跌水方式和水生植物对水体溶解氧含量的影响特征
2023-09-25韦建华李茵茵杨钙仁
韦建华 李茵茵 杨钙仁
(广西大学林学院 广西南宁 530004)
治理水体污染是一项复杂的系统工程,其中生物方法是近年来发展迅速的一种生态技术,其利用人工曝气充氧、天然跌水曝气复氧等方法提高水体溶解氧含量(DO),通过利用和强化微生物的生命活动,对水体中污染物进行转移、转化及降解,从而使水体得到净化[1]。在不同溶解氧含量条件下,DO值越大,污染物去除率越高[2]。
城市封闭水体流动性小、复氧速率慢,水体的自净功能弱,人工强化增氧可有效提高水体的自净能力[3-4]。跌水增氧法是增加污染水体中溶解氧含量的有效且清洁的方法,广泛应用于水污染治理及水景观工程中[5]。重力跌水是氧气从空气向水体的传质过程,影响传质效率因素较多。已有研究发现,跌水增氧效果与跌水高度及跌水流量有关[6],跌水高度、跌水断面宽度越大,增氧效果越好[7],但目前针对不同跌水方式间的增氧效果差异了解不多。植物根系是湿地氧气的重要来源,并显著影响湿地的污染物净化功能[8]。水生植物在生长良好的情况下可从根系分泌氧气,其分泌速率主要受光照强度、气温、通气组织等调控,泌氧使得湿地形成好氧、兼性好氧和厌氧区,总体上提高了根周微生物活性,促进污染物质的降解和去除[9-10]。但目前对公园常用水生植物的泌氧功能了解并不全面与深入,限制了此类植物在应用时的科学设计。本研究拟通过不同跌水方式和水生植物水培试验,了解和量化几种跌水方式以及不同水生植物间的增氧差异,为重力跌水和水生植物的水体增氧应用提供科学依据。
1 材料与方法
1.1 材料
1.1.1 器材多孔角钢支架、塑料水箱(不同容积)、不锈钢水槽、黑臭水体、250 mL针筒、自配无氧水、自配污水、红色绑绳、PVC管、海绵。
1.1.2 植物狐尾藻(Myriophyllum verticillatum)、金鱼藻(Ceratophyllum demersum)、苦草(Vallisneria natans)、凤眼莲(Eichhornia crassipes)、睡莲(Nymphaea tetragona)、萍蓬草(Nuphar pumilum)、黄花鸢尾(Iris wilsonii)、千屈菜(Lythrum salicaria)、再立花(Thalia dealbata Fraser)、风车草(Cyperus alternifolius)、美人蕉(Canna indica)、花叶芦竹(Arundo donax)、梭鱼草(Pontederia cordata)。
1.2 方法
1.2.1 重力跌水试验
1.2.1.1 跌水装置设计设置3种跌水装置,分别为台阶式、坡流式和瀑布式,装置由进水区、跌水区和出水储存区3个部分组成,跌水高度均为3 m。3种装置的进水区均由2个100 L的塑料大桶构成,出水储存区均为一个20 L的塑料桶。3种跌水装置的跌水区构造如下。
台阶式跌水区:由多孔角钢支架和6个长30 cm、宽30 cm、深25 cm的水箱构成,对准箱口一宽边居中割出一宽度为20 cm、深度为5 cm的排水口,2个相邻水箱的出水口高差(即每级跌水高度)为50 cm(图1-a)。坡流式跌水区:由多孔角钢支架、台阶式跌水区水箱和一长424 cm、宽20 cm、深5 cm的水槽构成,水槽倾斜度为45°(图1-b)。瀑布式跌水区:由台阶式跌水区水箱和一长3 m、宽20 cm、厚5 cm垂直于地面放置的水槽构成(图1-c)。
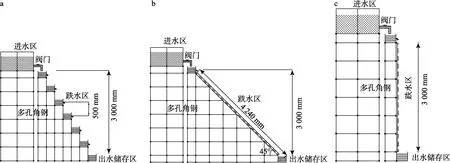
图1 跌水装置
1.2.1.2 指标测定试验用水均采自广西大学东校园碧云湖,中度黑臭,用水量为200 L,跌水时长均为180 s。各试验重复3次。分别于第60、120和180 s,用250 mL针筒采集出水储存区水样,置于250 mL容量瓶,并迅速用碘量法测定其溶解氧含量(DO)。
1.2.2 植物根系增氧试验
1.2.2.1 供试植物选择依据观赏价值高、易获取且病虫害少等特点,参考相关文献,确定9种供试植物(表1),用于测定其对水体溶解氧含量的影响。试验前用自来水对所有植物进行7 d的预培养试验,选择标准株,称量其鲜重;然后烘干,称量其干重,计算每种植物的生物量。试验时,控制每个桶内的生物量均为30.4 g。
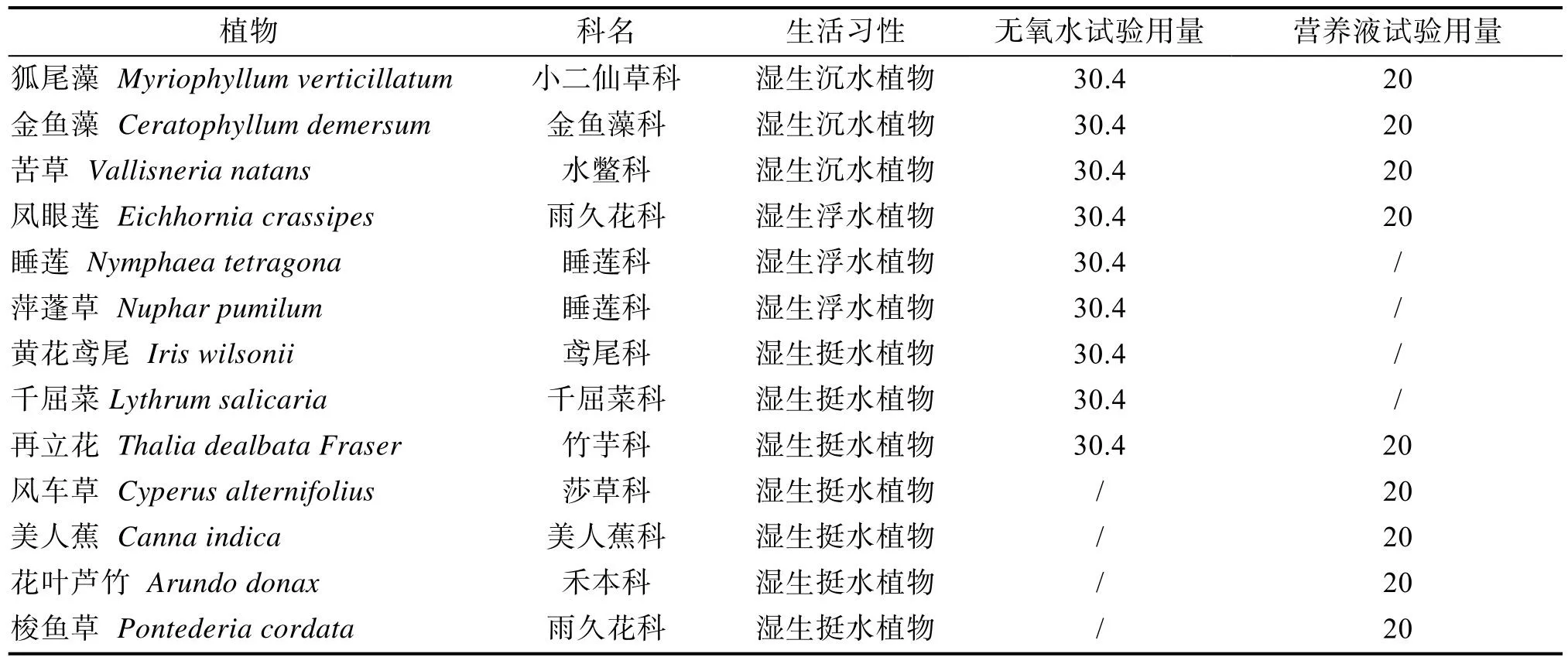
表1 供试植物基本信息
待采用自配无氧水水培的水生植物预培养完成后,对9种供试植物进行简单的分析,根据其对水体溶解氧含量影响的大小,进行了初步筛选;在接下来的自配营养水水培水生植物试验中,除去了睡莲、萍蓬草、千屈菜和黄花鸢尾4种植物,增加了风车草、美人蕉、梭鱼草、花叶芦竹4种挺水植物。在第二次水生植物对水体溶解氧含量影响试验前,用自配营养液对所有植物进行7 d的预培养,而后选择标准株,称量其鲜重;再烘干,称量其干重,最后计算每种植物的生物量。在试验时,控制每个桶内的生物量均为20 g。
1.2.2.2 指标测定自配无氧水由自来水中加入消氧剂配置而成,消氧剂包括二氯化钴(催化剂)和亚硫酸钠。相关化学方程式为:
自配污水根据霍格兰法,对水中N、P、COD等进行配置,前5个参数参考富营养化水体设置。配置好的污水各指标及浓度分别为:COD 50 mg/L、TN 8 mg/L、氨氮4 mg/L、硝氮4 mg/L、TP 2.8 mg/L、DO 9.2 mg/L。无氧水试验于夏季室外进行,见图2。在塑料桶(桶高20 cm,上部直径30 cm,下部直径20 cm)内盛放9 L自配无氧水,而后将根系差异较小的沉水植物和浮水植物均匀放置在小桶中。挺水植物选取植株和根系相对一致的9株,放置于桶中,每桶放置3株,用红色绑绳将其固定在桶中间。9种植物均设置1个空白对照值,所有处理均设置3个重复,共30组。除空白组外,每个桶内的水生植物生物量均为30.4 g。此后,在每天的8:00,12:00,16:00,20:00进行取样,连续2 d,测量温度、溶解氧含量2项指标。每天采集完水样后,用无氧水补充至初始水面高度(20 cm),补充被蒸发及采集使用的水。
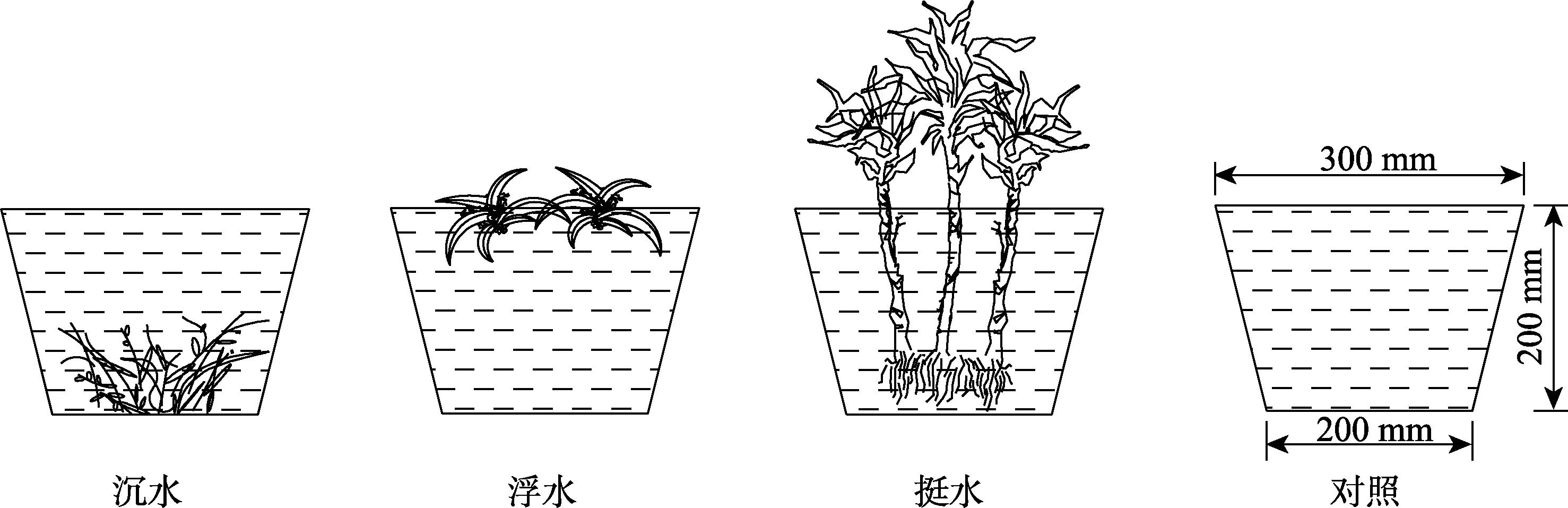
图2 无氧水试验装置
污水试验于冬季室内大棚进行,见图3。在塑料桶(桶高45 cm,上部直径35 cm,下部直径28 cm)内盛放9 L试验用水。沉水植物选取植株和根系差异较小的,均匀放置在桶中间。浮水植物和挺水植物同样选取植株和根系差异较小的,在桶的上面放置一块方格PVC板,剪出小孔,让植物通过小孔,并用海绵围住剪口,以起到固定植物的作用,确保只有根部浸没在水中。试验设置为9个植物处理和1个空白处理,共10个处理,每种处理设置3个重复,共30组。浮水植物和挺水植物的种植密度均为每桶4株,沉水植物若干,每个桶内的生物量均一致,均为20 g。每隔5日进行取样,共4组数据,为期20 d,测量温度、溶解氧含量2项指标。每次采集完水样后,将试验用水补充至初始水面高度,补充被蒸发及采集使用的水。
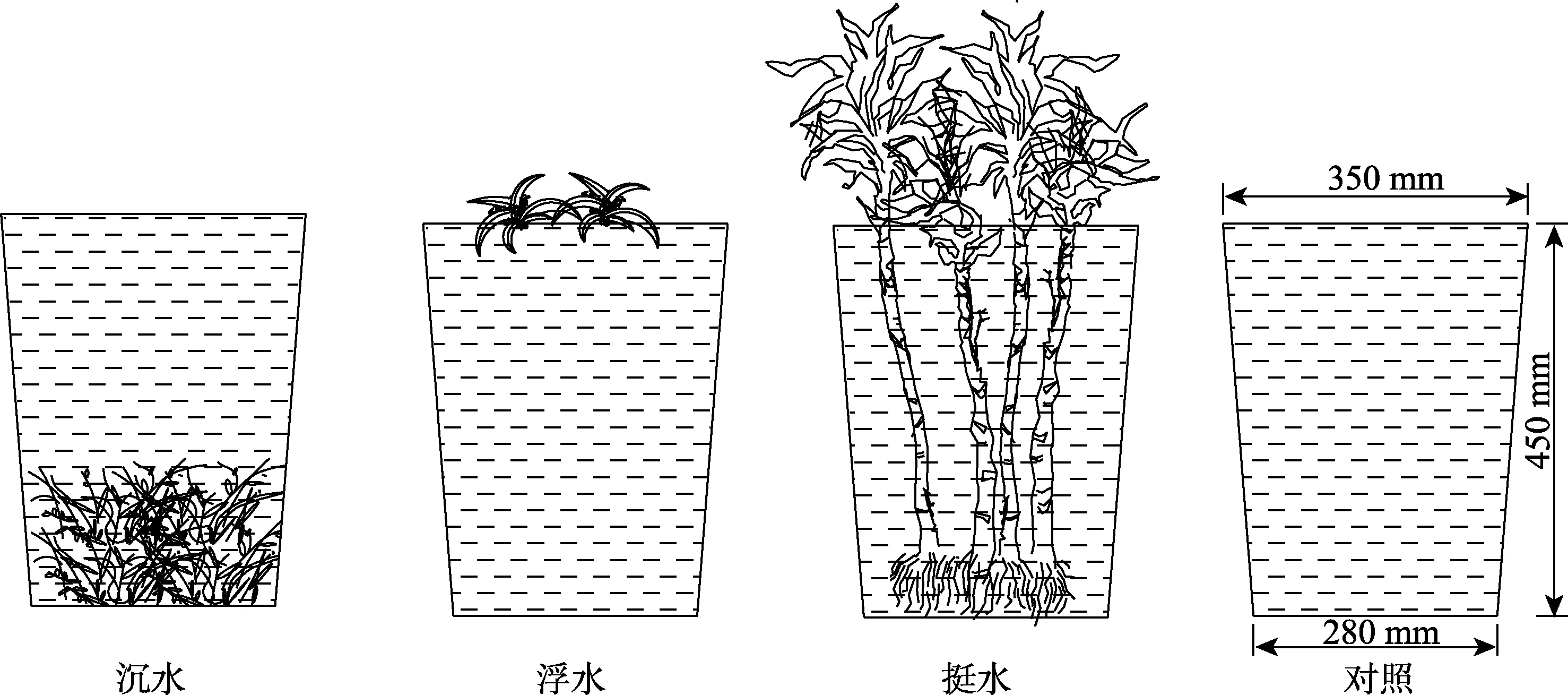
图3 污水试验装置
1.2.3 数据处理通过SPSS 22.0软件对数据进行单因素方差分析判断各因素对实验指标影响的显著性,采用Excel 2016和Origin 2021软件绘制图表。
2 结果与分析
2.1 不同跌水方式的增氧效果比较
如图4所示,3种跌水方式相对空白对照,都显著增加了水体中溶解氧含量(DO),表现为阶梯式>瀑布式>坡流式,增加的DO分别达到2.00、0.60、0.20 mg/L,相比空白组,分别增氧66.7%、20%、6.7%。
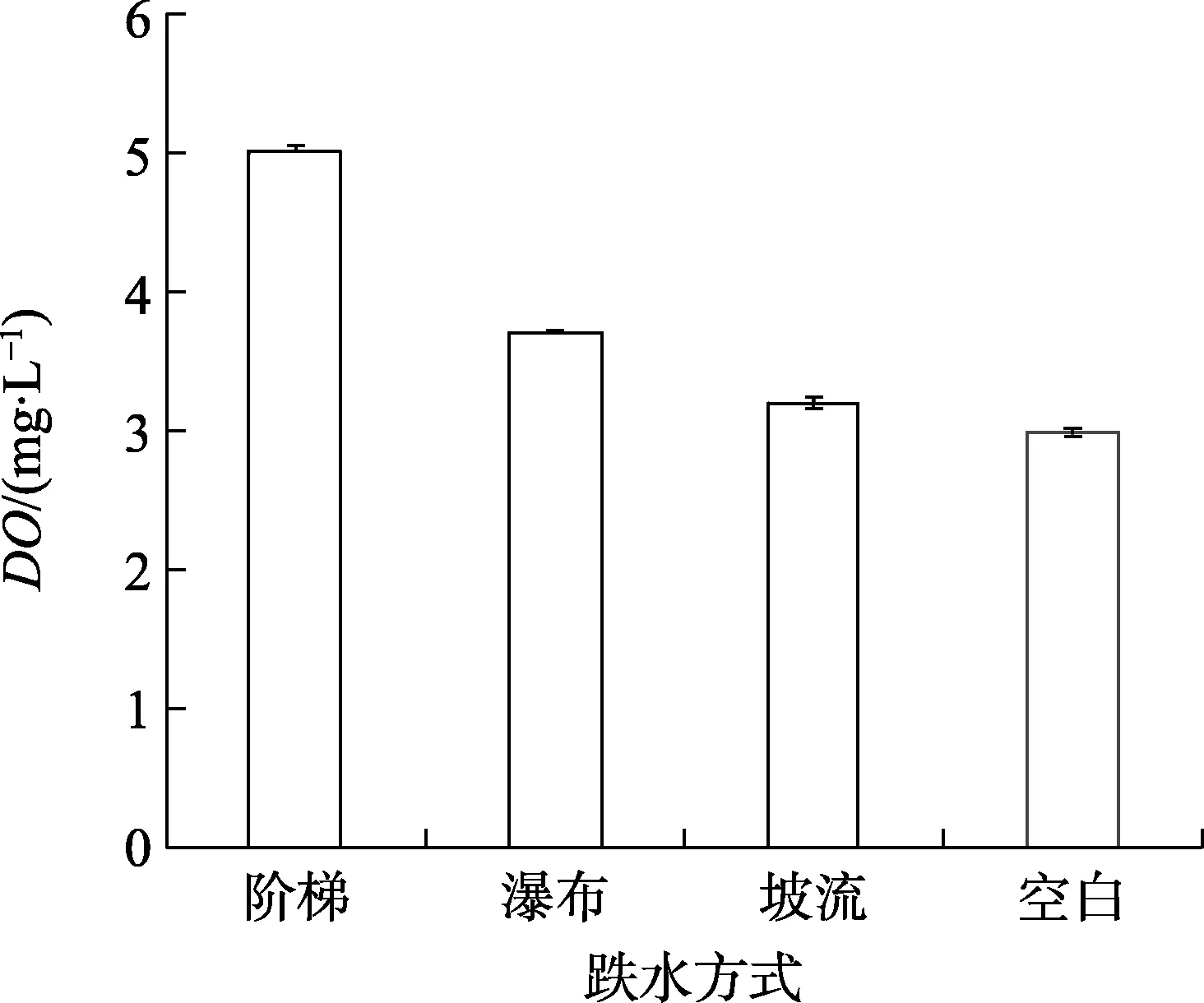
图4 不同跌水方式对溶解氧含量(DO)的影响
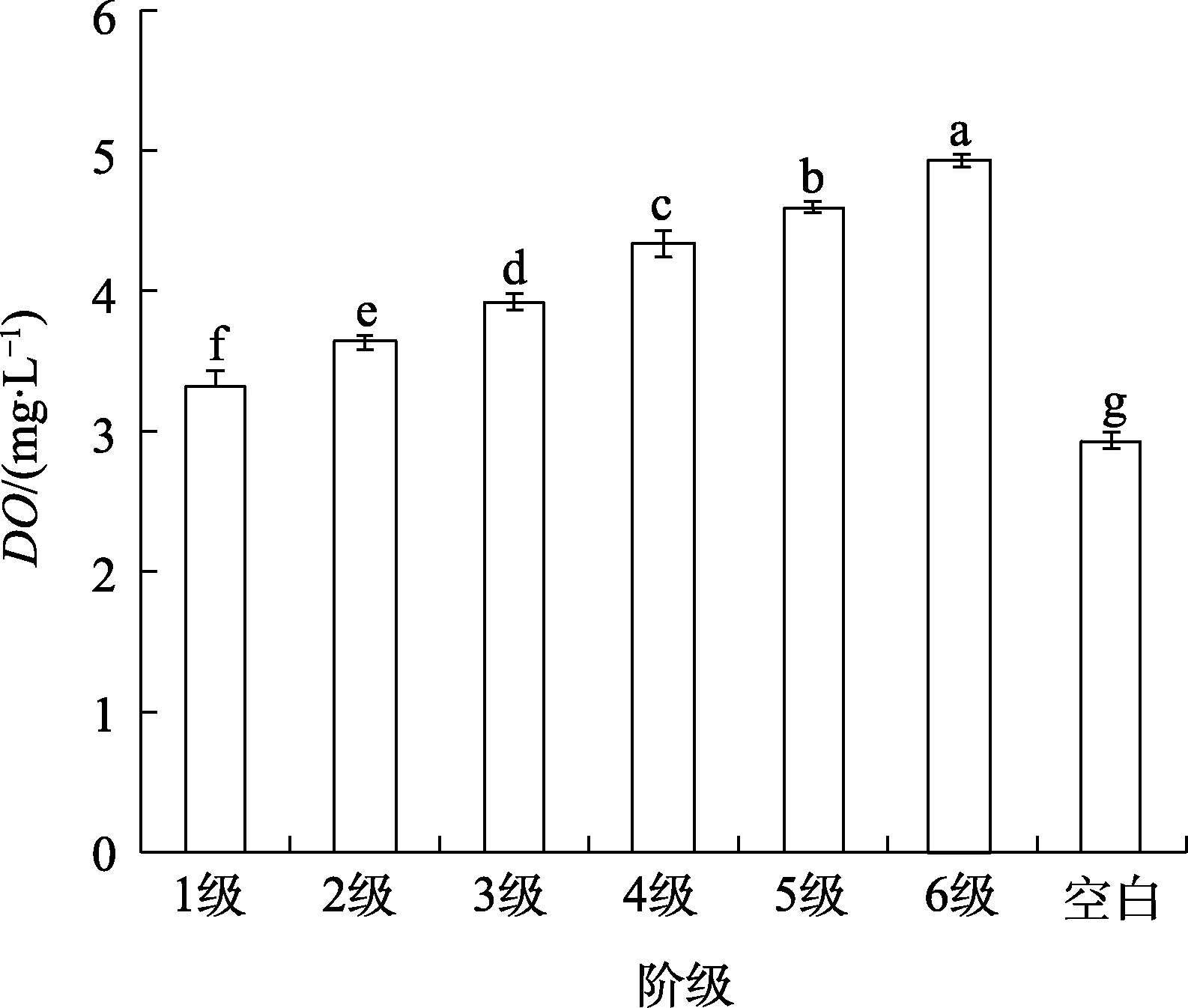
图5 不同台阶级数对溶解氧含量(DO)的影响
2.2 不同台阶级数跌水溶解氧增氧效果比较
如5图所示,在水体温度为30℃时,初始溶解氧含量(DO)为3.00 mg/L的污水分级沿着六级阶梯跌下,水中的DO逐渐升高,每级台阶的增加量为0.20~0.50 mg/L,每增加一级阶梯都显著增加DO;到最后一级阶梯上,DO已经达到了5.00 mg/L,比未经过跌水的空白组高出了2.00 mg/L。
2.3 无氧水中不同植物根系泌氧能力比较
2.3.1 沉水植物如图6所示,苦草的根系泌氧能力最强,最高溶解氧含量(DO)达到9.60 mg/L,接近于此温度(26℃)下的饱和浓度;金鱼藻次之,最高DO为7.00 mg/L,狐尾藻最低,只有4.30 mg/L。从试验时间8:00开始,种植苦草桶内的水体DO随着时间的延长和光照的增强而显著上升,到16:00达到一天内峰值。经过一个晚上,DO下降至3.00 mg/L,第二天8:00,DO又开始快速上升,在第二天16:00达到试验期内的峰值(9.60 mg/L)。苦草和金鱼藻的曲线趋势基本相同,而狐尾藻不一样。以种植狐尾藻为主的水体DO在第一天都有上升的趋势;到第二天,水中DO则基本不变。
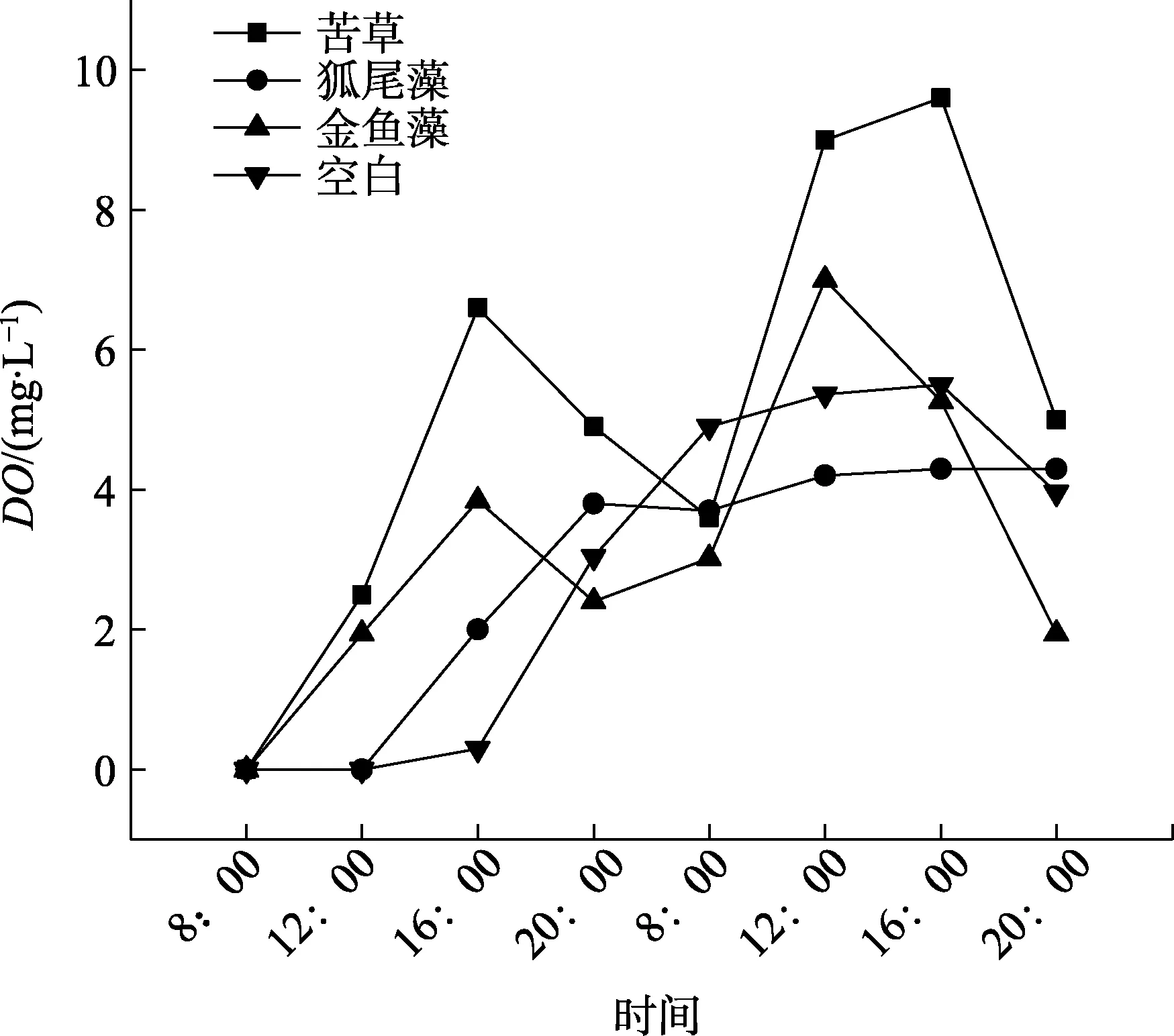
图6 沉水植物溶解氧含量(DO)随时间变化
2.3.2 浮水植物如图7所示,3种水生浮水植物中水体溶解氧含量(DO)表现为:水葫芦>睡莲>萍蓬草。随着时间变化趋势基本保持一致,在第一天12:00~16:00时,DO快速增加;16:00—20:00,增速缓慢,20:00之后则下降,第二天呈现相同趋势,并在16:00达到峰值。3种浮水植物中,水葫芦的根系泌氧能力最强,最高DO值为4.40 mg/L。
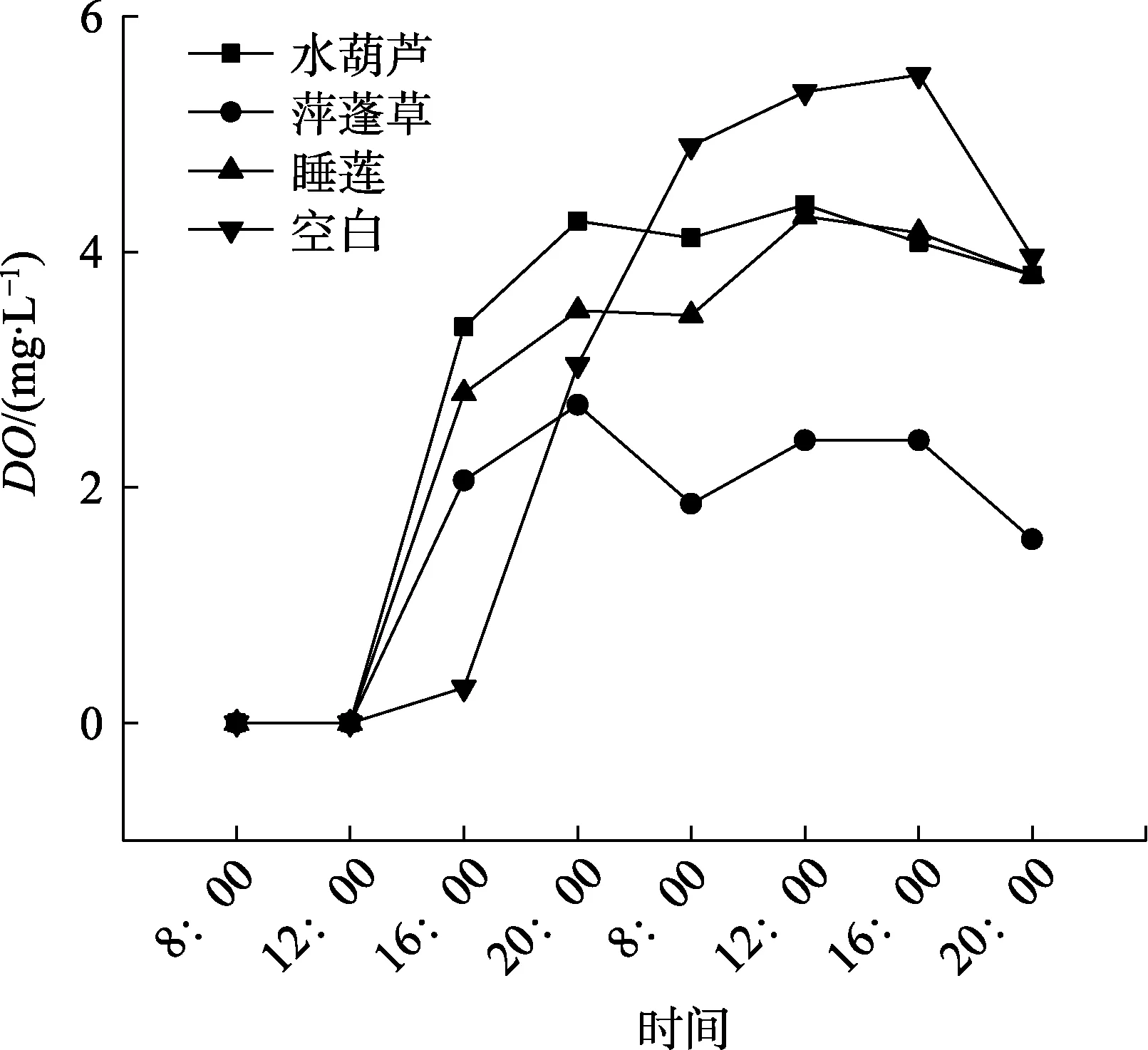
图7 浮水植物溶解氧含量(DO)随时间变化曲线
2.3.3 挺水植物如图8所示,千屈菜的根系泌氧能力最强,最高溶解氧含量(DO)达到5.92 mg/L;再力花次之,达到4.60 mg/L;黄花鸢尾最低,仅有3.40 mg/L。挺水植物的根系泌氧能力从强到弱依次为千屈菜>再力花>黄花鸢尾。第一天,随着时间的延长,所有挺水植物DO逐渐上升,第二天16:00到达峰值;16:00—20:00,DO有下降的趋势。第一天20:00到第二天16:00的变化曲线趋于平缓,DO值较稳定。
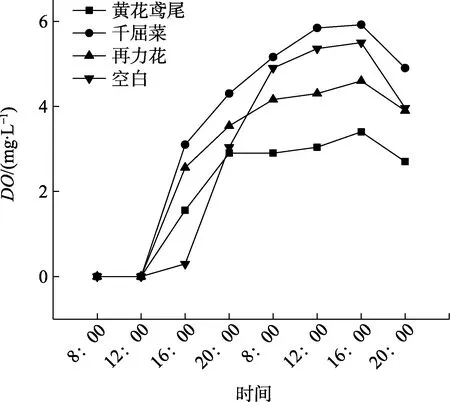
图8 挺水植物溶解氧含量(DO)随时间变化曲线
2.4 污水中不同植物根系泌氧能力比较
由图9可知,溶解氧含量(DO)9种植物水体中DO都显著少于空白组。其中,梭鱼草、再力花、花叶芦竹和水葫芦的水体DO较高,接近于空白组,为8.00~9.00 mg/L。风车草、美人蕉、金鱼藻和苦草大致相同,为6.00~7.00 mg/L。狐尾藻最低,仅有5.00 mg/L。水生植物对富营养液水体DO的影响从强到弱依次是:梭鱼草≈再力花≈水葫芦≈花叶芦竹>苦草≈美人蕉≈风车草>金鱼藻≈狐尾藻。
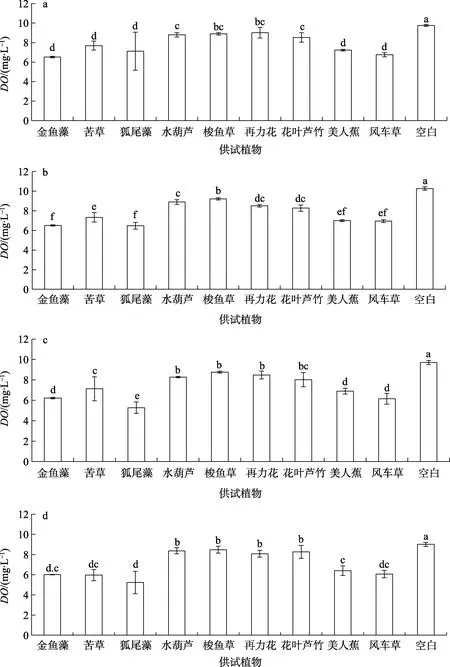
图9 不同时期下水生植物对溶解氧含量(DO)的影响
挺水植物组中,水体溶解氧含量组在20 d内基本维持在一个稳定的范围内,以美人蕉为例,水中的DO在6.80~7.20 mg/L浮动,无差异性(p>0.05)。而沉水植物组中,水体溶解氧含量不高,比挺水植物平均值低2.00 mg/L左右,随着时间的延长,DO不断减少。此外,未经处理的空白组DO随时间推移有所下降,从开始的9.80 mg/L到9.00 mg/L,下降了0.80 mg/L。
3 讨论与结论
3.1 讨论
本研究结果表明,不同跌水方式的增氧效果大小为:阶梯式>瀑布式>坡流式,对比图4、5发现:瀑布流跌水式的增氧能力接近于台阶跌水的第一和第二级,瀑布式跌水只有一次跌水碰撞过程,第一级阶梯式跌水同样经过一次跌水碰撞过程,高度仅有3 m的1/5,但增氧效果只差0.10 mg/L,说明跌水碰撞的氧气传质过程的次数和跌水高度共同影响跌水增氧效率。
每一级跌落的过程中,都伴随着一次水与水跌落碰撞的过程,这个过程水体发生扰动,增加了氧气与水面的接触面积,加快了氧气融入水中,形成溶解氧含量的过程[11]。阶梯式跌水共经过6次跌水碰撞过程,最后水体溶解氧含量增幅达到2.00 mg/L,而瀑布流和坡流方式,都只经历了一次跌水碰撞过程,因此,阶梯式跌水的增氧效率最好。瀑布式和坡流式虽然也可显著增加水体溶解氧含量,但效果不及阶梯式跌水。在相同水流量的情况下,与空气接触面大,则增氧程度高,液相和气相的交互面会发生物质的交换,即空气中的溶解氧含量会扩散到水体中,扩散的动力是浓度差[12-13]。
在台阶式跌水试验中,水体中的溶解氧含量始终未达到饱和状态(30℃,饱和溶解氧含量为8 mg/L),证明在溶解氧含量水平较低的状态下,多级重力跌水增氧方式具有显著的效果;但在高度为3 m的状态下,经过六级台阶,增氧不完全,说明此时溶解氧含量有增加的空间。此外,各级台阶具有显著性差异,说明台阶式的跌水增氧方式可以较好地增加水中的溶解氧含量,且每经过一次跌水碰撞,水中的溶解氧含量会显著上升。
从生态学宏观角度研究水生植物对溶解氧含量的影响发现,水生植物在富营养化水体内构造了一个生境,为好氧微生物、兼性厌氧微生物和厌氧微生物提供栖息环境[14-15]。在小型水生生态系统中,水生植物可以同化吸收水体中富含的N、P等物质,用于自身的生长发育,并转化为生物量,达到净化水质的效果,微生物特别是好氧微生物,可矿化降解水中有机物,达到净化水体的作用[16]。除水生植物和微生物对富营养水体具有净化作用外,植物根系和微生物的耦合作用是修复水体的重要机制,表现出不一样的去除污染物机制。在植物根际范围内,植物的根际分泌物能够为根际微生物提供碳源,影响根际微生物的群落结构及其对有机物的分解[17]。而根际微生物在植物的营养循环、根部环境的维持、有害污染物的降解以及植物的生长代谢方面发挥着重要的作用,二者相互作用,形成共生关系。
根据本研究分析得出,影响水体溶解氧含量的主要因素有以下几个:
第一,不同生活类型和不同种类植物对溶解氧含量影响效果不同,沉水植狐尾藻DO趋于稳定,可能是根系泌氧产生的氧气和整株植物呼吸作用在水里摄取的氧气达到了平衡。研究显示,浮水植物凤眼莲叶柄连在茎上,中下部有膨胀的类葫芦状的气囊,通气组织发达,并且叶片的叶面系数、叶绿素含量和光合速率均处于高水平状态,因此,凤眼莲的根系泌氧能力十分优越[18-19]
第二,温度对不同水生植物水体溶解氧含量的影响。首先体现在对水生植物的影响,温度下降,水生植物内各种生物酶活性也随之下降,从而影响整个植株的活性。以苦草为例的沉水植物[20-21],一般生长期为3—11月,10月为生物量最大时期,此时苦草成熟结果,种子落在底泥,等到次年3月,种子在水中逐渐萌芽生长。可见,在试验期内,沉水植物处于生长后期,受到低温胁迫,水体中植物枯落物增加,造成试验用水发生污染,水中溶解氧含量降低[22]。但以挺水植物为例的再力花和凤眼莲,在低温条件下,植物生长正常,可自然越冬。因此,相对于挺水植物和浮水植物,沉水植物受低温胁迫的影响更显著。
第三,光照强度对不同水生植物的水体溶解氧含量的影响。由图6~8可知,水生植物中水体溶解氧含量随着光照强度的变化而发生改变,上午的光照强度强,则水中溶解氧含量快速上升;下午的光照强度更强,但由于植物自身的午休现象,气孔关闭,虽然水中的溶解氧含量也在增大,但增大的速率比上午低;在傍晚或夜晚,处于弱光照或无光照时,水体的溶解氧含量下降。由此可以推测,当光照强度大时,植物的光合速率也增大,植物根系分泌氧的速率加快,水中溶解氧含量大幅度上升;当光照强度弱或无光照时,植物的光合速率为零,呼吸作用消耗氧气,水中的溶解氧含量降低。
第四,水中微生物对不同水生植物水体溶解氧含量的影响。从自配污水的试验来看,对照组的水体溶解氧含量均显著高于9个植物组,由此猜测,空白组在试验初期没有水生植物,水体中的微生物含量较少,因此溶解氧含量还处在一个比较高的水平;但随着时间的延长,水中的微生物增加,尤其厌氧微生物增加,水体的溶解氧含量逐渐下降,慢慢发展为腐化黑臭水体,空白组的水体也随时间推移而腐化,溶解氧含量在20 d内减少了0.80 mg/L。不同水生植物处理的水体溶解氧含量差异也十分显著,原因是水生植物构造的小型生态系统、水生植物的根系泌氧活动和微生物矿化分解共同对水体溶解氧含量产生影响[23-24]。水生植物根系泌氧增加水体溶解氧含量,在根区形成了一个“生物反应圈”,好氧微生物利用根系分泌出来的氧气对水中富含N的有机物进行氨化—硝化—反硝化作用,从而分解矿化含氮有机物,净化水质[25]。因此,用挺水植物5组数据进行推测,对氮、磷有机物去除率高的水生植物组,溶解氧含量低下;对氮、磷有机物去除率低的实验组,溶解氧含量反而高。
3.2 结论
(1)在跌水高度为3.0 m的条件下,不同跌水增氧方式的增氧效果差异显著(p<0.05),3种跌水增氧方式的增氧能力由强到弱的顺序为:阶梯式>瀑布式>坡流式,增氧率分别为66.7%、20.0%、6.7%。
(2)同一种类型水生植物不同物种对水体溶解氧含量的影响差异显著(p<0.05),在用自配无氧水开展的植物根系泌氧试验中,所研究的3种沉水植物由强到弱顺序为苦草>金鱼藻>狐尾藻;浮水水植物由强到弱顺序为水葫芦>睡莲>萍蓬草;挺水植物由强到弱顺序为千屈菜>再力花>黄花鸢尾。
(3)不同水生植物对水体溶解氧含量的影响具有较大差异,根系泌氧能力为沉水植物>挺水植物>浮水植物,水体中溶解氧含量受3种类型植物生理学特性、生长周期、光照强度、温度等因素影响。
将增氧效果最佳的阶梯式跌水方式和根系泌氧能力较强的湿生植物应用于公园封闭性水体净化的设施,可达到生态治理和景观美化的双重效果,对城市净化黑臭水体和构建美观水体具有重要意义。
