瑞森无核’葡萄自根树均于2015年定植,嫁接树于2017年采用绿枝高接法进行嫁接,树龄均为6 年生。采用顺行龙干+(V+水平)叶幕架式栽培,东西行向,株行距2.0 m×3.5 m。每连续10 株(5 档)为1 小区,3 次重复,随机区组排列。葡萄生长季不进行摘心处理,田间土肥水管理和病虫害防治管理同当地常规。
1.3 测定指标与方法
1.3.1葡萄叶片质量、光合参数及叶绿素荧光参数测定 葡萄叶片光合参数及叶绿素荧光参数测定于7月中旬晴天11∶00进行,以健壮结果母枝基部以上第5~6 片健康代表性功能叶为测定对象。利用Li-6400 型光合仪(Li-Cor Inc.,Lincoln,NE,USA)进行光响应曲线的测定。光合有效辐射按照2 500、2 100、1 700、1 300、900、600、300、200、100、40、15、0 µmol·m-2·s-1的顺序设定,CO2浓度设定为400 µmol·mol-1,温度为25 ℃,气体流速为500 mmol·s-1。利用直角双曲线修正模型求得光补偿点(light compensation point, LCP)、暗呼吸速率(dark respiration rate,Rd)、表观量子效率(apparent quantum yield, AQY)、最大净光合速率(maximum net photosynthetic rate,Pnmax)指标[16]。
叶绿素荧光参数采用FMS-2 型荧光仪(Hansatech Instruments LTD., Norfolk,UK)在树体直接测定。将选好的叶片经过充足自然光适应,测定自然光下叶片叶绿素荧光参数,得出光系统Ⅱ(photosystem Ⅱ complex, PS Ⅱ)电子传递速率(electron transport rate, ETR)和实际光化学效率(actual photochemical efficiency,ΦPSⅡ)。叶片暗适应30 min 后,测定暗适应下叶片初始荧光值(original fluorescence,Fo)、最大荧光值(minimal fluorescence,Fm)、PSⅡ最大光能转化效率(PSⅡmaximum light energy conversion efficiency,Fv/Fm)、PSⅡ潜在光化学效率(PSⅡPotential photochemical efficiency,Fv/Fo)等荧光参数,计算非光化学猝灭系数(non-photochemical quenching, NPQ)。
单叶质量采用称量法测量,叶绿素质量分数采用乙醇浸提法[17]测定。
1.3.2植株生长势测定 8 月初统计每株新梢叶片数量,用卷尺测量新梢长度,采用一刀切的方式剪下所有新梢超过1.5 m 的部分并称量作为夏季生长量。秋季修剪后用游标卡尺测量嫁接口上5 cm处接穗径粗和嫁接口下5 cm 处砧木径粗;测定1 年生枝条第2 节粗度,每小区随机测定60 个数据。
1.3.3果实品质及产量测定 葡萄采收期(9 月28 日)实测每小区产量,计算平均单株产量。每小区随机采10 个代表性果穗,参照OIV(International Organisation of Vine and Wine)描述[18],对果穗紧密度进行赋值统计,并对口感中的肉质、甜味程度进行描述评价。常规方法测量果穗质量、果粒质量和果粒纵径、横径;采用PAL-1型手持数显折射仪(Atago, Tokyo, Japan)测定可溶性固形物(total soluble solid, TSS)含量;采用CR-400 手持色差计(Konica Minolta, Tokyo, Japan)测定果粒赤道部位的色泽指标L、a、b值,计算出红色葡萄果实色泽指数(color index of red grape,CIRG)[19]。可滴定酸(titratable acidity, TA)含量采用NaOH 滴定法[20]测定;维生素C(Vc)含量采用钼蓝比色法[21]测定;果皮中总类黄酮含量采用NaNO2-Al(NO3)3比色法[22]测定;总花色苷含量采用pH 示差法[23]测定,以花青素-3-葡萄糖苷(cyanidin-3-glucoside equivalent, CGE, mg·g-1)表示。固酸比(TSS/TA)为可溶性固形物含量与可滴定酸含量的比值。
1.4 数据处理
采用SPSS 17.0和Microsoft Excel 2010软件进行数据统计与分析,试验数据以平均值±标准差表示,Duncan’s法进行多重比较(P<0.05),主成分分析参考周广生等[24]的方法。
对指标值进行均衡归一化处理,即将数据转变为纯量,公式如下。
式中,Wij和W′ij分别为第j品种第i指标正、负指标转化值;aij指第j个品种第i个指标值;aminij、amaxij分别指第j品种第i组指标的最小和最大值。
2 结果与分析
2.1 不同砧木下‘克瑞森无核’葡萄叶片性状分析
由表1 可知,砧木嫁接均显著减小了‘克瑞森无核’葡萄单叶质量,其中以CS/5BB 和CS/SO4 单叶质量最小,均显著小于其他砧穗组合。CS/5BB和CS/110R 叶绿素a 含量、叶绿素b 含量、类胡萝卜素含量和总叶绿素含量较自根树均显著增大,其中,叶绿素a 含量增幅分别为23.65% 和11.49%,叶绿素b 含量增幅分别为36.54% 和15.35%,类胡萝卜素含量增幅分别为28.57%和20.00%,总叶绿素含量增幅分别为26.50% 和12.50%;CS/SO4 和CS/河岸7 号叶绿素a 含量、叶绿素b 含量、类胡萝卜素含量和总叶绿素含量较自根树均显著降低,其中,叶绿素a 含量降幅分别为24.32%和11.49%,叶绿素b 含量降幅分别为30.77% 和15.38%,类胡萝卜素含量降幅均为17.14%,总叶绿素含量降幅分别为26.00% 和14.50%。与自根树相比,CS/SO4 和CS/河岸7 号的叶绿素a/b 值显著增大,增幅分别为12.01%和13.78%;而CS/5BB叶绿素a/b值显著降低,降幅为8.83%。CS/3309M、CS/山河3 号、CS/贝达与自根树的叶绿素相关指标均未表现出显著差异。结果表明,砧木嫁接均减小了‘克瑞森无核’葡萄单叶质量,但不同砧木对葡萄叶片叶绿素组成影响不同,5BB 和110R 提高了‘克瑞森无核’葡萄叶片的叶绿素含量,而SO4 和河岸7 号降低了‘克瑞森无核’葡萄叶片叶绿素含量。

表1 不同砧木下‘克瑞森无核’葡萄叶片性状Table 1 Leaf characters of ‘Creson seedless’ grape under different rootstocks
2.2 不同砧木下‘ 克瑞森无核’葡萄叶片光合参数分析
由表2 可知,表观量子效率(AQY)以CS/山河3 号最高,显著高于自根树20%;CS/SO4、CS/3309M、CS/110R 和CS/5BB 均较自根树显著降低,降幅分别为53.55%、60.00%、33.33% 和20.00%;而CS/河岸7 号和CS/贝达与自根树无显著差异。最大净光合速率(Pnmax)以CS/山河3 号和CS/贝达最高,显著高于自根树,增幅均为15.65%;CS/SO4、CS/3309M、CS/110R 和CS/河岸7 号均显著低于自根树,降幅分别为52.25%、33.67%、35.53%和24.73%;CS/5BB 和自根树无显著差异。CS/山河3 号光补偿点(LCP)显著低于自根树,降幅为23.48%;CS/SO4、CS/3309M、CS/110R 和CS/5BB 光补偿点相比自根树均显著增高,增幅分别为146.13%、129.01%、40.54% 和33.21%;CS/河岸7 号、CS/贝达与自根树无显著差异。相比自根树,CS/5BB 暗呼吸速率(Rd)显著提高,增幅13.77%,其他砧穗组合与自根树无显著差异。可见,山河3 号和贝达提高了‘克瑞森无核’葡萄叶片的光合能力和耐弱光能力,而SO4、3309M 和110R 降低了‘克瑞森无核’葡萄光合能力和耐弱光能力。

表2 不同砧木下‘克瑞森无核’葡萄的叶片光合参数Table 2 Photosynthetic parameters of ‘Creson seedless’ grape leaves under different rootstocks
2.3 不同砧木下‘克瑞森无核’葡萄叶绿素荧光参数分析
由表3 可知,CS/贝达、CS/5BB 和CS/SO4 的初始荧光值(Fo)显著大于自根树,增幅分别为28.03%、18.81%和8.50%;CS/河岸7 号则显著小于自根树,降幅为8.86%。CS/SO4、CS/5BB、CS/山河3 号和CS/贝达最大荧光值(Fm)相比自根树显著提高,其他砧穗组合与自根树差异均不显著。CS/5BB、CS/SO4、CS/山河3 号、CS/3309M、CS/110R相比自根树PSⅡ潜在光化学效率(Fv/Fo)、PSⅡ最大光化学效率(Fv/Fm)均显著提高,CS/贝达相比自根树显著提高了PSⅡ最大光化学效率(Fv/Fm),而CS/河岸7号PSⅡ潜在光化学效率(Fv/Fo)相比自根树显著降低,降幅为12.71%。CS/贝达和CS/山河3 号实际化学效率(ΦPSⅡ)相比自根树显著增大,增幅分别为16.67%和5.56%,其他砧穗组合与自根树无显著差异。CS/山河3 号、CS/SO4 和CS/贝达相比自根树非光化学猝灭系数(NPQ)显著降低,降幅分别为15.63%、26.56%和53.13%。CS/贝达和CS/SO4相比自根树PSⅡ电子传递速率(ETR)显著增大,增幅分别为19.25%和13.35%,其他砧穗组合与自根树差异均未达显著水平。可见,贝达和山河3 号提高了‘克瑞森无核’葡萄光能利用潜力和光能转化效率,而河岸7 号降低了‘克瑞森无核’葡萄叶绿素PSⅡ的潜在光化学活性。

表3 不同砧木下‘克瑞森无核’葡萄的叶绿素荧光参数Table 3 Chlorophyll fluorescence parameters of ′Creson seedless′ grape under different rootstocks
2.4 不同砧木下‘ 克瑞森无核’葡萄的生长势分析
由表4 可知,砧木嫁接均显著减小了‘克瑞森无核’葡萄新梢长度、夏季生长量和新梢粗度,新梢长度以CS/SO4 减幅最大,达54.60%,CS/5BB、CS/3309M 和CS/河岸7 号减幅较大,在36.91%~31.27%;夏季生长量同样以CS/SO4 减幅最大,达99.57%,其余砧穗组合减幅在71.00%~89.59%;7个砧穗组合新梢粗度减幅在14.16%~32.75%,以CS/河岸7 号减幅最大;CS/SO4 和CS/3309M 相比自根树还显著减小了新梢叶片数,减幅分别为19.50%和22.64%。CS/110R 相比自根树接穗径粗显著减小,减幅22.48%;砧木径粗以CS/110R最小,CS/SO4 和CS/贝达较小;CS/山河3 号、CS/贝达、CS/SO4 和CS/3309M 穗砧粗度比大于1.40,出现较明显的小脚现象,CS/河岸7 号小脚现象最不明显。结果表明,砧木嫁接均会抑制‘克瑞森无核’葡萄生长势,SO4嫁接的‘克瑞森无核’葡萄的生长势最弱,而河岸7 号嫁接的‘克瑞森无核’葡萄长势中庸且小脚现象不明显。
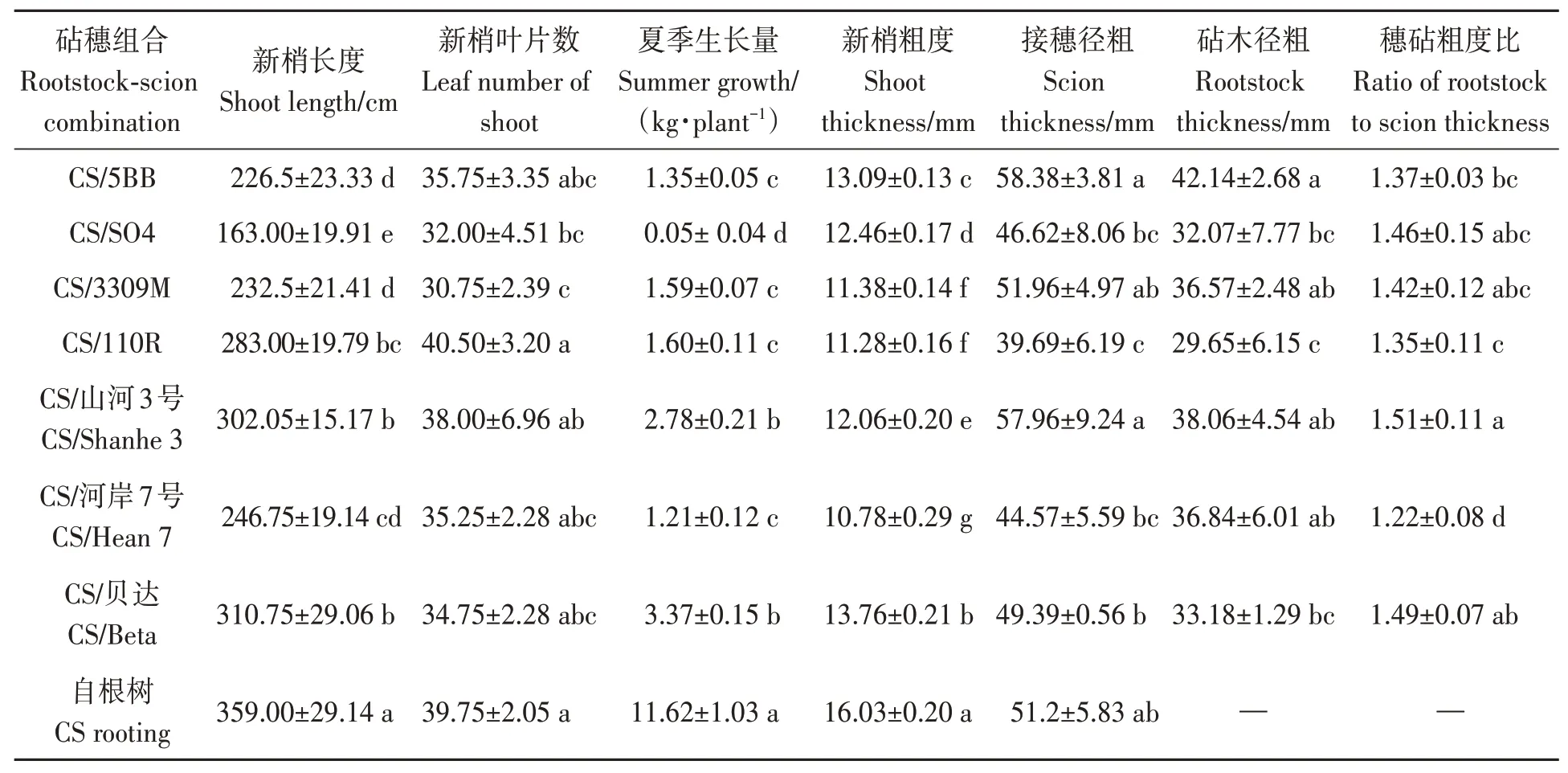
表4 不同砧木下‘克瑞森无核’葡萄生长势Table 4 Growth potential of ‘Creson seedless’ grape under different rootstocks
2.5 不同砧木下‘克瑞森无核’葡萄果实品质及产量分析
2.5.1不同砧木下‘克瑞森无核’葡萄外观品质及产量分析 由表5 可知,砧木嫁接均显著增大了‘克瑞森无核’葡萄果穗质量,其中以CS/贝达增幅最大,为476.25%,CS/山河3 号增幅较大,为310.85%,CS/110R 增幅最小,为135.14%;除CS/110R外,其余6个砧穗组合相比自根树单株产量均显著增大,以CS/山河3 号、CS/河岸7 号和CS/贝达增幅最大,分别为618.49%、587.39%和534.45%。CS/贝达、CS/SO4、CS/河岸7 号、CS/山河3 号和CS/5BB 砧穗组合相比自根树果粒质量均显著增大,增幅为17.58%~69.92%;相比自根树,CS/SO4、CS/河岸7 号和CS/贝达纵径、横径均显著增大;CS/SO4、CS/5BB 和CS/3309M 相比果形指数自根树均显著增大。除CS/110R 外,其余6 个砧穗组合果穗紧密度均显著大于自根树,果穗紧密度由疏松变为适中或偏紧;CS/3309M 和CS/110R 萎蔫率相比自根树均显著升高,增幅分别为和34.62%和50.79%,其余5 个砧穗组合相比自根树萎蔫率均显著降低;除CS/110R 外,其余6 个砧穗组合相比自根树均显著降低了葡萄色泽指数CIRG 值,CS/3309M 的CIRG 值较高,其他砧穗组合均较低,尤其是CS/贝达存在严重着色不良现象。结果表明,砧木嫁接抑制‘克瑞森无核’葡萄营养生长的同时促进生殖生长,但过度的抑制营养生长并不利于产量及果实品质的形成;虽然产量低、果粒质量小的砧穗组合葡萄颜色指数CIRG 值较高,着色较好,但更易产生萎蔫,而中等的树势及产量更易获得好的外观品质。

表5 不同砧木下‘克瑞森无核’葡萄的外观品质和产量Table 5 Appearance quality and yield of ‘Creson seedless’ grape under different rootstocks
2.5.2不同砧木下‘克瑞森无核’葡萄内在品质的分析 由表6可知,除CS/3309M和CS/110R外,其余5 个砧穗组合相比自根树均显著降低了可溶性固形物含量,以CS/5BB 和CS/SO4 降幅最大,为20.04%和20.34%;除CS/山河3 号和CS/5BB 外,其他5 个砧穗组合相比自根树均显著降低了可滴定酸含量,以CS/SO4 和CS/110R 降幅最大,分别为25.35%和18.31%;相比自根树,CS/110R 固酸比值显著增大,增幅达24.61%,CS/山河3 号和CS/5BB 固酸比值相比自根树显著降低,降幅分别为14.70%和18.63%,其他砧穗组合与自根树无显著差异。砧木嫁接的‘克瑞森无核’葡萄维生素C 含量相比自根树均显著降低,降幅在3.17%~19.51%;CS/贝达总类黄酮含量相比自根树显著增高,增幅6.60%,CS/山河3 号与自根树无显著差异,其他砧穗组合相比自根树均显著降低,降幅4.74%~36.92%;砧木嫁接的‘克瑞森无核’葡萄总花色苷含量相比自根树均显著减小,CS/110R 和CS/3309M 降幅较小,分别为25.33%和64.00%,其他砧穗组合降幅均超过77.33%。CS/3309M、CS/110R 和自根树因萎蔫率较高,葡萄果肉质地软,CS/3309M 和CS/110R 因固酸比值高,口感表现为甜,CS/5BB 和CS/山河3 号固酸比值低,表现为酸甜,其他砧穗组合表现为酸甜适中。可见,砧木嫁接均会对‘克瑞森无核’葡萄着色产生不利影响,河岸7 号、3309M 和110R 嫁接的‘克瑞森无核’葡萄表现出较高的内在品质。

表6 不同砧木下‘克瑞森无核’葡萄的内在品质Table 6 Internal quality of ‘Creson seedless’ grape under different rootstocks
2.6 不同砧穗组合果实品质及产量性状的主成分分析和综合评价
选取与‘克瑞森无核’葡萄果实品质及产量密切相关的果穗质量、果粒质量、果穗紧密度、萎蔫率、单株产量、可溶性固形物(TSS)、可滴定酸(TA)、固酸比(TSS/TA)、维生素C(Vc)、总类黄酮、CIRG、总花色苷共12 项指标进行主成分分析,其中萎蔫率和TA为负向指标,根据公式(2)转化,其余指标根据公式(1)转化。经主成分分析后得到3 个主成分,累计贡献率达到89.850%(表7),说明3 个主成分能够全面反映8 个砧穗组合12项指标信息。
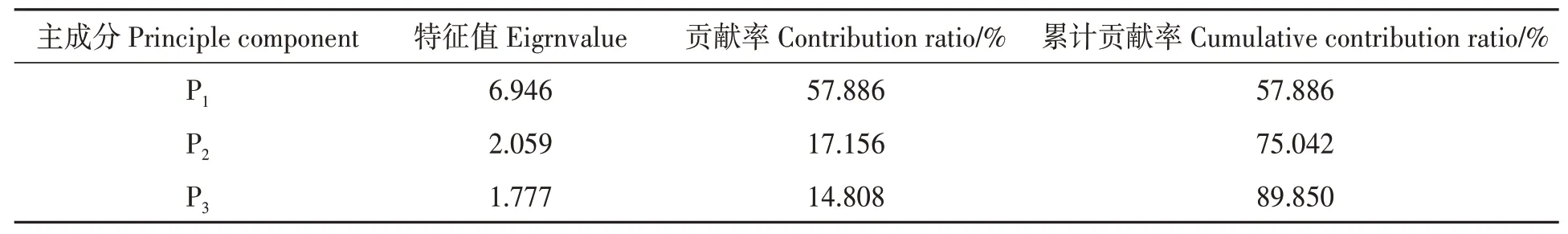
表7 主成分的特征值、贡献率及累计贡献率Table 7 Characteristics value, contribution ratio and accumulated variance of principal components
由表8可知,第1主成分(P1)中载荷值较大且为正值的指标有果穗质量、果粒质量、果穗紧密度、萎蔫率和单株产量,主要反映葡萄丰产性及外观品质信息;第2 主成分(P2)中载荷值较大且为正值的指标有可滴定酸和固酸比,主要反映葡萄口感品质信息;第3 主成分(P3)中载荷值较大且为正值的指标有可溶性固形物和总类黄酮,主要反映葡萄的营养品质信息。

表8 成分距阵Table 8 Component matrix
以标准化后的指标值乘以各主成分特征向量得出各砧穗组合的综合指标值,以各主成分特征值占所提取主成分特征值之和的比例作为权重,建立主成分综合模型:F=0.644F1+0.191F2+0.165F3。根据主成分综合模型计算综合主成分得分F(表9),各砧穗组合综合得分排名为:CS/贝达>CS/SO4>CS/河岸7 号>CS/山河3 号>CS/5BB>CS/3309M>CS/110R>自根树。可见,贝达、SO4和河岸7号嫁接的‘克瑞森无核’葡萄在丰产性、品质方面综合表现较好。结合3 个砧木对‘克瑞森无核’葡萄植株生长的影响结果进行综合评价,贝达嫁接的‘克瑞森无核’葡萄生长势偏强,新梢粗度均超过12.0 mm,有明显的徒长趋势,且出现较为明显的小脚现象(穗砧粗度比为1.49);SO4 嫁接的‘克瑞森无核’葡萄过度削弱生长势,并不利于产量的获得;河岸7 号嫁接的‘克瑞森无核’葡萄生长势中庸,新梢粗度适中,无明显小脚现象,且可溶性固形物含量高,固酸比值较大。综上,吐鲁番地区‘克瑞森无核’葡萄嫁接砧木优选河岸7号。

表9 综合主成分值得分Table 9 Comprehensive principal component score
3 讨论
3.1 不同砧木对‘克瑞森无核’葡萄叶片光合特性及叶绿素荧光特性的影响
叶绿素含量与植物对光能的捕获和积累密切相关,反映植物光合作用能力的强弱。本研究中,5BB显著提高了‘克瑞森无核’葡萄叶片叶绿素含量,山河3 号、贝达和自根树同样具有较高的叶绿素含量,最大净光合速率较大,光补偿点较低,表现出了较强的光合能力和耐弱光能力;而SO4 和河岸7 号则显著降低了‘克瑞森无核’葡萄叶绿素含量,增大了叶绿素a/b 值,SO4嫁接的‘克瑞森无核’葡萄光合能力最弱,且光补偿点最高,耐弱光能力最差,3309M 嫁接的‘克瑞森无核’葡萄同样也表现出了较弱的耐弱光能力,而郝燕等[25]研究认为SO4 提高了河西走廊‘贵人香’葡萄光合能力,韩晓等[12]研究认为3309 提高了‘87-1’葡萄耐弱光能力,与本研究结论不一致。叶绿素荧光可以作为光合作用的有效探针,PSⅡ最大光能转化效率(Fv/Fm)反映植物潜在的最大光合能力,即光能利用潜力,PSⅡ潜在光化学效率(Fv/Fo)代表PSⅡ的潜在光化学活性,反映原初光能转化效率及PSⅡ潜在量子效率[26]。本研究中除河岸7 号外,其他砧木均显著提高了‘克瑞森无核’葡萄PSⅡ最大光能转化效率(Fv/Fm),表现出较强的光能利用潜力,5BB、SO4、山河3 号、3309M 和110R 还显著增强了PSⅡ的潜在光化学活性;贝达嫁接的‘克瑞森无核’葡萄初始荧光值(Fo)较高,说明其对环境变化敏感,叶片类囊体膜更易受损,但贝达提高了‘克瑞森无核’葡萄PSⅡ电子传递速率(ETR),具有较强的光能转化效率。本研究中不同砧穗组合间叶绿素荧光参数表现出了明显差异,而韩晓等[12]研究认为,贝达、SO4、3309嫁接的‘87-1’葡萄间PSⅡ潜在光化学效率(Fv/Fo)和PSⅡ最大光能转化效率(Fv/Fm)无显著差异。可见,砧木对不同生态条件下不同接穗品种的光合特性和荧光参数影响存在明显差异。
3.2 不同砧木对‘克瑞森无核’葡萄生长结果习性的影响
葡萄营养生长与生殖生长为相互依存、相互制约的对立统一关系。营养生长为生殖生长提供物质和能量基础,生殖生长过程产生的激素类物质能够促进物质代谢和转运,有利于营养生长。营养生长过剩容易造成新梢生长与果实发育矛盾, 导致落果或花芽发育不良而难座果、产量降低[27]。本研究中,7 个砧木嫁接均显著降低‘克瑞森无核’葡萄生长势、增大果穗质量,5BB、SO4、山河3 号、3309M、河岸7 号和贝达还显著增大单株产量,这一研究结果与钟海霞等[28]研究认为5BB、SO4、贝达能显著增大天山北麓地区‘克瑞森无核’葡萄产量的结果一致,而与110R、SO4、5BB 利于增强‘克瑞森无核’葡萄生长势的结论不一致,可见,砧木对葡萄生长习性的影响效果还受栽培地域的环境、土壤等多种因素调控。自根树过度的徒长势必造成大量落花落果,产量降低,砧木的调控作用促进营养生长向生殖生长转变,增大果穗质量及产量,SO4 嫁接的葡萄产量低也说明过度的削弱生长势并不利于产量的获得。
3.3 不同砧木对‘克瑞森无核’葡萄品质的影响
砧木通过影响接穗品种生长势、成熟期及果穗微气候,导致果实品质发生改变。本研究中,5BB、SO4、山河3 号、河岸7 号和贝达均显著降低‘克瑞森无核’葡萄可溶性固形物含量,SO4、3309M、110R、河岸7 号和贝达均显著降低可滴定酸含量,这一研究结果与钟海霞等[28]研究认为5BB 和SO4 能显著提高天山北麓产区‘克瑞森无核’葡萄可溶性固形物含量的结果不一致,而与SO4降低可滴定酸含量的研究结果较一致。110R和3309M 降低了‘克瑞森无核’葡萄可滴定酸含量,提高了固酸比值,促进葡萄早熟,这与孙磊等[29]在瑞都香玉品种上的研究结论一致。贝达虽然降低了‘克瑞森无核’葡萄可滴定酸含量,但总花色苷含量及固酸比值依然较低,表现出延迟成熟的特性,这与前人在‘贵人香’[25]、‘克瑞森无核’[28]葡萄上的研究结果一致。值得关注的是,吐鲁番产区‘克瑞森无核’葡萄着色不良和成熟葡萄易萎蔫的问题依然存在,如何通过栽培手段调控营养生长和生殖生长平衡及果际微气候来解决葡萄着色及成熟葡萄抗萎蔫问题依然是产区‘克瑞森无核’葡萄优质栽培的重点工作。
综上所述,7 个砧木嫁接的‘克瑞森无核’葡萄及自根树在生长、光合特性及果实品质方面存在明显差异。砧木嫁接均会抑制‘克瑞森无核’葡萄生长势,增大果穗质量,降低葡萄CIRG 值和总花色苷含量;5BB、SO4、3309M、山河3 号、河岸7号和贝达嫁接的‘克瑞森无核’单株产量显著增大。综合分析认为,吐鲁番地区‘克瑞森无核’葡萄优选砧木为河岸7号。
