砂岩型铀矿床微生物多样性及铀成矿作用
——以二连盆地哈达图铀矿床为例
2023-09-30林效宾李西得刘武生刘红旭刘旭
林效宾,李西得,刘武生,刘红旭,刘旭
(核工业北京地质研究院 中核集团铀资源勘查与评价技术重点实验室,北京 100029)
近几年,砂岩型铀矿床微生物成矿作用研究逐渐受到重视,越来越多证据表明微生物参与了绝大多数砂岩型铀矿床的成矿作用。研究现今铀矿床内部微生物菌落组成将有助于对微生物成矿作用的认识。早在20 世纪,在实验室条件下就已经证实H2S 对U(Ⅵ)的还原作用需满足溶液中U(Ⅵ)浓度远高于自然界水体中U(Ⅵ)浓度才可进行[1];有机质需在高温条件下(>120 ℃)才能直接还原U(Ⅵ),在低于120 ℃条件下,有机质对U(Ⅵ)的还原效率极低[2-4]。因此简单的认为砂岩型铀矿为U(Ⅵ)在氧化还原过渡带部位被有机质(生物碎屑、腐殖酸)、硫化物(黄铁矿等)、还原性气体(H2、H2S、CH4等)等还原物质还原为U(Ⅳ)而富集成矿,或认为微生物仅仅间接的参与了铀成矿等认识似乎还需要进一步进行商榷。
1962 年Woolfolk 等首次确定了微乳球菌可以还原U(Ⅵ)[5],之后1991 年Lovley 等首次在实验室证实某些细菌能将U(Ⅵ)还原为U(Ⅳ)[6],打开了微生物铀成矿作用研究的序幕,特别是近几年随着分析测试手段的更新换代,微生物参与铀矿形成的研究愈来愈受到众多学者所关注。国外有关铀的生物成矿作用研究主要集中于微生物与有机质的关系以及对微生物富集铀的人工合成实验研究上[6-9],同时在微生物成因矿物同位素组成、非生物和生物铀转化机制、微生物群落多样性及转录活性等方面均取得了进展[10-13]。相对而言,国内微生物铀成矿作用研究起步较晚,研究程度还相对较低,目前相关工作主要可划分为以下3 个方面:一是从矿物学、地球化学等方面直接或间接证据入手,例如根据铀矿物形态、磷元素含量、黄铁矿硫同位素、方解石碳同位素等特征来证实微生物参与了铀矿体的形成[14-23];二是通过开展微生物铀成矿作用模拟实验,推断主要细菌在砂岩型铀矿床的成矿过程中的作用[24-28];三是通过采集矿床岩心或地下水样品,在实验室进行微生物提取、扩大培养、测序等工作,进而确定铀矿床内部的微生物组成,探讨不同地球化学环境分带中微生物的多样性以及不同菌属对铀的迁移、富集成矿的影响[26,29-37]。上述第一个方面的工作可直观判定微生物是否参与了成矿作用,第二个方面工作可直接证实微生物在铀元素氧化还原过程中的功能和作用,第三方面工作主要能证实在矿床内存在大量的微生物菌落,虽然其结果与成矿期微生物菌落组成可能存在一定的差异,但现今与成矿期含矿含水层同样被深埋且受地表各类环境影响较小,可认为两个时期所处的地球化学环境、微生物菌落组成也相似。上述3 个方面的工作相辅相成,共同佐证了中国北方部分砂岩型铀矿床成矿作用与微生物密切相关。
自然界中砂岩型铀矿的形成环境远比实验室条件复杂,现今铀矿床内部的微生物种类及丰度虽然无法与成矿期相比,但只要现今铀矿床内的地下水仍存在微弱的水交替作用,就定会滋生一定量的微生物,而检测这些微生物菌落的组成,就成为了解成矿期微生物与铀成矿作用的一个窗口,这也是前文第三个方面工作可进行“将今论古”的基础条件。因在实验室微生物扩大培养的过程中培养环境与铀成矿时的环境差异较大,所以通过采集岩心或地下水样品提取、扩大培养后再进行测序的方法可能导致所得到的微生物组成及其占比与实际的微生物组成及占比发生显著变化,存在一定的缺陷。因为地层中的微生物主要生活在水体及岩石颗粒表面,本文主要尝试以二连盆地哈达图铀矿床为研究对象,利用大量水文孔地下水样品进行微孔滤膜过滤,直接利用过滤后的滤膜进行16S rDNA 扩增测序来鉴定微生物种类,从而避免环境改变而导致的微生物组成及其占比的变化,能够更直观的了解铀矿床不同地球化学带主要微生物组成及丰度,为微生物铀成矿作用提供数据支撑。
1 地质概况
哈达图铀矿床位于二连盆地乌兰察布坳陷齐哈日格图凹陷北部,该凹陷受东南部苏尼特隆起、西南部东方红凸起、西北部赛乌苏凸起和东北部塔木钦凸起所夹持,呈近南北向展布,长约60 km,宽10~25 km,区内发育有近东西向西拉木伦基底构造、北东向盖层内控盆构造和北西向新构造[38-40]。根据钻孔资料,自下而上揭遇地层有:下白垩统赛汉组下段(K1s1)、赛汉组上段(K1s2)、上白垩统二连组(K2e)、古近系(E)和新近系(N),矿区及附近地表仅出露古近系,且大面积被第四系覆盖,其他盖层无露头(图1)[39-41]。铀矿化主要产于下白垩统赛汉组上段第二旋回古河道砂体内,多定位于河道拐弯、变缓等水文情势发生变化的部位[42]。矿物学特征表明,黄铁矿为与铀矿物伴生的主要金属矿物,其形态主要为不规则小颗粒状、草莓状、立方体状等,其δ34S 值介于-45.0‰~5.0‰,指示有部分黄铁矿中的硫为微生物的脱硫酸作用成因;方解石胶结物的δ13CPDB值介于-9.5‰~-0.8‰,反映了其与生物成因有关[38]。草莓状黄铁矿的产出及碳、硫同位素组成特征,均指示微生物参与了哈达图铀矿床的成矿作用。该矿床成因有多种观点,可分为以渗入型氧化还原为主的成矿[42-44]以及渗出成矿[45]两种观点。
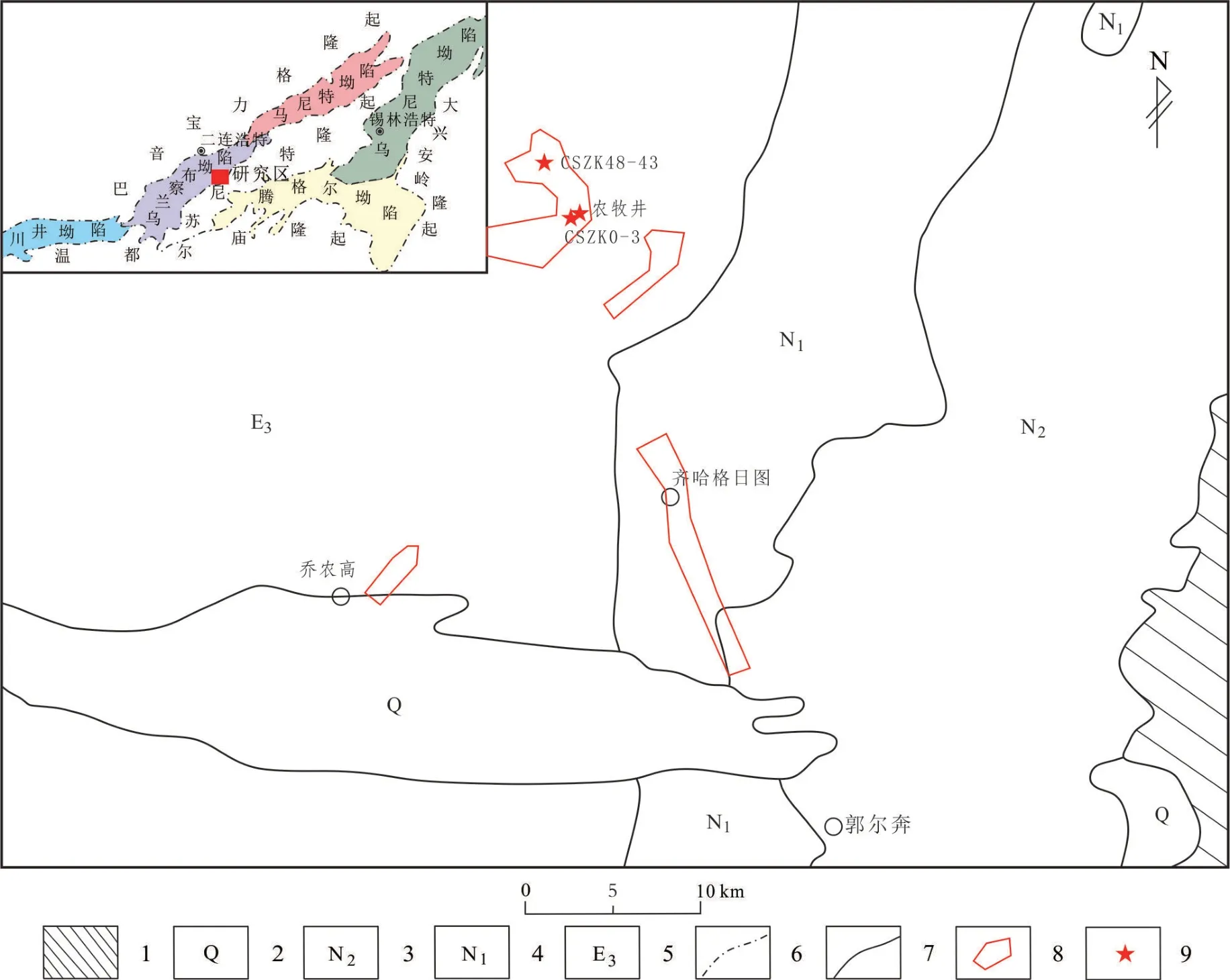
图1 二连盆地哈达图铀矿床地质简图Fig.1 Geology sketch of Hadatu uranium deposit in Erlian Basin
2 样品采集及分析测试
2.1 样品采集与处理
本文分别选取哈达图铀矿床2 个水文孔及1 个农牧井作为研究对象(图1),在水文孔滤水管上、中、下3 个深度各采集地下水样品5L 以上,采样时尽量避免对水文孔地下水的扰动,每次取样前对取样工具进行灭菌处理。本次共计采集7 件样品,现场进行各项水质参数的测定,测试结果见表1。所有采集样品当天用0.22 μm的微孔滤膜进行过滤,将过滤好的滤膜放入灭菌离心管内,做好标记后密封置于-18 ℃冰箱中保存,尽快干冰运输送至实验室,并进行细菌及古菌的16S rDNA 高通量测序工作。
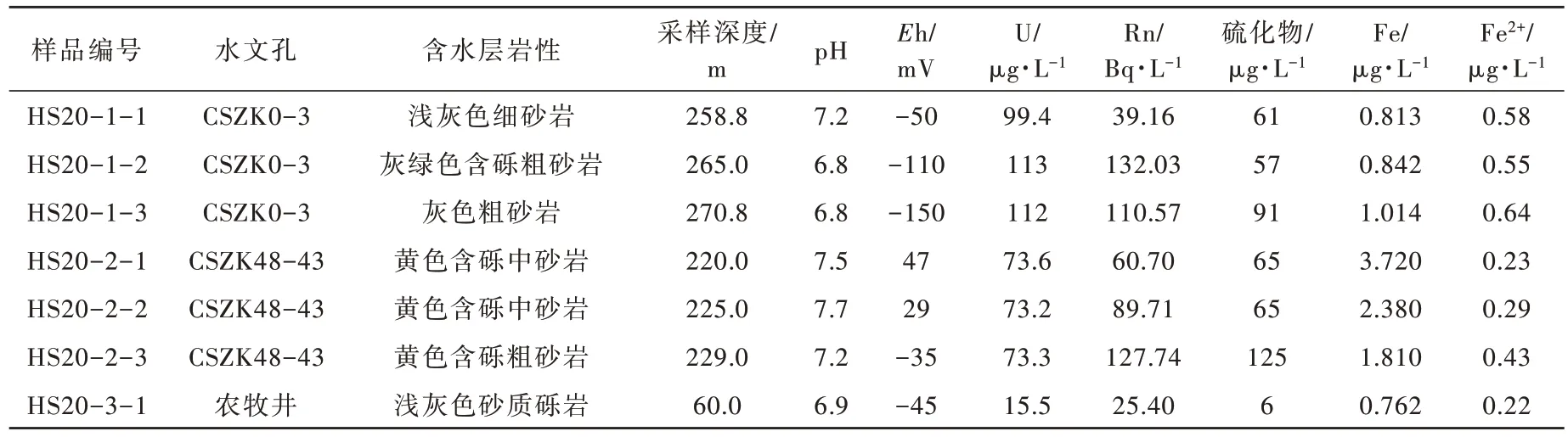
表1 哈达图铀矿床水文孔地下水定深采样样品信息Table 1 Groundwater depth and parameters of samples from hydrological borehole in Hadatu uranium deposit
2.2 样品分析测试
从DNA 提取到上机测序为实验室分析流程,主要包括以下步骤:1)微生物组总DNA提取;2)目标片段PCR 扩增,以细菌或古菌目标序列中的保守区域设计相应引物,并添加样本特异性Barcode 序列,进而对rRNA 基因可变区(单个或连续的多个)或特定基因片段进行PCR 扩增,在所送7 个样品中细菌PCR 扩增全部合格,古菌PCR 扩增仅CSZK0-3 号水文孔中的3 个样品合格,其余样品无法开展下一步测试工作;3)扩增产物磁珠纯化回收;4)扩增产物荧光定量,将PCR 扩增回收产物进行荧光定量,荧光试剂为Quant-iT PicoGreen dsDNA Assay Kit,定量仪器为 Microplate reader(BioTek,FLx800);5)测序文库制备,采 用Illumina 公司的TruSeq Nano DNA LT Library Prep Kit 制备测序文库;6)上机进行高通量测序,采用Illumina MiSeq/NovaSeq 平台对群落DNA 片段进行双端(Paired-end)测序。
3 分析测试结果
3.1 分类单元数统计
将高通量测序的原始下机数据进行一些列处理后,对于细菌或古菌的16S rDNA 基因,选 用Greengenes 数据库(Release 13.8,http://greengenes.secondgenome.com/)[46]进行物种分类学注释。通过对抽平后的ASV/OTU 表格进行统计,获得每个样本中的微生物群落在各分类水平的具体组成,计算不同样本在各分类水平所含有的分类单元数,采集的样品分类单元数越大,代表其含有的微生物种属种类越多。微生物群落中物种分类学注释,常用界、门、纲、目、科、属、种7 级分类注释系统,虽然用16S rDNA 测序可以得到最详细的分类等级为种,但由于测序片段通常只有300~500 bp,其大部分只在科水平和属水平上得到较高可信度的注释,由图2、3 样品测序数据分类单元数分布中可看出“种”这一水平分类单元数相对较少,而“科”、“属”这两个水平的分类单元数相对较多,所以在后文微生物种属的论述中将主要以属水平进行微生物种类的探讨。
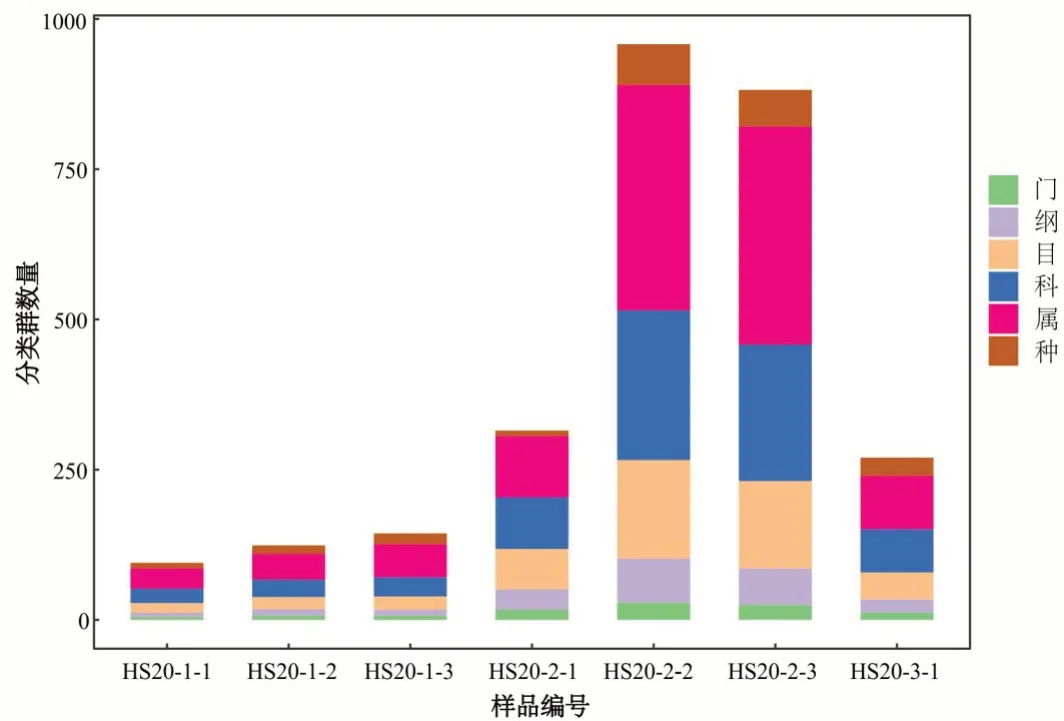
图2 各水平细菌分类单元数统计图Fig.2 Statistics of the number of bacterial taxon at each level
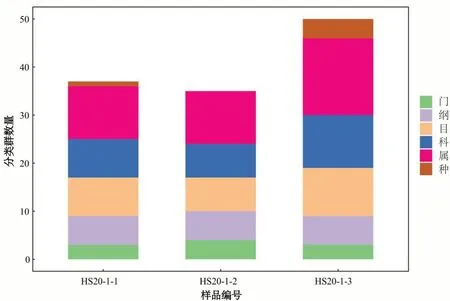
图3 各水平古菌分类单元数统计图Fig.3 Statistics of the number of archaeal taxon at each level
3.2 细菌属水平物种组成
细菌是原核微生物的一大类群,在埋藏较深的铀矿床内部仍旧活跃。综合统计哈达图铀矿床7 个样品16S rDNA 分析结果,分布较多的主要有以下细菌 :假红杆菌属(Pseudorhodobacter)、异常球菌属(Deinococcus)、不动杆菌属(Acinetobacter)、海洋杆菌属(Pelagibacterium)、盐单胞菌属(Halomonas)、游动球菌属(Planomicrobium)、动性球菌 属(Planococcus)、副球菌属(Paracoccus)、假单胞菌属(Pseudomonas)、涅斯捷连科氏菌属(Nesterenkonia)、类诺卡氏属(Nocardioides)、嗜冷杆菌属(Psychrobacter)、节杆菌属(Arthrobacter)、Saccharimonadales、Leucobacter、新鞘氨醇杆菌属(Novosphingobium)、罗尔斯顿菌属(Ralstonia)、Chloroplast、Subgroup_6、鞘氨醇单胞菌属(Sphingomonas)、Hymenobacter(图4)。整体看铀矿体部位地下水中好氧细菌和厌氧细菌并存,且以好氧菌占优。受地下水pH 值、Eh 值、含水层有机质含量等要素的控制,不同水文孔及不同深度采集的地下水微生物组成差别较大,在热图上差异性较明显(图5)。CSZK0-3 号水文孔地下水还原性较强,Eh 值范围为-150~-50 mV,pH 值范围为6.8~7.2(表1),赋水岩性主要为灰色砂体,其优势细菌组成主要为假红杆菌属(Pseudorhodobacter)、不动杆菌属(Acinetobacter)、游动球菌属(Planomicrobium)、动性球菌属(Planococcus)等,多为具呼吸代谢的化能异养菌属;CSZK48-43 号水文孔地下水还原性相对较弱,Eh 值范围为-35~47 mV,pH 值范围为7.2~7.7(表1),赋水岩性主要为黄色砂体,相较于CSZK0-3 号水文孔更偏氧化环境,其优势细菌组成主要为海洋杆菌属(Pelagibacterium)、盐单胞菌属(Halomonas)、假红杆菌属(Pseudorhodobacter)等,多为好氧化能异养型菌属;样品HS20-3-1为农牧井,含水层位为古近系,埋深浅,有机质含量低,其细菌组成主要为异常球菌属(Deinococcus)、副球菌属(Paracoccus)等。

图4 哈达图铀矿床地下水细菌属水平物种组成柱状图Fig.4 Histogram of horizontal species composition of bacteria in groundwater of Hadatu uranium deposit
3.3 古菌属水平物种组成
古菌是一群具有独特的基因结构或系统发育生物大分子序列的单细胞原核生物,多活在地球上极端的生态环境或生命出现初期的自然环境中,以自养和异养生活为主,其具有特殊的生理功能,如在超高温、高酸碱度、高盐及无氧状态下生活;具有独特的细胞结构,如细胞壁骨架为蛋白质或假胞壁酸,细胞膜含甘油醚键;以及代谢中的酶作用方式既不同于细菌又不同于真核生物。古菌按照其生活习性和生理特点,大致可分为3 个类群:极端厌氧的产甲烷菌、极端嗜盐菌以及在低酸和高温环境中生活的嗜热嗜酸菌[47]。根据16S rDNA 测序结果,仅CSZK0-3 号水文孔不同深度的3 个样品古菌扩增成功,这可能与所在赋水砂岩有机质的含量有关,CSZK48-43 号水文孔及农牧水井采样部位的砂岩有机质含量较低或几乎不发育。统计这3 个样品的古菌组成,含量排在前19 位的古菌主要为:甲烷叶菌属(Methanolobus)、Nitrososphaeraceae、Bathyarchaeia、Candidatus_Nitrocosmicus、甲 烷杆菌属(Methanobacterium)、暂定亚硝化螺菌属(Candidatus_Nitrososphaera)、Candidatus_Nitrosotenuis、Group_1.1c、Nitrosotaleaceae、鬃毛 甲烷菌(Methanosaeta)、Marine_Group_II、甲烷囊菌属(Methanoculleus)、甲烷短杆菌属(Methanobre-vibacter)、暂定亚硝化侏儒菌属(Candidatus_Nitrosopumilus)、盐薄片菌属(Halolamina)、Candidatus_Nitrosoarchaeum、甲烷螺菌属(Methanospirillum)、Hydrothermarchaeota、Candidatus_Nitrosotalea(图6)。按微生物功能可归纳为产甲烷菌(包括甲烷叶菌属、甲烷杆菌属等)、亚硝化菌(包括Nitrososphaeraceae、暂定亚硝化螺菌属等)及少量其他古菌(Group_1.1c、Marine_Group_II等)。
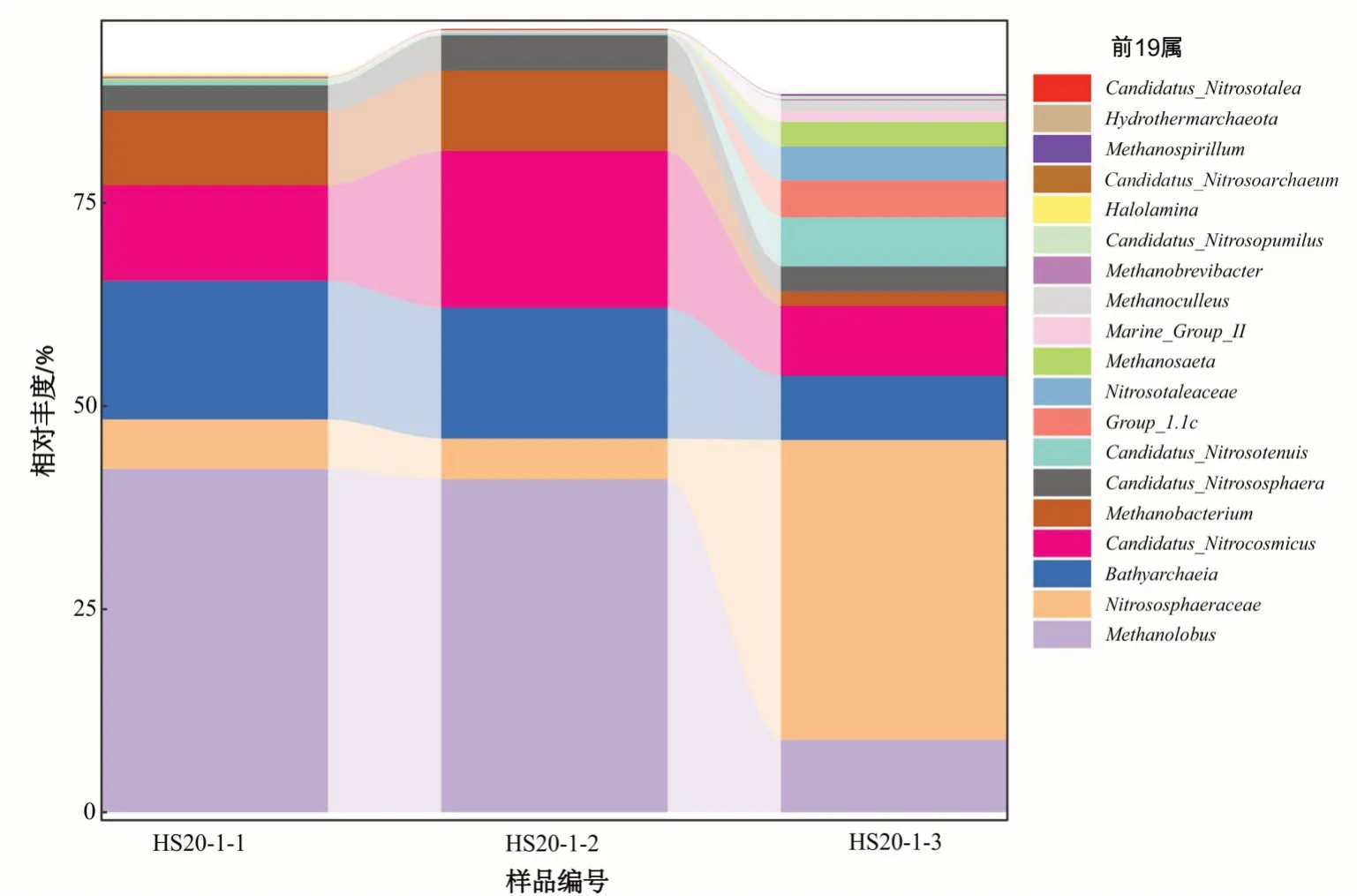
图6 哈达图铀矿床地下水古菌属水平物种组成柱状图Fig.6 Histogram of horizontal species composition of archaea in groundwater of Hadatu uranium deposit
4 讨论
哈达图铀矿床中的微生物数量大,种类多,并且分布广泛,证实了在该铀矿床内部不同地球化学环境带存在不同组合类型的微生物。按功能可以划分为硫酸盐还原菌、硫杆菌、产甲烷菌、硝化菌、亚硝化菌、反硝化菌、铁细菌等,无论矿床为渗入成因或渗出成因,这些细菌在矿床中的作用集中于参与铀矿床中碳、硫、氮、铁、铀等元素的氧化还原循环过程,在矿床地球化学元素循环中起到不容忽视的作用[4,47]。
4.1 微生物与铀矿床中碳元素循环
碳元素能以还原形式存在,例如甲烷和有机物,也能以氧化形式存在,例如一氧化碳和二氧化碳,在铀矿体深层封闭环境下,有机质被微生物分解的最终产物(包括氨离子、硫化物、甲烷)不断积累,成为其他微生物能量源[31,47]。哈达图铀矿床细菌和古菌的大部分种属主要以有机和无机碳作为营养来源,可分为化能异养型和化能无机自养型。绝大多数细菌属于化能异养型,例如假红杆菌属、不动杆菌属、游动球菌属、动性球菌属等,其碳源和能量均来自地层之中的有机质,利用有氧呼吸、无氧呼吸产生其生命活动所需的能量;古菌中的产甲烷古菌属为化能无机自养型,在铀矿层中占据着重要地位,其代谢的基本途径为以CO2作为电子受体,H2作为电子供体,自养生成甲烷和水,另一途径是在包括乙酸、甲醇、甲酸等含少量C 和O 的底物中异养生长产生甲烷。哈达图铀矿床地层中有机质的存在为包括碳循环细菌在内的各类细菌能量的主要来源,各类细菌的存在离不开有机质的存在,而该地区含矿层有机质相对缺乏,通过岩石酸碱度、共伴生元素等分布特征认为富矿石带可能具有深部油气或热水的改造作用[38],而深部流体所携带的有机质为微生物菌落提供了持续能源。哈达图铀矿床碳循环的主要路径为:含矿层部位原生或深部流体携带的有机质被地下水体中的微生物不断消耗,最终以CO2气体的形式逸散或随地下水迁出。
4.2 微生物与铀矿床中硫元素循环
参与地层中硫元素循环的细菌主要为硫氧化菌和硫酸盐还原菌(Sulfate-reducing bacteria,简称SRB)[47-48],前者在酸性水中分布较广,在硫化物氧化过程中起到重要作用,后者主要分布于厌氧环境水体中,能够氧化多种有机质,将高价态的硫酸盐还原为低价态的硫化物。两种细菌在铀矿床水体中可同时存在,SRB 还原硫酸盐的过程中为各种硫的氧化细菌提供电子供体,而硫的氧化细菌能够将低价态硫或单质硫氧化为硫酸盐,为硫酸盐还原菌提供生存所需的电子受体,这一循环过程构成了硫的氧化态和还原态的往复变化,为与硫有关的各类细菌的生长繁殖提供了必要的物质来源,两类细菌的共同作用促进了矿床中硫元素的循环。溶解氧一定的条件下,含水层中有机质含量越高,越有利于硫酸盐还原菌的生长[47,49-51]。在哈达图铀矿床水文孔地下水中参与硫氧化的细菌数量较少,主要为硫杆菌属(Thiobacillus),总量未达到0.001%;参与硫酸还原的细菌最高丰度为0.392%,主要有Desulfosporosinus、脱硫杆菌目(Desulfobacterales)、脱硫弧菌目(Desulfovibrionales)、除硫单胞菌目(Desulfuromonadales)、Desulforhabdus等,他们均为厌氧菌,主要功能为利用葡萄糖、乳糖、蛋白质、天门冬素、甘氨酸、丙氨酸、天门冬氨酸、乙醇、甘油、琥珀酸等有机质作为供氢体,将硫酸盐、亚硫酸盐、硫代硫酸盐和次亚硫酸盐等还原形成硫化氢。与同处于二连盆地的芒来铀矿床相较而言,哈达图铀矿床水文孔地下水中与硫循环相关的微生物占比要小的多(芒来铀矿床地下水中硫氧化菌与硫酸盐还原菌总丰度高达19.192%),原因可能与地下水交替强度及硫酸盐、有机质含量有关。受水质参数和围岩有机质含量差异的影响,不同水文孔SRB 的组成也有差异,有机质含量较高、还原性较强的CSZK0-3 号水文孔地下水主要为Desulfosporosinus,该细菌可通过将硫酸盐还原为硫化物来产生能量,并且与产生甲烷的古细菌竞争营养物质,可以减少古菌的活动,抵抗甲烷的形成;相对偏氧化环境的CSZK48-43 号水文孔地下水主要为脱硫弧菌目(Desulfovibrionales),其中HS20-2-1 样品SRB 含量极低;作为参照组的HS20-3-1农牧井地下水样品,未检测到任何与硫循环相关的微生物种群(表2)。
4.3 微生物与铀矿床中氮元素循环
铀矿床内部地下水氮元素主要以3 种形式存在:分子氮(N2),占水中气体的75%以上;有机氮化合物;无机氮化合物(NH3、NH4+、NO3-)。无论是化能异养型细菌或是化能自养型微生物的生长繁殖都需要从环境中获得所必须的氮源,在铀矿床的氮元素的地球化学循环中硝化菌和亚硝化菌、反硝化菌起到了主要作用[31,47]。它们通过代谢作用可将氨态氮氧化为亚硝酸盐、硝酸盐,也可以经反硝化作用将硝酸盐还原为氮气、氨态氮,从而促进矿床中氮元素的循环。自然界中含氮有机化合物经微生物分解后的产物为氨,经微生物(包括亚硝化菌和硝化菌)的硝化作用,转化为硝酸态或亚硝酸态氮。在哈达图铀矿床参与硝化作用的微生物主要为古菌中的亚硝化菌(包括Nitrososphaeraceae、暂定亚硝化螺菌属等);参与反硝化作用的微生物主要为细菌中的假单胞菌属(Pseudomonas)、盐单胞菌属(Halomonas)、副球菌属(Paracoccus)等,它们均能将硝酸还原为氮气,加之矿床地下水氧气的消耗,造成地下水中气体氮气占比增高。
4.4 微生物与铀矿床中铁元素的循环
铁循环涉及几种不同的微生物。涉及铁氧化的细菌主要有氧化亚铁硫杆菌、嘉利翁式菌属、硫化叶菌属、球衣细菌属及纤发菌属等,它们能完成铁的氧化,将亚铁离子转化为铁离子。铁还原发生在厌氧条件下,虽然很多微生物在它们的代谢过程中能还原少量的铁,但大部分铁的还原是通过特殊的铁呼吸微生物来实现的,如还原金属地杆菌、还原硫地杆菌等,它们以铁离子作为氧化剂,从有机物中获取生长所需的能量[48]。在哈达图铀矿床水文孔地下水中铁的氧化细菌占比较少,仅见有少量的纤发菌属等;而三价铁的还原菌较多,多种古细菌和细菌都能够把三价铁还原成二价铁,例如副球菌属(Paracoccus)、假单胞菌属(Pseudomonas)、硫酸盐还原菌(SRB)等。
4.5 微生物与铀矿床中铀的还原、富集
在典型的渗入铀成矿过程中,氧化环境中的地下水铀元素主要以二价铀酰离子存在,溶解度高,具有很强的迁移能力;在还原环境中,铀以四价离子形式存在,水中铀元素含量极低,其水迁移能力极低;有氧条件下,在铁细菌、硝化细菌、亚硝化细菌、硫杆菌等微生物作用下,黄铁矿及还原态的铀被浸出,氧化为Fe3+、SO42-、U(Ⅵ)溶解于含矿含水层中随水迁移;U(Ⅵ)离子随地下水迁移至不完全氧化带和矿石带后,三价铁还原菌、硫酸盐还原菌等微生物能够利用含矿含水层中所富含的有机质作为能源,将Fe3+还原为Fe2+,SO42-还原为H2S,并同时可将U(Ⅳ)还原为U(Ⅵ)。而渗出铀成矿过程,也有氧化还原的过程,来自深部高压、较高温度并富有机酸、氢气、铀等组分的流体在迁移至氧化砂体部位时,环境条件变化致使其中的物质产生沉淀[45]。在此过程中,微生物在有机酸分解、还原性气体氧化、铀的吸附沉淀等方面同样起到了一个不可或缺的作用。在不同的氧化-还原带部位,微生物的种类各不相同,随地下水径流方向不断向前滚动迁移,形成了矿床中铀元素的浸出、迁移、沉淀的循环,为微生物成矿作用提供了条件。在氧化还原过程中,Fe3+的氧化力可将U(Ⅳ)氧化为U(Ⅵ),所以在成矿过程中,只有当Fe3+被大量还原为Fe2+后,U(Ⅳ)才可大量生成,这也是为什么大多数砂岩型铀矿床内的铀矿物多附着于相伴生的黄铁矿表面或附近的原因。另外,根据前人对特定微生物菌种吸附铀的实验表明[31],在哈达图铀矿床地下水中还存在很多对铀具有吸附作用的微生物种属,例如假单胞菌属(Pseudomonas)、Bacillus等,这些细菌虽然占比相对较少,但其对铀元素的吸附容量却很大。SRB 可还原六价铀已经过大量实验所证实,且混合菌群对于六价铀的还原率要高于单菌株的还原率,而有机物在很大程度上影响微生物对铀的还原效率;死亡的SRB 也可以在一定程度上吸附体系中存在的游离的铀化合物[2,47-48,50-51]。
在砂岩型铀矿床成矿过程中碳、硫、氮、铁等元素循环与铀元素的循环密不可分,铀元素的溶解、迁移、还原沉淀离不开上述元素的氧化还原作用,虽然现今矿床内部地下水中微生物的组成并不能直接证明哈达图铀矿床成矿作用绝对与微生物作用有关,但不可否认微生物在铀矿床内部大量存在,而各元素的循环又离不开微生物的参与,硫、碳同位素证据表明微生物在铀矿床成矿期大概率参与了铀成矿作用[38,45],微生物通过对氧、碳、氮、硫、铁等元素价态的调节,直接或间接导致了铀元素价态的变化。具有成矿能力的微生物并不是单一孤立作用的,它们对碳、硫、氮、铁、铀等元素的迁移及氧化还原过程具有相互协调并相互制约的关系,所以深入研究铀矿床中微生物系统与成矿系统的关系,了解微生物在铀矿床不同地球化学带的生态分布及进一步探索生物成矿作用机理,将对认识砂岩型铀矿床成因和成矿机理具有深远意义[47,51]。
4.6 哈达图铀矿床微生物分布与铀成矿
哈达图铀矿床微生物组成与经典层间氧化带型铀矿床微生物组成相比较而言差异较大,主要表现在矿化带仍以好氧-兼性厌氧细菌为主,而还原性细菌占比较少。按照采样深度划分为3 个带,统计每个样品属水平的分类数量及占比前两位的属,可以看出3 个带的细菌种类和地球化学功能差异较大(表3):还原带细菌属水平分类数量最低,主要以假红杆菌属(Acinetobacter)、不动杆菌属(Acinetobacter)、动性杆菌属(Planomicrobium)等为主,这些菌属均可以烃类等有机物质作为碳源而生长繁殖,古菌以甲烷叶菌属(Methanolobus)、Nitrososphaeraceae、Candidatus_Nitrocosmicus、Bathyarchaeia等为主,能够利用有机物作为能量产生甲烷或将氨氧化;弱氧化带细菌属水平分类数量最高且复杂,主要为Pelagibacterium、盐单胞菌属(Halomonas)等以烃类有机物为碳源的菌属,未检测到古菌;上覆盖层为弱还原环境,主要菌属为异常球菌属(Deinococcus)、副球菌属(Paracoccus)等,以各种有机化合物作为碳源异养生长,其中异常球菌属为一类对引起细胞致死效应的辐射有极强抵抗能力的细菌,未检测到古菌。
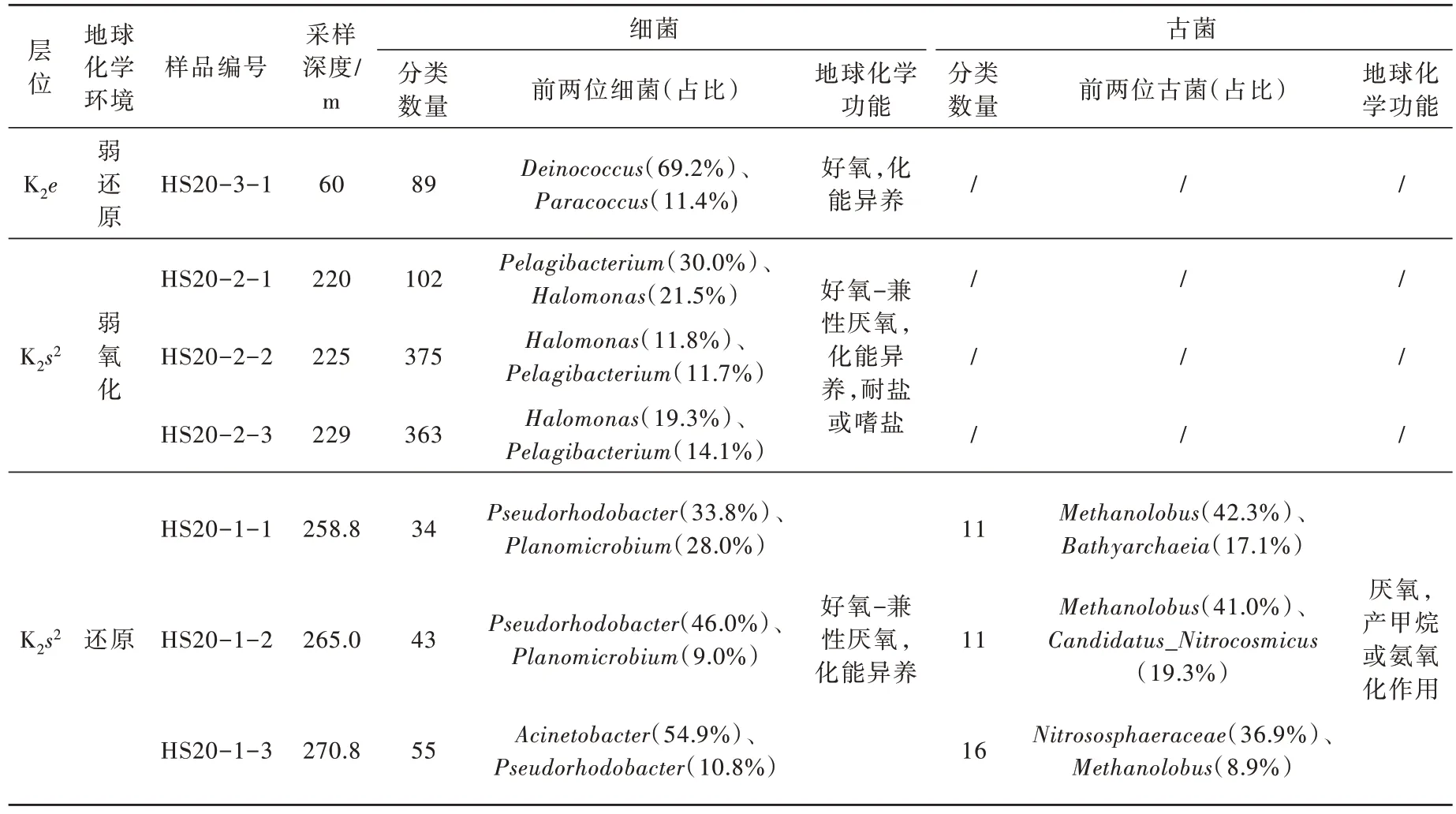
表3 哈达图铀矿床不同地球化学带属水平微生物分布特征Table 3 Distribution characteristics of microorganisms in different geochemical zones of Hadatu uranium deposit
从哈达图铀矿床细菌和古菌的分布情况来探讨铀成矿作用,笔者认为其与李子颖等[45]所提出的渗出铀成矿理论更为接近,深部灰色富有机质沉积建造在沉积成岩演化过程中,在一定物化条件下形成石油、天然气、氢气,同时产生各种有机酸(一般为富里酸和腐殖酸),这种富有机酸、氢气等组分的流体对磷、铀、钙、铁、硫等元素具有溶解迁移能力。当这些携带成矿物质具有相当高压力和温度的深部富有机质还原性流体,沿有利构造通道向上迁移至赛汉组上段红杂色砂体中时,温度和压力会明显降低,致使所处环境氧化性提高[45],细菌和古菌开始大量繁殖,其中以好氧菌为主,有机酸、H2等还原性物质被消耗,生成CH4、CO2、H2O等,此阶段主要以消耗易分解的轻烃为主,重烃则以地沥青的形式赋存在含矿砂岩之中,而所产生的甲烷在通常情况下非常稳定,能被围岩吸附或随地下水迁移,随着其逐渐积聚造成从富矿带至弱氧化带岩石中的CH4含量逐渐升高[38];原红色砂体三价铁矿物被逐渐还原为二价铁矿物,岩石颜色由氧化色转变为灰色、灰白色或灰绿色等色调,地球化学环境转变为以还原性为主,地下水pH 值、Eh 值较低,铁离子主要以Fe2+为主(表1),随着H2的消耗,H2S 开始与Fe2+反应生成FeS·nH2O,之后水陨硫铁与硫反应形成胶黄铁矿,脱水后逐渐变为黄铁矿;硫酸盐或硫化物在硫氧化细菌和硫酸盐还原菌作用下,可使硫元素在地下水中的同位素不断变重(CSZK0-3 孔δ34SV-CDT为6.2‰~6.3‰,CSZK48-43 孔δ34SV-CDT为7.4‰~7.7‰),而所形成的黄铁矿等矿物则表现为从核部至边缘硫同位素不断变轻[38,45];深部一同携带上来的氨气及铵盐在亚硝化及反硝化细菌共同作用下部分被转化为亚硝酸盐,该部位地下水中几乎不存在硝酸根离子;腐殖酸、甲酸、乙酸等有机酸能与铀及其他许多金属元素牢固结合形成有机络合物,使水中金属元素含量大大增高,随着H2、有机酸等物质被微生物所消耗,铀也开始发生沉淀,优先被先形成的黄铁矿、黏土矿物所吸附,虽然地下水中铀元素被卸载,但相较于传统层间氧化带型铀矿床铀矿带地下水而言,因有机酸含量较高在矿体部位地下水中铀含量要高出10~100 余倍(表1)。进入弱氧化带之后,pH 值、Eh 值进一步增高,地下水中细菌种类及数量急剧增加,各元素的循环异常活跃,在还原带未被分解的有机酸及烃等有机物在此带被大量降解,但其中环烷烃难以被微生物利用,在地下水样品析出气体中检测有约31.98×10-6的环己烷成分;弱氧化带参与硫元素循环的细菌数量依旧较少,以脱硫酸作用为主,SO42-稍微降低,H2S 含量与还原带相当,在地下水样品析出气体中检测有H2S、羰基硫、SO2等成分;残余Fe2+被携带至此带后,大部分已经被氧化为Fe3+,铁离子不断积聚,此时铁还原细菌数量较占优势;从还原带携带的氨气、铵盐及亚硝酸盐,在反硝化细菌及硝化细菌的共同作用下氮元素在N2、NH3、NO2-、NO3-等形态间转变,地下水中的硝酸根离子含量增至数个毫克每升(表1);地下水中有机酸含量进一步降低,虽然pH 值、Eh 值均增高,但铀含量相较还原带有所降低(表1),铀在红色砂岩中依然可以富集成矿。作为对比参照的二连组地下水,水文地球化学特征与赛汉组上段地下水较为相似,同样主受深部还原流体的控制,此段也形成了一定规模的铀矿化,但因含水层下部发育有厚层泥岩层,导致断裂向上的导水能力减弱,输入的有机质等还原物质较少,微生物发育种类少,主要为能适应极端环境的微生物,与硫、铁、氮元素循环相关的微生物几乎不发育或仅少量发育。
5 结论
1)二连盆地哈达图铀矿床地下水16S rDNA高通量测序结果表明,含矿层地下水中发育有大量的细菌和古菌,细菌以好氧细菌和厌氧细菌并存,且以好氧菌占优势,不同水文孔及不同深度采集的地下水微生物组成差别较大,主要受地下水水质参数、含水层有机质含量等要素的控制;古菌仅存在于围岩有机质含量较高的CSZK0-3号水文孔之中,主要为产甲烷菌及亚硝化菌。
2)矿床内存在的微生物与含矿含水层碳、硫、氮、铁、铀等元素循环密切相关,不同组合的微生物菌落通过参与各元素的氧化还原过程进而直接或间接参与了铀的迁移、富集与沉淀作用。
致谢:核工业二四三大队康世虎队长,核工业二〇八大队吕永华处长、任晓平高工、何大兔研究员为本文的野外样品采集给予了大量帮助,在此一并感谢!感谢匿名评审专家、编辑老师对本文提出的宝贵意见和建议。
