安徽大龙山国家森林公园冬季针叶树种光合特征
2023-09-28孙俊李锦隆赵郑郑邓鑫顾洪亮
孙俊,李锦隆,赵郑,郑邓鑫,顾洪亮
(1.安庆师范大学资源环境学院, 安徽 安庆 246133;2.福建师范大学福建省植物生理生态重点实验室,福建 福州 350007)
光合作用是植物与大气间进行二氧化碳交换的主要途径,是森林生态系统能量流动和物质循环的基础。光合能力的强弱也会决定着森林的初级生产力,是反应不同树种生存竞争力的主要特征。然而,大部分的国内外研究更多关注生长季的植物光合特征而忽略了非生长季的光合作用,这不利于明晰植物不同生活阶段碳获取策略的调整规律。
净光合速率(Pn)是反映光合能力和光合潜力的重要指标之一[1]。Pn直接影响植物的生长发育,同时受叶片的解剖结构[2]、功能性状[3]、生长阶段和环境特征等的影响[4]。以往研究中已经发现植物的Pn与植物叶片功能性状间存在密切的协作关系[5,6]。例如,叶大小 (单叶面积) 和比叶面积 (单位干质量的叶片面积,SLA) 会随着光照强度的增加而降低[7]。即使在不同冠层位置,通过叶片比叶重,叶片氮、磷含量仍然可以预测亚热带植物的光响应特征[8]。因此,不同植物的比叶面积、比叶重等是反映植物光获取能力的重要指标。然而,比叶面积与光合速率间关系并不恒定。如有研究发现,披针叶野决明(Thermopsislanceolata)的净光合速率与比叶面积间呈显著的正相关关系[9],表明随着披针叶野决明的叶片增大变薄将有利于其提高光合产物产量。然而,也有其他研究发现,不同榛种质资源的光合速率与比叶面积间呈显著的负相关关系[10]。这些结果表明,比叶面积与光合速率间的关系可能会受到叶片生活型、种内变异、生长阶段等的影响。如不同生长季节的杉木比叶面积变化与最适光合(Aopt)间也呈现显著的负相关关系[11],而黄山松光合速率与比叶重间的关系在冬季时期存在显著负相关关系,但是在夏季并未表现出较好的相关性[12]。鉴于冬季的光合能力对植物维持生存具有重要意义,有必要加强对冬季植物光合特征与比叶面积间关系的研究。
叶片水平的碳利用效率(CUE)是叶片光合和呼吸共同作用的结果,是叶片碳平衡的重要表征[13]。随着研究尺度的增加,从简单器官叶片到生态系统群落,CUE的数值变化逐渐降低。CUE还会受到森林类型和环境的影响,如热带雨林和北方针叶林的CUE较低,而温带及亚热带的CUE数值偏高[14]。另外,降雨量也会显著影响CUE的变化[15]。有研究发现,兴安落叶松叶水平的CUE值均大于0.8,且受到叶片氮磷含量、叶绿素含量和比叶重的调节[13],而杉木冬季叶片的CUE较高于夏季,又说明冬季的碳利用效率对植物的维持生长具有重要意义[11]。然而,针对冬季针叶树种的CUE一直缺乏足够的认识。SLA是重要的叶片功能性状指标,其与CUE间的关系是联系植物经济性状和生理性状间的纽带,是认识植物的碳获取策略的重要窗口。
马尾松和杉木广泛分布于我国亚热带地区,生长迅速,年均固碳量大,是荒山造林的重要树种。尽管国内外已经针对杉木和马尾松的光合特征[16-18]、针叶功能性状等进行了部分研究[19-21],但将针叶功能性状与光合特征联系起来探究的并不多见,尤其是在冬季。
1 试验地概况
试验地设在安徽省大龙山国家森林公园(116°59′35″—117°05′15″ E,30°37′26″—30°41′33″ N),位于安庆市北郊。公园地处北亚热带季风湿润气候区内,气候温和,雨量充沛。最高峰海拔693.4 m,森林公园总面积约为40.18 km2。年平均气温约为16.5 ℃,降水量1 389 mm。年平均无霜期247 d,光照充足。公园内有植物约180余科,800多属,1 400多种,常见主要树种包括松、柏、枫、檫、竹等。
2 试验方法
2.1 光合及光响应曲线测定
于2021年12月对大龙山的马尾松林(Pinusmassoniana)和杉木(Cunninghamialanceolata)林各设置400 m2(20 m×20 m)样地3块。马尾松平均树高和胸径分别为17.5 m 和28.6 cm,杉木的平均树高和胸径分别为13.4 m和13.7cm。样方设置好后采集植物标本,采集时间为9:00—15:00。首先,利用光合荧光测量仪测定样本在不同光强下的光响应曲线特征。测量之前先用饱和光强对小枝针叶进行20 min的光诱导,使针叶气孔完全开放。将光强梯度依次设置为 2 200、1 800、1 400、1 000、800、600、400、200、100、80、50、20、10和0 μmol·m-2·s-1。当光合速率稳定后开始测定,记录不同光强下的针叶光合特征参数。每个树种测定9条光响应曲线,利用photosynthesis软件拟合光响应特征参数,得到光饱和点(LSP)、光补偿点(LCP)及表观量子效率(AQY)。
然后每样地选取3棵,每棵标准木上选取3个小枝上的针叶用光合荧光测量仪Li-6800进行离体测量。测量时将叶室光强设定为1 600 μmol·m-2·s-1,平均气温为15 ℃,大气平均湿度60%,大气二氧化碳浓度平均约为400 μmol·mol-1。每小枝测量3次光合数据,取平均值,并记录其叶片面积、蒸腾速率、最大净光合速率、胞间二氧化碳浓度、气孔导度和饱和水压差。
叶片水平的瞬时碳利用效率(ICUE)作为叶片碳平衡的重要指标,是光合与呼吸综合作用的结果[13]。ICUE由下列公式获得:
式中,Pnmax为最大净光合速率(μmol·m-2·s-1),Rd为暗呼吸速率(μmol ·m-2·s-1),其中暗呼吸速率为绝对值。
2.2 暗呼吸速率及比叶面积测定
测完光合特征参数后,将小枝放入保鲜袋尽快带回实验室,置入避光暗箱中进行暗适应3 h。然后利用光合荧光测量仪避光测定针叶的暗呼吸速率,除将光强设置为0 μmol·m-2·s-1外,其余设定参数与光合特征测量设定一致。每小枝测定3组数据后记录平均值,并利用EPSON扫描仪测定针叶的面积。将叶片75 ℃烘干至恒质量后称量干质量,计算单位干质量的叶面积(比叶面积SLA,cm2·g-1):
SLA=叶面积/叶干质量
2.3 数据处理分析
利用Mircosoft Excel和SPSS19.0软件统计叶片的光合速率、叶片面积和干质量。通过Origin9.0拟合曲线并分析指标间的相关性系数与显著性系数。在SPSS中利用T-test检验2个树种针叶的光合参数特征、暗呼吸参数特征、比叶面积和瞬时碳利用效率间的差异。
3 结果与分析
3.1 杉木和马尾松光响应曲线特征
由图1可知,非生长季的杉木和马尾松光响应曲线变化具有显著差异,马尾松的LSP为1 581.36 μmol·m-2·s-1,LCP为24.64 μmol·m-2·s-1,Pmax为12.40 μmol·m-2·s-1,均显著高于杉木的413.28 μmol·m-2·s-1、7.28 μmol·m-2·s-1和4.33 μmol·m-2·s-1。二者的AQY差异并不显著(表1)。

表1 杉木和马尾松在光响应参数特征(N=9)
表4 大花序桉不同无性系生长性状隶属函数法评价结果
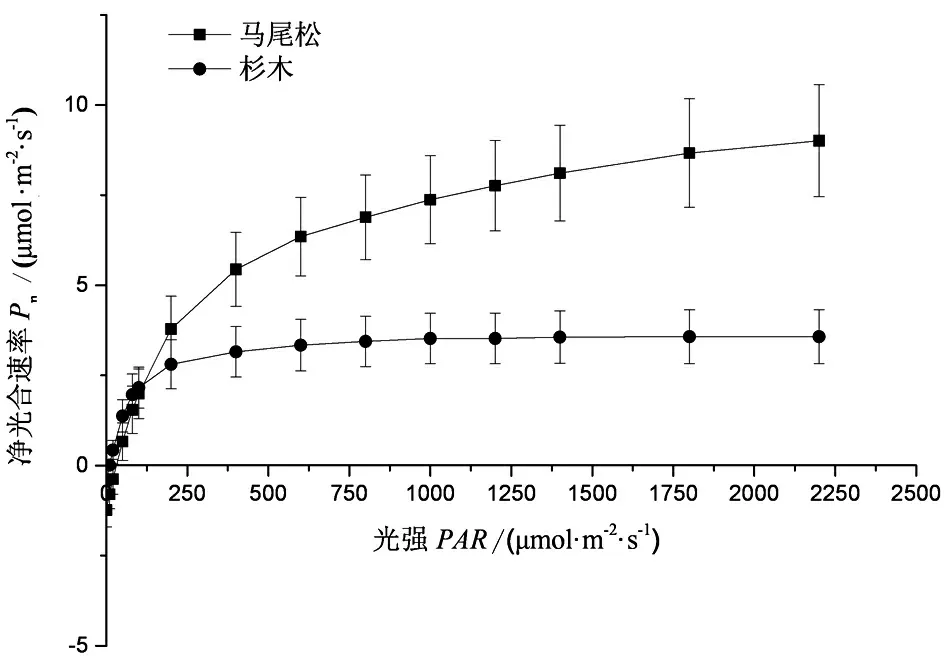
图1 杉木和马尾松的光响应曲线(N=9)
3.2杉木和马尾松冬季Pn、Rd、SLA和ICUE的差异
由图2可知,杉木和马尾松冬季的平均净光合速率Pn分别为2.69和8.76 μmol·m-2·s-1、平均暗呼吸速率Rd分别为0.65和1.82 μmol·m-2·s-1、平均比叶面积SLA分别为64.94和30.51 cm2·g-1,且具有显著差异,而叶片水平的平均瞬时碳利用效率ICUE则未显示出差别。

图2 杉木和马尾松暗呼吸参数特征、比叶面积和瞬时碳利用效率间的差异
由图3可见,Pn与SLA、Rd与SLA均呈现出显著的负相关关系,而ICUE与SLA之间的相关关系并不显著。这些结果表明,随着比叶面积的增大,杉木和马尾松的光合和呼吸速率会随之下降,但是对ICUE的影响并不明显。
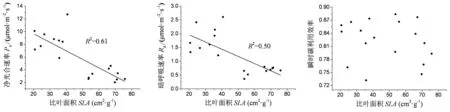
图3 杉木和马尾松针叶光合速率,暗呼吸及瞬时碳利用效率与比叶面积间的关系
4 讨论
4.1 杉木、马尾松的光响应特征
叶片与大气间的CO2交换性状是植物初级生产力的来源,在植物生物量的形成和积累中起着重要作用。宏观尺度的植物—大气间的碳固存模型,常被用于量化植物响应全球气候变化的热点问题,其中,最重要的是量化光合有效辐射与光合产量的关系[22,23]。这种关系在学界主要利用光响应曲线特征进行研究,而光响应曲线的影响因素可能会受到植物所处环境特征的影响[22,24]。在本研究中,2种针叶树种的光响应特征表现出显著差异,马尾松的LSP、LCP及Pnmax显著高于杉木。这表明马尾松和杉木的生态位分布存在差异,特别是光环境特征。本地区的马尾松生长优势明显,处在森林冠层的上层位置,能够获得充分的光照资源。有研究发现,在遮阴的条件下,植物叶片的气孔导度远小于不遮阴条件下[25],因此碳获取量可能会较低。本研究中,尽管杉木是速生的经济林树种,光合固碳能力强,但是由于其气孔导度为0.08 mol·m-2·s-1,低于马尾松的0.10 mol·m-2·s-1。因此,杉木的LSP、LCP及Pnmax均低于马尾松。LSP和LCP能够反映植物叶片对不同光强的利用能力,过去的研究表明,阴生植物叶片的LSP分布范围较广,从50~1 000 μmol·m-2·s-1,而LCP普遍低于20 μmol·m-2·s-1[26,27]。本研究中马尾松的LSP和LCP分别为1 581.36和24.64 μmol ·m-2·s-1,均高于这一范围。据此,本研究中认为马尾松的光照环境要显著优越于杉木。表观量子效率AQY通常能够反映植物在低光照下的自我调节能力,AQY的数值越大说明叶片吸收和转换光能的色素蛋白复合体越多,有助于提高植物低光照下的光资源利用能力[27,28]。本研究中的马尾松和杉木的AQY均值为0.07 mmol·mol-1,表明了杉木和马尾松在低光照的环境条件下拥有相似的光能资源利用能力。这一结果与2种树种的LSP、LCP和Pnmax存在显著差异的结果并不矛盾。如有研究发现,75%遮光处理对绣球(Hydrangeamacrophylla)的AQY影响与全光处理下的绣球并无显著差异,但会显著影响LSP、LCP、Pnmax、Pn及Rd[27]。因此,本研究认为,本地区的杉木和马尾松的光响应特性存在显著差异可能是因为自身所处的光环境差异造成的,但是这并不影响2种树对弱光环境的适应能力。冬季的气温较低,植物的光合碳获取更多的是为了维持生存。即使杉木的LSP、LCP、Pnmax、Pn及Rd普遍低于马尾松,作为亚热带分布最为广泛的物种,其可能主要得益于能够保持对低光环境下光资源利用能力。
4.2 杉木、马尾松SLA与光合特征间的关系
面对不同光环境,SLA是植物功能性状调节重要的指示特征[29],不仅能够表征植物的养分策略变化[10,30],还能够作为不同植物群落的普遍性特征[31],参与全球尺度的研究[5,32]。本研究发现,比叶面积在马尾松和杉木之间存在显著差异,2种针叶植物的比叶面积均较低,分布范围为20.62~75.88 cm2·g-1。有研究发现,在亚热带森林中,乔木植物平均SLA变化为76.9~461.8 cm2·g-1[33],而中国东部森林的灌木SLA变化为39.8~381.7 cm2·g-1[34]。这些结果显示阔叶物种的SLA高于针叶树种,也表明了针叶物种的叶片建造成本可能会更高。在冬季气温较低的情况下,较低的SLA能够提供更好的保温防栓塞性能,有利于针叶植物的安全过冬。
SLA与植物叶片功能性状间的关系研究主要集中在与Pn之间,本研究发现,杉木与马尾松的Pn与SLA整体上呈现显著的负相关关系。这可能是有以下几个原因:过去的研究发现SLA与Pn间既有负相关关系,也有正相关关系,但普遍认为具有高SLA的叶片将更多的氮分配给光合作用[35],从而增加其Pn。因此,即使有研究发现榛的种内Pn与SLA呈显著负相关关系[10],但是从草本植物披针叶黄华的结果看,具有高SLA的仍具有较高的Pn[9]。综合而言,Pn与SLA在阔叶和草本的种间比较上仍呈显著正相关关系。因此,这可能是针叶植物种间的Pn与SLA的关系受叶片氮分配的影响较小。另一方面,针叶物种叶片生长的年龄较大,较低的SLA显示其保守的性状进化策略特征。大多数植物的保守性状与获取性状沿“性状轴”呈现了不同的分布特征[36],这也说明较低的SLA与Pn、Rd之间可能是负相关的结论。遗憾的是,本文并未发现杉木和马尾松的ICUE与SLA间的相关性。前人研究认为,裸子植物的ICUE均值大约为0.80[37],这与本研究中的0.82与0.83具有相似性。无论如何,本文是将ICUE和SLA间的关系做了一次重要的尝试,这些结果也表明,在较低的SLA物种当中,确定其叶片氮分配和ICUE的变异可能会提高SLA在预测植物光合碳获取效率上的能力。
5 结论
本研究揭示了安徽省大龙山国家森林公园内2种针叶树种马尾松和杉木冬季的光响应特征及针叶功能性状间存在显著差异。马尾松的LSP、LCP及Pnmax显著高于杉木,马尾松的Pn、Rd显著高于杉木,但杉木的SLA显著高于马尾松。然而,二者的AQY和ICUE均无显著差异。综合来看,2种针叶树的Pn和Rd与SLA间均呈显著的负相关关系,但与ICUE的相关关系并不显著。因此,即使马尾松和杉木叶片之间的光合特征存在显著差异,但两种针叶树种冬季的碳获取效率存在相似性。另外,SLA能够调控马尾松和杉木冬季的Pn和Rd特征,但对ICUE的影响仍较薄弱。
