新丰江库区森林群落物种多样性随海拔梯度的变化格局*
2023-09-27吴国章甘先华张卫强黄芳芳王苏宇乔
吴国章 甘先华 张卫强 黄芳芳王 宁 苏宇乔
(1.华南农业大学 林学与风景园林学院,广东 广州 510642;2.广东省森林培育与保护利用重点实验室/广东省林业科学研究院,广东 广州 510520)
生物多样性是指一个区域内的所有植物﹑动物﹑微生物及各个物种所拥有的基因和各种生物与环境之间相互作用形成生态复合体和生态过程的总和[1]。全球气候变化和人类活动会加剧生物多样性的丧失﹑影响生态系统的稳定性,从而降低生态系统的服务功能[2-3]。物种多样性是对群落物种丰富程度的综合反映,是研究生态系统结构﹑功能及稳定性的基础[4-5]。环境因素对生物多样性变化的影响机制是生态学研究的重要主题[6],而海拔梯度对森林植物群落物种多样性影响的研究是近年来国内外众多生态研究学者关注的热点问题[7]。海拔梯度差异反映在温度﹑降水﹑光照﹑土壤等环境因子的变化,这些环境因子在海拔梯度上的变化比其在纬度梯度上的变化更加剧烈,由此产生的环境异质性影响着山地植物群落物种多样性的海拔分布格局形成[8-9]。因此,研究森林植物群落在不同海拔梯度上的变化,对于进一步深入研究生物多样性,揭示环境变化对森林生态系统稳定性的客观规律有重要意义。
目前对森林植物群落物种多样性随着海拔梯度变化的研究主要存在以下几种不同情况:包括物种多样性随着海拔升高而递减的负相关关系[10];物种多样性随着海拔升高呈现先升高后降低的趋势[11-12];在海拔梯度中间出现两次峰值的“双峰格局”模式[13];物种多样性随着海拔升高呈现先降低后升高的趋势[14-15];物种多样性随着海拔升高而升高的正相关关系[16];也有研究发现植物群落物种多样性跟海拔梯度没有明显相关性[17],并提出了水热动态假说﹑中域效应假说和环境能量假说等内在机制假说[18]。由此可见,植物群落物种多样性的海拔分布格局受到气候条件﹑地理环境和人为因素等影响而具有区域差异性。
β 多样性表示不同群落间物种组成的差异,Andrés Baselga 提出β 多样性可分解成物种周转组分和嵌套组分[19],前者指物种在不同群落间的更替导致群落间种类的差异[20],后者指拥有较少物种的群落是较多物种的群落的严格子集,物种增加或减少导致群落间种类的差异[21]。β 多样性分解有利于揭示区域间物种组成差异的形成机制,为生物多样性保护提供相应的保护对策。
新丰江水库是广东地区重要的水源地和水量调节枢纽,水域面积宽广,水质常年保持国家一类地表水标准,它维持着香港﹑深圳﹑东莞和惠州等地的用水安全和稳定,为粤港澳大湾区经济社会可持续发展提供了基本保障和重要支撑[22]。新丰江水库森林为库区提供了有效的生态服务功能,维护了库区生态安全格局,具有气候调节﹑水源涵养和生物多样性保护等重要生态效益。新丰江水库森林最高海拔超过1 000 m,目前新丰江水库森林的研究主要以不同的森林类型为对象[23],对植物群落物种多样性的垂直分布格局尚未见报道。本研究以新丰江库区森林群落为研究区域,采用样地法对植物群落进行详细调查,拟探讨以下问题:(1)新丰江库区森林群落物种组成和优势种分析;(2)植物群落α﹑β 多样性沿海拔梯度的分布规律;(3)周转组分和嵌套组分对物种组成差异的贡献。
1 材料与方法
1.1 研究区概况
新丰江水库位于广东省河源市东源县(114°15'~114°50'Ε,23°40'~23°10'Ν),是全国第七大水库和华南第一大水库,水域面积364 km2,多年平均水深28.7 m,最大水深93 m。属南亚热带季气候,冬夏季气候交替明显,雨热同期,年平均温度为20.7 ℃,年平均降水量为1 793.2 mm,年均相对湿度约为76%,降雨集中在3—8 月。土壤类型为黄壤和红壤,质地以中壤土和重壤土居多。新丰江库区周围为低山丘陵,植被类型为典型的南亚热带常绿阔叶林。
1.2 样地调查
在新丰江国家森林公园内170 m﹑280 m﹑525 m﹑738 m 和1 031 m 海拔地段内设置固定样地进行植物群落调查,每个样地内设置4 个20 m×20 m 的乔木样方,对样方内树高≥1.5 m 的木本植物进行每木检尺,记录种名﹑树高﹑枝下高﹑胸径和冠幅等信息;在每个乔木样方内沿对角线设置4 个5 m×5 m 的样方作为灌木样方,调查高度<1.5 m的所有木本植物的种类﹑个体数﹑高度及盖度等;在每个灌木样方内随机设置1 个1 m×1 m 的小样方作为草本样方,调查并记录草本植物种类﹑个体数﹑盖度和高度;使用GPS 定位,记录经纬度﹑海拔﹑坡度﹑坡向﹑坡位等生境基本信息。
1.3 优势度分析
优势度分析指标包括多度﹑相对多度(Relative abundance)﹑频 度(Frequency)﹑相 对 频 度(Relative frequency)﹑基面积(Basal area)﹑相对显著度(Relative prominence)和重要值(Importance value)等[24]。具体计算公式如下:
式中,ni为第i个树种的个体数,N为全部树种的总个体数。
式中,Fi为第i个树种的样方频数,F为所有树种样方频数之和。
式中,AB,i为第i个树种的胸高断面积,AB为全部树种的胸高断面积总和。
式中,乔木以胸高断面积表示相对显著度,灌木和草本以盖度表示相对显著度。
1.4 α 多样性计算
植物群落α 多样性用Margalef 丰富度指数﹑Shannon-Wiener 多样性指数﹑Simpson 多样性指数﹑Pielou 均匀度指数来测度[25-26]。计算公式如下:
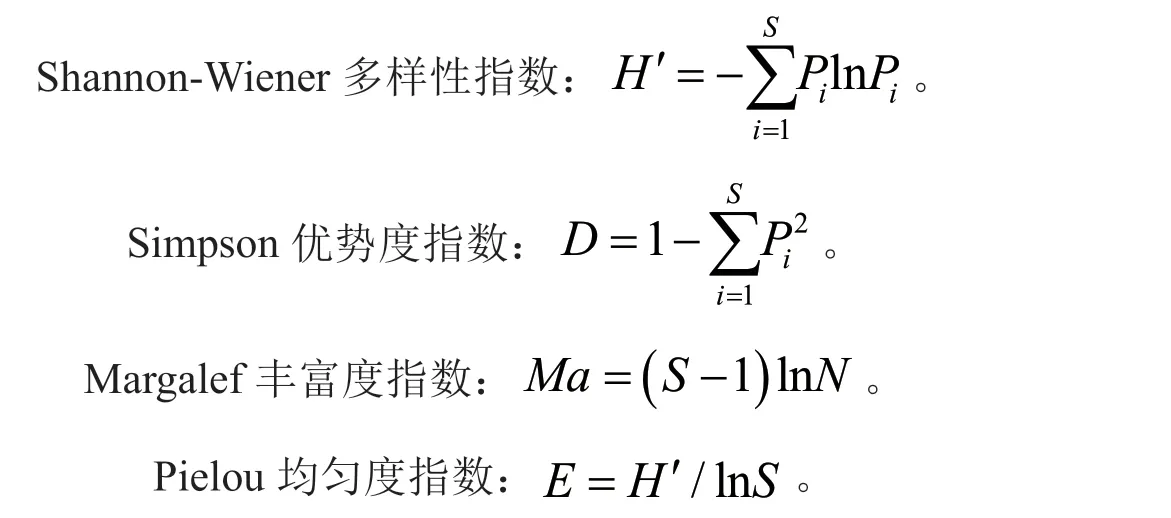
式中,S为物种数,N为物种总个体数,Pi为物种i的相对重要值。

表1 样地基本信息Table 1 Information of sampling sites
1.5 β 多样性计算及分解
本文采用物种丰度的Bray-Curtis 距离指数和基于存在和缺失数据(Presence-absence data)的Sørensen 指数以及Jaccard 指数作为配对样方间β多样性的度量,使用Baselga 等[19,27]提出的β 多样性分解方法,结合Legendre 等[28]对该方法的总结和改进,将群落间物种组成差异(即β 多样性)分解成周转组分和嵌套组分。

式中,βsor﹑βsim和βsne分别表示Sørensen 指数法中的β 多样性及其周转和嵌套组分,βjac﹑βjtu和βjne分别表示Jaccard 指数法中的β 多样性及其周转和嵌套组分,a为共有物种数,b和c分别是两个群落各自特有的物种数。Bray-Curtis 距离指数﹑Sørensen 相异性指数和Jaccard 相异性指数用于表征不同海拔的植物群落两两之间的物种相异程度,其值在0~1,数值越大表明2 个群落不相似程度越大。
1.6 数据分析
使用Excel 2021 软件对植物调查数据进行预处理,并完成植物组成统计和重要值计算。通过R 4.2.2 软件vegan 包和betapart 包分别分析植物α 多样性和β 多样性及组分。使用SPSS 25.0 软件对多样性数据进行单因素方差分析,采用Origin 2021 软件绘图。
2 结果与分析
2.1 植物群落的物种组成分析
在所调查的5 个样地中一共包括235 种维管植物,分别属于82 科151 属,其中蕨类植物11科17 属19 种,裸子植物3 科3 属3 种,被子植物68 科131 属213 种。数量优势科依次是五列木科﹑茜草科﹑樟科﹑壳斗科﹑蔷薇科﹑报春花科﹑叶下珠科﹑桑科﹑豆科和禾本科,共计10科51 属102 种,分别各占总科﹑总属﹑总种数的11.0%﹑29.5%﹑27.1%,单科单属单种占比为17.2%。其中,乔木层植物有108 种,灌木层植物有143 种,草本层植物有82 种;物种数在样地Ⅰ到样地Ⅴ中分别为156﹑167﹑141﹑122 和137 种,个体数分别为777﹑1 039﹑1 066﹑623 和891 株。
物种重要值可综合反映出某个物种在群落的地位与作用,是判定群落内建群种和优势种的重要依据[29]。新丰江库区不同海拔森林群落乔木层﹑灌木层和草本层的优势种存在较大差异(表2),乔木层重要值大于10%的在样地Ⅰ中有厚壳桂Cryptocarya chinensis(11.85%)﹑山乌桕Triadica cochinchinensis(11.12%),样地Ⅱ有米锥Castanopsis chinensis(16.90%)﹑鼠刺Itea chinensis(10.53%),样地Ⅲ有刺毛杜鹃Rhododendron championiae﹑鸭 公 树Neolitsea chui﹑栲Castanopsis fargesii﹑木荷Schima superba,样地Ⅳ有红楠Machilus thunbergii(27.77%)﹑浙江润楠Machilus chekiangensis(20.44%),样地Ⅴ红楠占绝对优势,占比39.26%。灌木层重要值大于10%的在样地Ⅰ有九节Psychotria asiatica(17.99%),样地Ⅱ有山血丹Ardisia lindleyana(12.55%)﹑九节(10.45%),样地Ⅲ有鸭公树(27.78%),样地Ⅳ灌木层重要值均小于10%,样地Ⅴ有木莓Rubus swinhoei(10.15%)。草本层重要值大于10%的在样地Ⅰ有芒萁Dicranopteris pedata(25.05%)﹑扇叶铁线蕨Adiantum flabellulatum(12.35%),样地Ⅱ有团叶鳞始蕨Lindsaea orbiculata(23.93%)﹑黑莎草Gahnia tristis(22.22%),样地Ⅲ有珍珠茅Scleria ciliaris(10.85%)﹑深绿卷柏Selaginella doederleinii(10.70%),样地Ⅳ有狗脊Woodwardia japonica(18.48%),样地Ⅴ有常春藤Hedera nep-alensisvar.sinensis(19.32%)﹑狗脊(16.83%)。
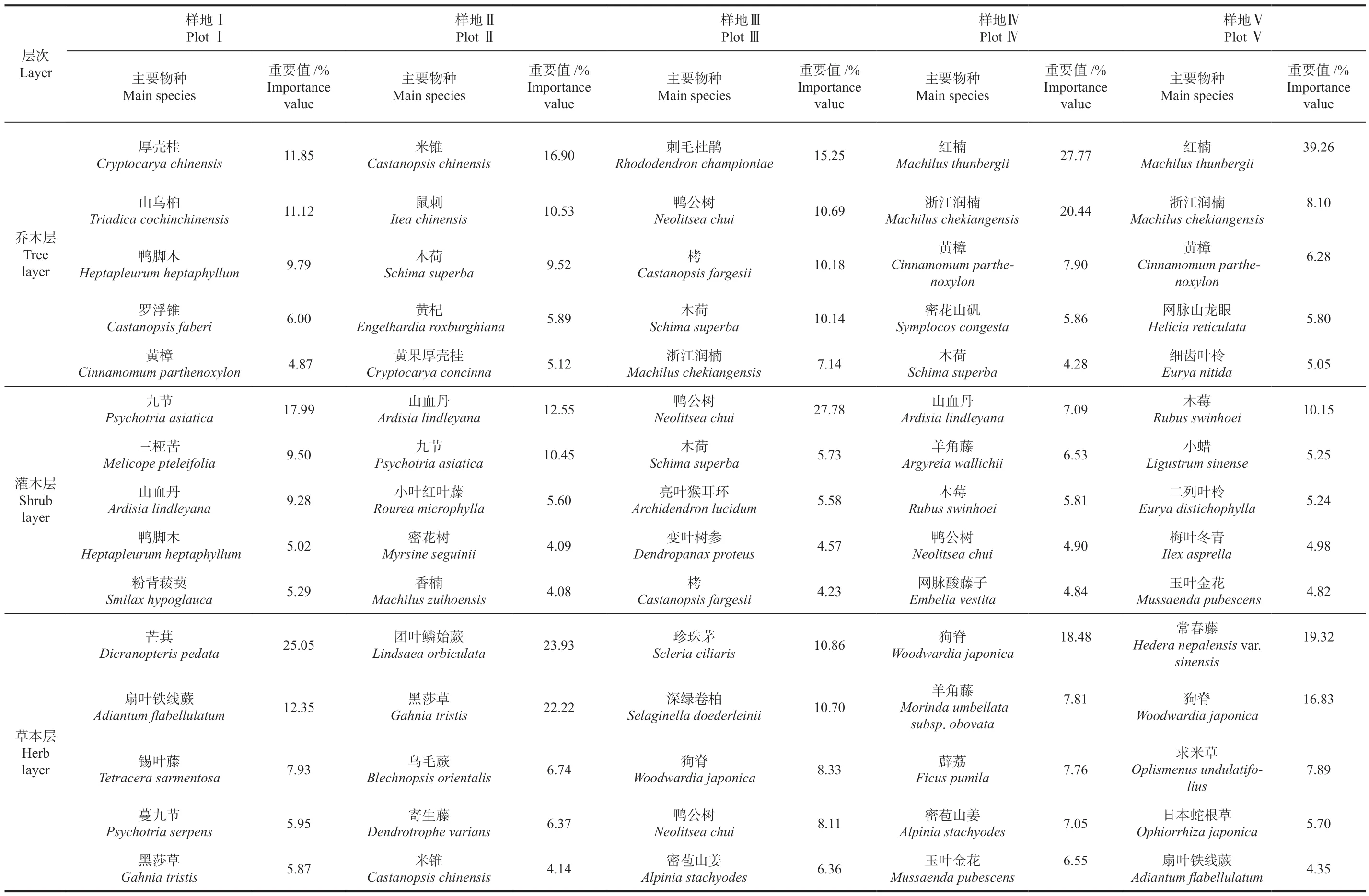
表2 不同样地主要植物组成及重要值Table 2 Composition and importance value of main plants in different plots
2.2 植物群落α 多样性沿海拔梯度的变化
沿海拔梯度各样地的α 多样性通过Margalef丰富度指数﹑Simpson 优势度指数﹑Shannon-Wiener 多样性指数和Pielou 均匀度指数进行表征。 乔木层α 多样性指数结果显示,随着海拔的增加,乔木层的Simpson 指数﹑Shannon-Wiener 指数﹑Margalef 指数和Pielou 指数均呈现出下降的趋势(图1)。从 灌木层α 多样性来看, 除了Margalef指数随海拔上升而逐渐增加外, 其他多样性指数最低值出现在中海拔525 m 处, 呈现出高﹑低海拔两边高,中海拔低的U 型格局(图2)。从草本层α多样性来看,Simpson 优势度指数﹑Shannon-Wiener 多样性指数和Margalef 丰富度指数呈现出单峰格局(图3),峰值出现在中海拔525 m 处,但各海拔间多样性指数波动较小, Pielou 指数峰值在中高海拔738 m 处,通过方差分析可知,草本层除Pielou 指数外其他3 种指数在不同海拔间均无明显差异。从植物群落的不同层次来看,各α 多样性指数均表现出灌木层>乔木层>草本层的特征。
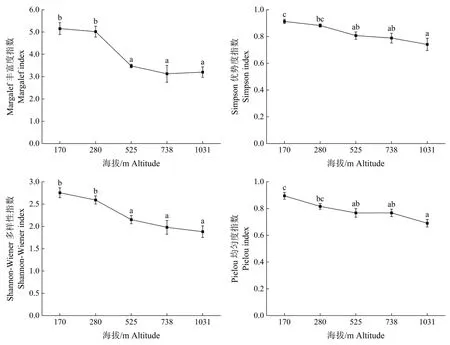
图1 不同海拔梯度下乔木层的α 多样性指数Fig.1 Alpha diversity indexes of tree layer at different elevations

图2 不同海拔梯度下灌木层的α 多样性指数Fig.2 Alpha diversity indexes of shrub layer at different elevations

图3 不同海拔梯度下草本层的α 多样性指数Fig.3 Alpha diversity indexes of herb layer at different elevations
2.3 植物群落β 多样性及分解
各海拔梯度样地间的β 多样性通过Bray-Curtis 距离指数﹑Sørensen 相异性指数和Jaccard 相异性指数进行表征。海拔相邻的两样地间,3 种指数显示出物种组成存在较大差异(图4),Bray-Curtis 指 数 介 于0.57~0.82 之 间,Sørensen 指 数 为0.46~0.60,Jaccard 指 数 为0.63~0.75。随 着 海 拔上升,基于物种有无的Sør ensen 指数和Jaccard指数呈现出相同的变化趋势,最大值出现在样地Ⅲ~Ⅳ(525~738 m)之间,最小值出现在样地Ⅰ~Ⅱ(170~280 m)之间,而考虑到物种个体数的Bray-Curtis 指数则显示样地Ⅱ~Ⅲ(280~525 m)之间拥有更大差异性,最小值出现在样地Ⅳ~Ⅴ(738~1 031 m)之间。
β 多样性是指不同群落间物种组成的差异,由物种周转(或物种替换)和嵌套(或丰富度差异)这两种过程决定。本研究使用BAS 法对Sørensen 相异性指 数和Jaccard 相异性指数分解成物种周转组分和物种嵌套组分。结果表明,两种指数的物种周转组分均在95% 以上,而物种嵌套组分均小于5%,物种周转组分明显大于物种嵌套组分,随着海拔上升,物种嵌套组分占比逐渐降低(表3)。因此,在海拔梯度上各植物群落物种组成差异由更替控制。
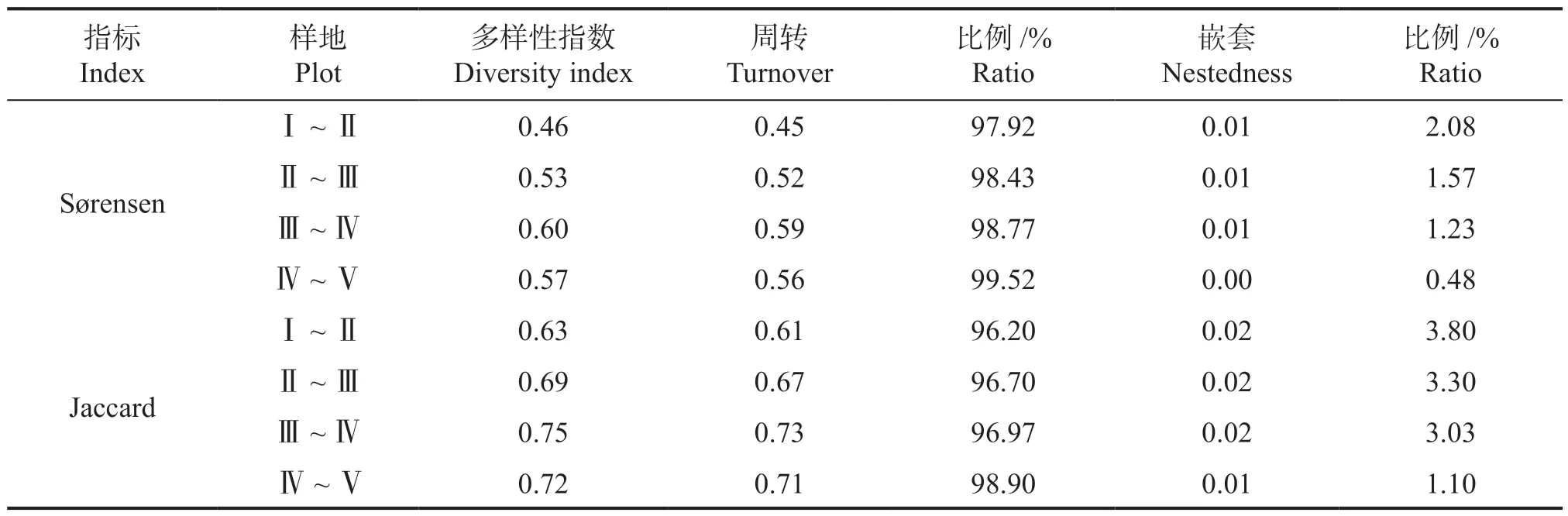
表3 不同样地的β 多样性指数及其分解组分Table 3 Beta diversity indexes and their components in different plots
不同海拔森林群落独有物种数量在物种周转过程中发生变化,影响着物种组成差异。结果显示,不同海拔独有物种整体的重要值在群落的不同层次上范围在17.98%~67.16%之间(表4),而单种独有物种重要值平均只有2.62%,其中大于5%的只有样地Ⅰ的扇叶铁线蕨(12.35%)﹑锡 叶 藤Tetracera sarmentosa(7.93%)﹑ 蔓 九 节(5.95%),样地Ⅱ的团叶鳞始蕨(23.93%)﹑九节(10.45%)﹑寄生藤(6.37%)﹑黄杞(5.89%)﹑小叶红叶藤(5.60%)﹑黄果厚壳桂(5.12%),样地Ⅲ的刺毛杜鹃(15.25%)﹑珍珠茅(10.86%)﹑深绿卷柏(10.70%)﹑栲(10.18%)﹑密苞山姜(6.36%)﹑ 蜂 斗 草Sonerila cantonensis(5.71%)﹑亮叶猴耳环(5.58%)﹑金毛狗Cibotium barometz(5.30%), 样 地Ⅳ羊 角 藤(7.81%)﹑ 薜 荔(7.76%)﹑山血丹(7.09%)﹑密苞山姜(7.05%)﹑玉 叶 金 花(6.55%)﹑ 乌 毛 蕨(6.03%)﹑ 乌 蕨Odontosoria chinensis(6.00%)﹑ 木 莓(5.81%)﹑淡竹叶Lophatherum gracile(5.48%)﹑团叶鳞始蕨(5.45%),样地Ⅴ的常春藤(19.32%)﹑求米草(7.89%)﹑网脉山龙眼(5.80%)﹑日本蛇根草(5.70%)﹑小蜡(5.25%)﹑细齿叶柃(5.05%)。

表4 不同样地独有物种的数量和重要值Table 4 The number and importance of unique species in different plots
3 讨论与结论
本研究结果表明,新丰江库区森林群落不同层次植物物种多样性指数随海拔的变化趋势并不一致。乔木层的Simpson 指数﹑Shannon-Wiener指数﹑Margalef 指数和Pielou 指数随着海拔上升均显示出明显的下降趋势,这与王秀云等[30]﹑李旭华等[31]﹑唐志尧等[32]对物种多样性海拔分布格局的研究结果类似。森林群落的海拔和至水域距离是影响植物多样性的关键因素[33-34],光照﹑水分和热量等环境因子随着海拔升高而发生变化,由于新丰江水库大面积水体蒸发的原因使得低海拔区域空气水分更加充足,气温变化较稳定,另外高海拔地区土壤厚度较薄,养分稀少,因此低海拔地区相比高海拔地区拥有更适宜植物生长的水热和养分条件。灌木层植物除了Margalef 指数随着海拔增加而增加外,Simpson 指数﹑Shannon-Wiener 指数﹑Pielou 指数呈现出中间低两边高的格局,可能是由于鸭公树优势种现象明显,其重要值为27.78%(其他灌木均小于6%),数量为288 株(占比为49.23%),其他灌木的生长受到抑制,从而造成了灌木层在中海拔地区分布不均匀,同时也使得灌木层物种多样性相比其他海拔要低。草本层植物的Simpson 指数﹑Shannon-Wiener 指数和Margalef 指数在海拔梯度上虽有波动,但各群落间没有显著的变化差异,这与贾美玉等[35]﹑周阳等[36]﹑高远等[37]的研究结果相似,同时草本层植物各项多样性指数均明显小于灌木层和乔木层,这可能是草本层除了受海拔梯度整体影响外,草本植物物种多样性主要受制于群落结构[38],草本植物于乔木层和灌木层之下,在光照﹑热量和水分资源的竞争中处于劣势,上层植物对光热资源的拦截削弱了在海拔上光照﹑水热差异对草本植物生长的影响。乔木层对海拔梯度变化的响应相对灌木和草本更加敏感[39],乔木相对灌木和草本要高大,暴露在空气中的顶芽更容易受海拔上升带来的温度变化影响,因此海拔越高,乔木层物种多样性越小[40-41]。因此,水热条件差异可能是影响新丰江库区森林群落植物α 多样性海拔变化的重要因素。
β 多样性度量时空尺度上物种组成的变化,体现了群落物种组成的差异[20,42]。结果表明,除了Sørensen 指数在170~280 m 海拔样地间为0.46,新丰江库区其他相邻海拔样地间β 多样性均大于0.5,说明样地间不相似程度较高,物种组成具有较大差异。基于物种有无的Sørensen 指数和Jaccard 指数随海拔上升先增加后减少,最大值出现在525~738 m 中间偏上的海拔,而基于物种丰度的Bray-Curtis 指数则显示峰值出现在280~525 m中间偏下的海拔位置,可能因为海拔280 m 和525 m 处拥有最多的物种个体数,加剧了两个不同海拔的样地间的物种组成差异,3 种β 多样性指数峰值都与中海拔有关,说明群落更替速率在中海拔处增加。
β 多样性分解是将物种周转(或物种替换)和嵌套(或丰富度差异)两种过程对总体beta 多样性的作用进行拆分,有助于进一步探究群落物种组成差异的驱动机制[43]。通过对Sørensen 和Jaccard 两种相异性指数进行分解,结果显示物种周转组分明显大于物种嵌套组分,可知在海拔梯度上各植物群落物种组成差异由更替控制,这与梁红柱等[44]对太行山东坡中段植物多样性垂直分布格局的研究结论相似,可能是因为海拔梯度带来水热条件的变化加剧了环境异质性,为植物提供了更多样的生存环境[45]。虽然物种数在不同海拔梯度上存在较小程度的波动,但通过β 多样性的分解可知物种丧失或增加对物种组成的贡献明显不及物种周转或替换。β 多样性分解成周转和嵌套两个过程有助于对不同地区或不同群落有针对性地制定生物多样性的保护策略,在以嵌套组分为主的生境应着重保护物种多样性高的区域[46],本研究中β 多样性分解结果显示物种周转占主要地位,反映了新丰江库区不同海拔森林群落独有的物种较多,但因单物种的重要值在不同层次中较低,物种在竞争中处于劣势,所以在生物多样性的监测中尤其需要关注独有种和重要值低的物种数量的动态变化。
本研究通过调查新丰江库区森林群落发现乔木层多样性指数随着海拔升高,呈现下降趋势;灌木层除了Margalef 指数随海拔上升而逐渐增加外,其他多样性指数呈现出两边高,中间低的U型变化趋势;草本层除Pielou 指数外其余3 种指数在不同海拔间均无明显差异。这意味着该新丰江库区森林群落的乔木层对海拔梯度的变化更为敏感,相对于灌草层更容易受到温度和水分变化的影响,因此在植物群落保护政策中应该着重关注乔木层生长特征﹑结构动态和物种多样性变化。除了Sørensen 指数在样地Ⅰ~Ⅱ中为0.46,3 种相异性指数在其他样地间均大于0.5,群落间物种组成差异主要由周转组分控制,这意味着新丰江库区不同海拔森林群落间的物种组成差异较大,不同的植物群落独有种较多,建议在新丰江库区从低海拔地区到高海拔地区采取全面﹑整体﹑严格的植物群落保护措施,对不同海拔的区域实行统一规划和管理。
本研究揭示了海拔梯度对新丰江库区森林群落α 多样性和β 多样性的影响,以及周转和嵌套组分对β 多样性的贡献,为新丰江库区森林群落生物多样性保护和森林生态系统的保护提供了一定的理论依据。本研究局限于尚未涉及土壤﹑区域小气候和地理距离等其他因素对物种多样性变化的解释,综合考虑多方面因素对物种组成的影响能够更深入地探究生物多样性的环境梯度格局和背后的驱动机制以及加强对物种多样性的保护。
