多年鼠类防控对高原鼠兔种群数量及植物群落结构的影响
2023-10-08宋文杰张海兰洛藏昂毛卡着才让艾得协措向志强李文金
宋文杰, 张海兰, 洛藏昂毛, 卡着才让, 艾得协措, 周 喜, 向志强, 喻 望, 李文金*
(1.兰州大学生态学院, 甘肃 兰州 730000; 2.河南蒙古族自治县草原工作站, 青海 黄南州 811599)
自上个世纪八十年代以来,由于草地的不合理利用以及气候变暖,青藏高原的草地退化现象愈发严重,同时也伴随着小型啮齿类动物种群爆发,进而发生鼠害[1,2]。害鼠种类主要有高原鼠兔(Ochotonacurzoniae)和高原鼢鼠(Eospalaxfontanierii)等。据统计,高原鼠兔对草地的危害占全国鼠害面积的35%~45%[1]。高原鼠兔(Ochotonacurzoniae),隶属于兔形目、鼠兔科、鼠兔属,主要分布于西藏、青海、甘肃南部、四川西北部,是该区域的小型优势哺乳动物[3]。我国每年投入大量的人力物力财力进行鼠类防控,然而鼠害依旧严重。调查显示,近年来青海省草地鼠害年均危害面积约826.67万公顷,严重危害面积年均720万公顷[4],因此,鼠类防控任务仍然艰巨而繁重。
近年来高原鼠兔对草地植物群落结构和土壤等成分的影响已有大量研究[5-10],高原鼠兔对植物和土壤的影响有很大不确定性,有积极的作用,也有消极的影响[5-10],很大程度上取决于干扰的强度[6,7,10]。然而,这些研究主要基于空间上鼠类密度差异的观测,不能代表真正意义上的鼠类种群密度。此外,尽管我国草地开展了多年的鼠类防控项目,但目前为止,仍不清楚鼠类防控对鼠类种群和植物的影响如何?鼠类防控后鼠兔种群动态的理论研究表明,当灭鼠频率为每年1次,灭杀率小于70%时,鼠兔种群一年后便恢复到原来的状态;当保持灭鼠频率和持续时间不变,灭杀率为90%时,恢复到鼠兔原种群大小只需两年[11]。因此,低频次短时间灭鼠并不能有效控制鼠兔种群数量,无法降低鼠害。但是当灭鼠频次不变,灭鼠率在80%以上,而且灭鼠活动持续3~5年,理论上便可有效控制鼠害[11]。因此,鼠类防控对鼠兔种群数量影响很大程度上取决于灭鼠时间、灭鼠频率以及灭鼠率。
鼠兔种群密度能改变植物群落和土壤的结构和功能。观测啮齿动物种群密度与植被格局的研究发现,啮齿动物种群高密度地区的植被盖度、地上生物和高度均低于低密度区域[2]。鼠类对草地表层土壤有显著影响,坍塌的鼠兔洞口会形成浅坑,在风蚀作用和鼠兔挖掘洞穴行为的共同影响下互相连接,形成草地斑秃,最终导致草地变成黑土滩[12]。通过网围栏去除高原鼠兔发现,高原鼠兔的去除显著地增加了植物群落平均高度和禾草的盖度,而降低了豆科植物和杂类草盖度[13]。在青海果洛地区的观测表明,害鼠会通过啃食破坏植物根系使其向土壤表层聚集,并导致莎草类和禾草类植物的生物量急剧减少,而不是主要食物的杂草类植物生物量趋于增加[14]。近年来,青藏高原地区高寒草甸的鼠害问题逐渐成为研究热点[15],然而目前为止,很少有研究探究多年鼠类防控对鼠兔种群数量以及植物群落结构的影响。本次研究基于2013—2021年连续九年,每年春夏秋三次灭鼠的监测数据,研究鼠类防控持续时间以及防控月份对鼠兔种群数量和植物群落结构的影响,分析鼠兔种群数量和植物群落结构的关系,从而为高寒草地有效的鼠类防控提供理论指导。
1 材料与方法
1.1 研究区概况
研究区域位于青海省河南县南旗村周边地区,地理坐标为北纬34°41′10″~34°54′30″,东经101°23′00″~101°33′40″,平均海拔约为3 520 m。该地区属于高原大陆性气候带,处于高原亚寒带湿润气候区。高原鼠兔活动期间(4—10月),月平均降水量为77.2 mm,平均气温为6.8℃。土壤类型为高山草甸土,主要质地为轻壤土和中壤土。在该区域内,主要植物为黑褐苔草(Carexatrofusca)、矮生嵩草(Kobresiahumilis)、美丽风毛菊(Saussureapulchra)、箭叶橐吾(Ligulariasagitta)等。
1.2 试验设计
1.2.1样地设置 在河南县南旗村选择了高原鼠兔洞口数大于1 600个·ha-1的高寒草甸作为固定的试验区域。每年设置了4个50 m×50 m的样方,随机选择其中1个样方为捕杀样方,对其中的高原鼠兔进行完全捕杀,剩余三个样方为密度调查样方,进行高原鼠兔数量统计。
1.2.2灭鼠与监测时间 由于高原鼠兔的繁殖具有季节性,主要集中在每年的4月至8月,尤其在4月下旬到5月上旬之间其繁殖率达到高峰[16]。因此本次试验选择在高原鼠兔种群大量繁殖开始时(4月下旬)进行第1次灭鼠,繁殖期结束时(8月中旬)进行第2次灭鼠。同时也选择在高原鼠兔越冬储粮活动高峰时(10月下旬)进行第3次灭鼠。
1.2.3监测内容与方式 植物群落结构:每年第二次调查时(8月中旬)在3个密度监测样方内分别设置3个1 m2的样方,记录样方内主要物种、植被盖度以及平均高度,并采集植物的地上部分,烘干后称重得到地上总生物量。
高原鼠兔:灭杀采用定面积捕尽法:在捕尽样方内所有的洞口上放置捕鼠器(超过样地边界的洞口也布置了捕鼠器),每天检查五次捕鼠情况(上午9:30,11:00,下午13:00,15:00,17:00各检查一次),检查时将捕获的高原鼠兔及时收集起来,并重新放置捕鼠器,连续捕捉3天。按照捕杀鼠数/有效洞口数公式计算洞口系数。由于高原鼠兔警惕性强,在捕食过程中会有多次警戒行为[17],捕捉难度较大,因此数量统计采用堵洞开洞法:在监测样方内堵塞所有鼠洞洞口,并记录总洞口数,经过24小时后再查看被鼠打开的有效洞口数。洞口系数×有效洞口数便可得到种群密度[18]。
降雨量与温度:通过研究区域附近设置的野外气象监测装置监测了2013—2020年每个月的降雨量与平均气温。
1.3 数据处理方式
用R软件的基础包进行数据分析;segmented包进行分段式线性回归分析。为了弄清气候、鼠兔种群大小与鼠类防控对地上生物量的影响,把每年生长季(5—8月)的平均气温和总降雨量视为气候因素,将灭鼠的年份与月份视为鼠类防控因素,进行多元线性回归分析。通过R语言中的glmm.hp包,采用“平均共享方差”算法分解边际方差,将每个变量的部分方差与平均共享方差结合,得到各变量及其间的相互重叠对因变量的贡献效应[19]。同样的方法分析了气候(全年总降雨量和全年平均温度)与鼠类防控持续时间对鼠兔种群大小的影响效应。利用ggplot2,tidyverse以及VennDiagram包进行数据统计分析与可视化。
2 结果与分析
2.1 多年鼠类防控和防控月份对高原鼠兔种群动态的影响
随着鼠类防控持续时间的增加,高原鼠兔种群数量总体呈现下降趋势(图1)。相对于4月份防控,8月和10月的防控显著地降低了鼠兔种群密度。分段式线性回归分析表明:在2016年10月之前,防控后鼠兔种群数量显著下降,之后缓慢下降。这说明防控前4年(2013/04—2016/10),鼠兔种群急剧下降,之后种群数量保持稳定(2017/08—2021/10)。

图1 连续多年鼠类防控过程中鼠兔种群大小随时间变化
2.2 多年鼠类防控对高寒草甸植物群落结构的影响
多年鼠类防控可影响高寒草甸植物群落结构,其中,植被高度、盖度以及地上生物量随鼠类防控时间的增加而显著增加(图2)。2016年之后的植被盖度和地上生物量比前4年有较大提升,例如,地上生物量约400 g·m-2增长到了将近850 g·m-2,平均盖度从约70%提升到90%。

图2 三个植物群落指标随时间的变化
2.3 鼠兔种群数量与植物群落结构之间的关系
植被高度、盖度以及地上生物量均随着鼠兔种群数量的增加而减小(图3),其中地上生物量的R2=0.279大于其他两个指标的相关系数,这说明地上生物量和鼠兔种群数量之间的负相关关系最为密切,也可以说鼠兔种群数量对地上生物量的变化具有较大的影响。

图3 三个植物群落指标
2.4 降水、气温、鼠类防控和高原鼠兔种群密度对植物群落地上生物量影响的相对贡献
生长季总降雨量(每年5—8月份降雨量总和)对地上生物量的影响最强,高达0.248 2,占总体效应的38.17%;鼠类防控持续时间对生物量的个体效应为0.207 7,占总体效应的31.94%;鼠兔种群大小产生的个体效应为0.110 5,占比16.99%;而生长季平均温度对生物量的影响最小,仅为0.083 9,占总体效应的12.9%(图4A)。
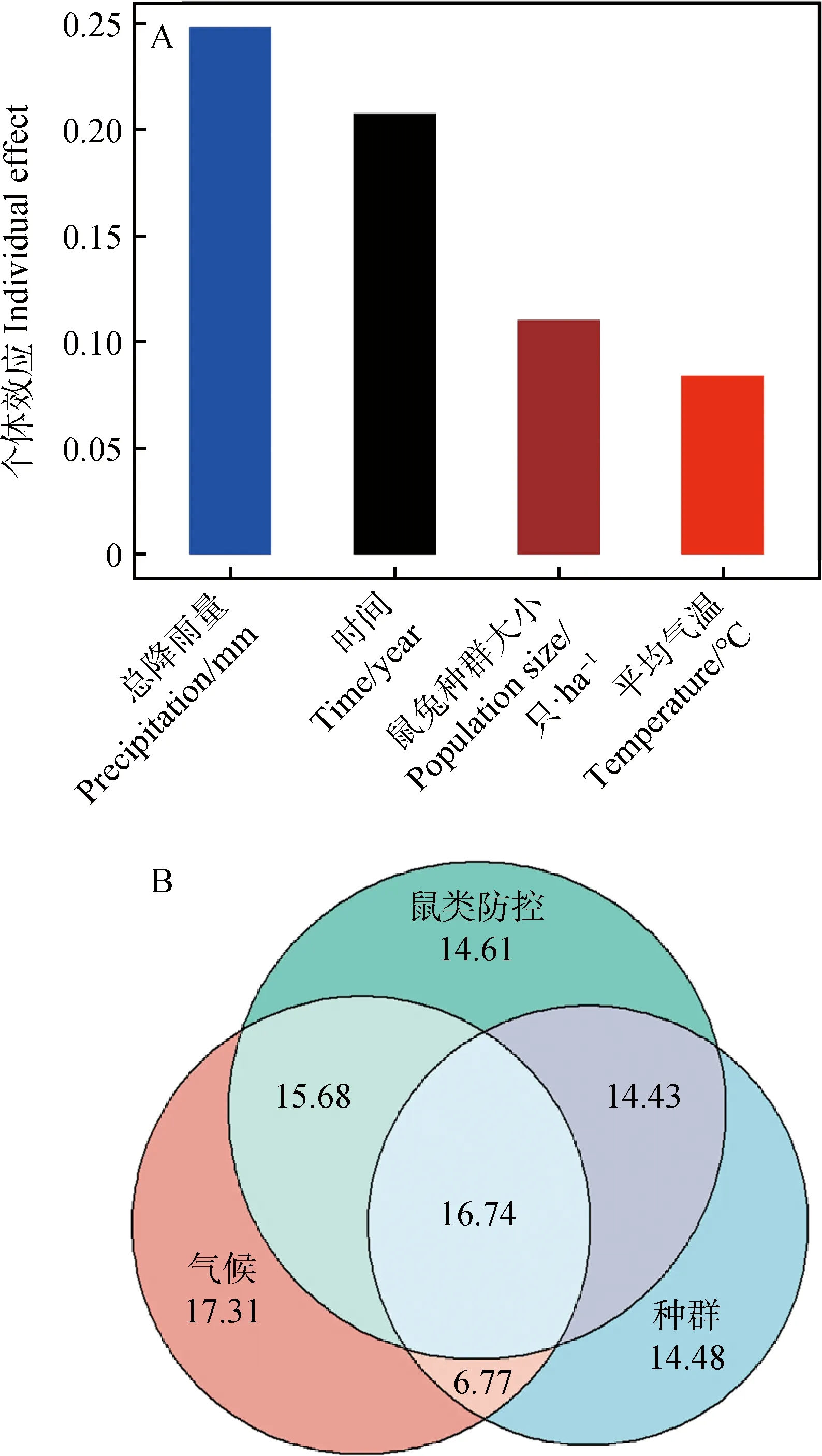
图4 生长季降雨量,生长季平均气温,鼠类防控以及鼠兔种群大小对植物群落地上生物量的影响
把降雨和温度视为气候因子,气候因子对地上生物量的影响是最大的,占总体的17.31%;鼠类防控对生物量的影响次之,占总体的14.61%;而鼠兔种群大小却是影响最小的,占14.61%(图4B)。交互作用分析表明,气候、鼠类防控和高原鼠兔种群大小三者的交互作用对植物群落地上生物量的影响最大,气候和鼠类防控的交互作用次之,鼠类防控和高原鼠兔种群大小的交互作用较小,而气候和高原鼠兔种群大小的交互作用最小。
3 讨论
3.1 多年鼠类防控对鼠兔种群动态的影响
随着年限增加,4月份属兔种群数量下降,而8月份和10月份的鼠类防控显著地降低了鼠兔种群数量。造成这一现象的原因有两个:其一是4月份高原鼠兔刚越冬完,其冬季死亡率高达91.04%[20],如果当年有低温或是雪灾,那4月份的种群数量就会受到极大的影响。再者,4月份是高原鼠兔的繁殖期初期,4月份的种群数量相对较小。其二是4月份监测的结果距离前一年10月份的监测过去了6个月,需要考虑到鼠兔的迁入和迁出。研究区域的防控区和非防控区相隔不远,灭鼠后由于鼠兔数量猛减,为毗邻的非防控区鼠兔的迁入创造了食物和栖息条件。而相对来说4月到8月,8月到10月间隔时间不久,迁入与迁出的影响相对较小,种群数量随时间有明显下降。因此,极端天气、防控的月份以及防控持续的时间都会影响鼠兔种群大小。为弄清气候(全年总降雨量和全年平均温度)与鼠类防控持续时间对鼠兔种群大小的影响,使用R的glmm.hp包进行多元混合线性回归分析,结果表明,气候因素对鼠兔种群大小影响的解释度可达42.77%,而鼠类防控对鼠兔种群大小影响的解释度仅为14.24%,气候和鼠类防控的交互效应强于它们的独立效应(图5)。
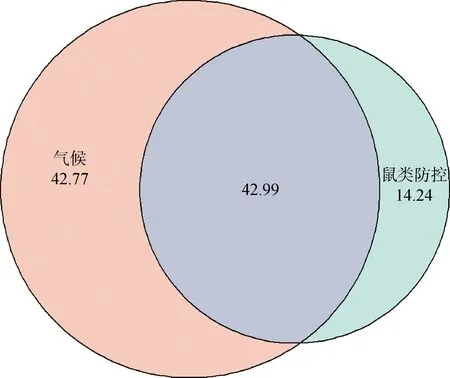
图5 气候与鼠类防控对鼠兔种群大小的影响及其交互作用
前四年鼠兔种群大小下降较快,后五年下降(图1B)的可能原因是:第一,前四年鼠兔种群密度处于较高的水平(2013年8月密度为140只·ha-1),且前3年的年降水量较低,地上生物量也较低(<350 g·m-2),这样每只鼠兔个体能获得的食物资源较少,因此鼠兔种群的出生率较低而死亡率较高,种群数量下降的也比较快。而第四年鼠兔种群密度已经减少到相对较低的水平(2016年8月密度为64只·ha-1),相较于第一年的最大密度减少了约50%。此时每只鼠兔个体获得的资源充足,出生率上升而死亡率下降,甚至其他区域内的鼠兔也会迁入该低密度区域以获得更多的资源,因此鼠兔种群数量恢复较快,导致四年后的鼠类防控效果不显著。第二,由于前四年鼠兔数量多,其天敌老鹰的数量也较多;鼠类防控后由于鼠兔数量减少,老鹰、藏狐等捕食者出现该区域概率较低,使得被捕食的鼠兔减少了,因而总体数量的下降趋势。
本文研究结果与梁杰荣等[11]的模型预测结果相似,即鼠兔种群大小随鼠类防控时间呈现锯齿状波动下降,而且鼠类防控频率、灭杀率以及持续防控时间都要达到一定的条件才能有效控制鼠害。本研究采取持续防控模式,每年捕杀3次,每次灭杀率大于25%,进而,鼠类种群数量才保持在较低水平。有研究表明,鼠兔种群的恢复速率受到鼠类防控方式的影响[21]。单一手段鼠类防控后2年鼠兔种群数量不仅恢复了,而且超过原有水平。人工捕杀与生态防治的综合治理措施对鼠类群落具有长期的干扰作用,表现为种群数量不稳定的波动,鼠类群落多样性、均匀度指数波动较大。如通过C型肉毒梭杆菌生物毒素进行两年灭杀后,配合围栏封育措施就能使鼠兔种群数量显著低于鼠类防控前[21]。鼠类防控后残鼠的数量恢复与其繁殖强度成正比,繁殖力越强,其数量恢复越快,种群数量呈受环境限制逻辑斯蒂增长曲线。单一灭鼠措施对鼠类群落的作用仅限定于灭鼠当年,危害鼠种种群恢复的周期短,鼠类群落的优势种仍为草地的主要危害鼠种,多样性、均匀度指数在灭杀期间下降,非灭鼠时期无明显变化[21]。此外,在长期使用抗凝血杀鼠剂灭鼠后,高原鼠兔产生了抗药性,导致灭杀效率降低[22]。这说明单一的捕杀方式、较低的捕杀率会导致灭鼠效率的低下,需要更长的时间才能控制住鼠害。目前主流的防治鼠害手段是人工捕杀与生态防治(C型肉毒素防治+毒杂草防除+虫害防治+补播+封育+休牧育草+保护天敌+合理放牧+恢复植被)[23]。因此,具有不同繁殖率的鼠兔种群,其在相同灭鼠率下,植被群落结构差异的情景下,恢复速率有所差异。
3.2 植物群落结构和功能的变化
在甘南藏族自治州,重度退化的草地在经过一次灭鼠处理以及补播后,莎草科、禾本科植物的盖度均有所增加,经过几年的演替后,毒杂草的盖度也逐步降低,植被总盖度呈递增趋势。莎草科、禾本科以及其他科杂草的生物量也有显著的提升[24]。这和本次研究的结果是一致的:植物群落结构随着鼠类防控持续时间发生了较大的变化,并在2016年有大幅度的增加(图2B,C)。注意到该时间节点与高原鼠兔种群终止高速率下降的时间节点一致(图1B),这表明连续多年鼠类防控使得高原鼠兔种群下降,进而促进植被盖度和地上生物量的恢复。通过分析植物群落植被高度、盖度和地上生物量与高原鼠兔种群之间的关系(图3),证实了鼠类防控在促进高寒草甸植被恢复中具有一定的作用。而在三个植物群落指标中,地上生物量和鼠兔种群大小之间的相关性是最密切的。这是因为高原鼠兔年采食牧草量占整个草地总初级生产力的21%到46%[25],随着高原鼠兔种群大小的增加,高寒草甸杂草斑、斑秃面积持续增大,进而使得草地生产力下降[26]。
除了考虑鼠兔对植物群落结构和功能的影响外,还需要考虑到其他因素的影响。2016年8月当地迎来了近几十年来最大的降水量(200 mm以上)与较高的平均气温,这会促进植物的光合并使地上生物量增加。对两个气候因素和生物量之间的相关性分析结果表明,总降雨量和生物量之间有显著的正相关关系,而平均温度与生物量之间则没有显著关系。此外,灭鼠不仅影响了当年的鼠兔种群大小,其多年的累积效应还会改善区域内的生态环境。线性回归变量重要性排序表明:在对不同因素个体效应的分析中(图4A),生长季的降雨量对生物量产生的个体效应是最大的,灭鼠时间和鼠兔种群大小的综合效应次之(图4B)。鼠类防控对植被地上生物量有影响,而气候变化(特别是降雨增加)可显著促进高寒草甸植被恢复。
4 结论
本文通过研究长时间鼠类防控(2013—2021年)对高原鼠兔种群数量和高寒草甸植物群落结构的影响,发现多年连续鼠类防控有效地降低了高原鼠兔种群密度,特别是防控前四年,其下降速率较大。同时,多年连续鼠类防控促进了植物群落盖度和生物量的恢复,特别是2016年及其之后植被盖度和生物量增加幅度较大。植物群落结构与高原鼠兔种群数量之间呈现显著的负相关关系,说明高密度的鼠兔种群数量降低了植物盖度和地上生物量,不利于植物群落的恢复。气候因子和鼠兔种群大小共同影响植物群落地上生物量,而且气候因子,特别是生长季的降水量,对鼠兔种群密度的影响大于鼠类防控持续时间的效应,同时其对植物地上生物量的影响也大于高原鼠兔种群密度的影响。因此,在研究鼠类防控对植物群落结构和鼠类种群大小影响时,气候是一个不可忽视的关键因子。此外本研究中,我们并没有考虑捕食者天敌(臧狐、臭鼬等)对高原鼠兔种群数量的影响。因此,多年的持续鼠类防控能有效降低鼠兔的种群密度,但其对植物群落结构和功能的影响仍不确定,未来需要更多的鼠类防控实验检验其对植物群落结构和功能的影响。
