园林废弃物堆肥复配菌剂的研究进展
2023-09-19陈宏伟
钱 玮, 陈 新, 陈宏伟, 高 宇
(1.苏州科技大学化学与生命科学学院,江苏 苏州 215009;2.张家港市园林苗圃园林科技研究所,江苏 苏州 215600)
近年来,随着城市外扩步伐的不断推进,城市绿化也得以快速发展。园林废弃物作为重要的生物质原料,其体量逐年增加[1]。因此,如何高效实现废弃物的资源化与清洁生产已受到广泛关注[2]。目前对于废弃物堆肥化处理的研究,主要集中于外源微生物菌剂的研制,以期改善堆体微生物的丰富度与多样性,从而构建出更多的降解酶系来完成对物料的选择性破坏,使之成为可再生利用的资源[3-4]。 自20 世纪40 年代以来,我国彭家元与陈禹平就通过添加高温富集培养出的纤维分解菌,短时间内实现了物料腐熟,为我国农业积肥造肥开辟了新道路。 但早期此类菌剂微生物较为单一,功能菌株抗逆性不足且很难长时间维持较高活性,因此,以复配菌剂作为堆肥的外源添加物已逐渐成为近年来的主流趋势[5-6]。 如席北斗[7]等在生活垃圾中添加的复合菌群(固氮菌、酵母菌、纤维素分解菌等),可使堆体温度24 h 上升至68 ℃,15 d 内有机物下降33.3%,腐熟时间也同比缩短了18 d。 Hou T[8]等将耐热的硝化细菌和硫氧化细菌复合加至污水污泥中进行堆肥,从而提高了生境中氮硫代谢的关键酶活与元素转化。 Zheng W[9]等研究表明添加蚯蚓粪和生物炭使秸秆堆肥中不动杆菌相对丰度增加,细菌群落Shannon 和Pielou 指数分别增加了9.42%和9.33%,实际应用中黄瓜增产达46%。因此,为了园林废弃物的高效利用,从而深入复配菌剂的探究,促使其在堆肥过程中协同发挥出更为显著的效果具有重要意义。
1 复配菌剂的研制
园林废弃物是一种富含木质纤维的堆肥材料,其中大量的木质素与半纤维素牢固地结合在一起,将纤维素致密地包裹起来,从而严重影响了微生物的降解与外界氧化[10-11]。 同时,其强烈的季节性与空间异质性,也直接阻碍了园林废弃物专用菌系的研制。 因此,通过合适的方法构建出一组关于木质素纤维素降解能力强的复合菌系,已成为园林废弃物高温快速堆肥的关键所在。 目前复配菌剂的研制方法主要包括:
1.1 功能菌株的组合
功能菌株的组合是指研究者根据自己的实验意图,选择一些已知的菌株或从堆肥物料中筛选的功能菌进行优化组合,从而得到一组针对性较强的复合菌剂[12]。 目前常见的组合方式有:①根据菌株的不同功能进行复配,如何慧中[13]等通过3 株具有木质素降解能力的真菌与7 种营养菌进行组合,发现菌群间可以彼此协同、联合作用,既能实现对外界环境的快速适应,又可完成对木质素的高效降解;②利用统计学方法(正交设计、均匀设计等)重组菌剂,如尹爽[14]等通过正交试验对不同菌株的比例、发酵时间与温度进行探索,从而得出S-3 菌株与白腐菌等比混合而成的复合菌剂,在30 ℃条件下发酵17 d 的作用效果最佳,其对纤维素和木质素的降解率分别可达35.60%和37.41%;③根据自然状态下的原始比例配制菌剂,如李天枢[15]等将不同温度下所分离筛选的菌株按照自然比例进行增殖和复配,由此而来的菌剂升温迅速,且对碳、氮的固定效果皆优于市场所售菌剂。
1.2 直接筛选法
直接筛选法摆脱了复合菌剂以纯培养手段为基础的固有模式, 是一种通过对特定时期的土著微生物进行富集或连续传代驯化,使之保持原有的协同关系而对物料产生降解效果的研制方法。其主要操作包括:①扩大培养,如徐智[16]等将特定环境下的堆肥原料直接接种至液体培养基中震荡培养, 使其有效活菌数达到理想数量级的培养物作为内源复合菌剂。研究表明,接种该菌剂的物料升温迅速且腐熟效果明显优于外源菌剂。 ②连续传代驯化培养,如崔宗均[17-18]等将高温阶段的物料接种至含滤纸的蛋白胨纤维素培养液(PCS)中,50 ℃下静止培养。 当浸在培养液中的滤纸条刚要断裂时,将其转接至相同的新鲜培养液中,重复传代数次,最终得到一组纤维素分解能力强的复合菌系MC1。 该菌系4 d 内对稻草秸秆的降解率可达60%以上。 ③腐料回用,如宋修超[19]等将堆肥腐熟后的物料直接作为复合菌剂,并将其添加至新的堆肥处理中。 结果表明,腐料使得堆肥的高温期提前了20 d,且平均温度同比上升了5 ℃左右。 两种复配菌剂研制方法的比较见表1。

表1 两种复配菌剂研制方法的比较
2 复配菌剂在堆肥中的群落结构演替研究
2.1 宏基因组学的应用
微生物的群落结构是动态变化的,它们与环境变量息息相关,其中劣势菌株因生长限制而被相继淘汰。相反,其他菌株却因此优势代之,最终达到一个相对稳定的状态。 故在针对复合菌剂的研究过程中,有必要采取一种合适的分析方法,用以揭示堆肥各阶段的群落结构以及演替规律,为菌剂微生物的后续影响提供一定的理论依据。
传统的分析方法是建立在分离与培养的基础上,通过对纯种微生物的形态特征或显微观察来了解群落结构的分布情况[20]。该方法操作简单、廉价,但由于培养基和培养条件的限制,目前可通过此法鉴定的微生物仅占环境总量的0.1%~10%,这显然不足以描述自然条件下群落结构的真实状态[21]。
宏基因组学技术是一种不依赖于传统微生物的分离与培养,而是直接通过提取环境样品中所有微生物(可培养、不可培养的活菌以及未降解的死菌)完整的遗传学信息,并经过合适的载体将其转运至稳定表达的宿主细胞中,以实现对所需目的基因的筛选[22-23]。 同时,随着高通量测序与分子鉴定技术的飞速发展,使得宏基因组学在挖掘有效生物学信息的过程中具备更加准确、全面、可操作性等优势,从而充分保障了后续微生物的种属鉴定以及相应功能的预测, 全面阐释了各群落结构间的差异性或与环境因子间的关联程度。 如Nagata N[24]等利用宏基因组鸟枪法对胰腺导管癌(PDAC)患者的肠道和唾液进行了研究。结果发现,患者样本中的物种组成与相对丰度在疾病早期就已出现严重失调。进一步说明了PDAC 对肠道和口腔微生物的影响。Du C[25]等通过构建宏基因组文库对岱海湖沉积物中抗生素抗性基因(ARGs)和金属抗性基因(MRGs)的多样性进行了研究。 其中共鉴定分析出了16 种ARGs 类型(主要是杆菌肽、链霉素、四环素、β-内酰胺和磺胺的耐药基因)与15 种MRGs 类型(主要为多金属耐药基因),从而解释了湖泊生境中的多种抗性行为。
2.2 群落结构及演替规律
2.2.1 复配菌剂的影响
在微生物的群落组成与演替规律的探索过程中,宏基因组测序技术向研究者提供了更加全面准确的信息。研究表明,复合菌剂的施加对原始微生物的群落组成有着显著影响。其中真菌对环境变化的适应能力较弱,往往在物种组成的方面表现出比细菌更大的差异性[26]。而相比之下,细菌的多样性受接种的影响较小,整体呈现出比真菌更多的种类和数量,这是由于细菌可利用的资源广,遇复杂因素的抗性强[24]。 而在园林废弃物的堆肥过程中,菌剂的添加也同样对微生物的群落演替造成了影响。如夏金利[28]等通过对堆肥样本的PCA分析发现,各处理中真菌的群落结构因菌剂的施加而出现了明显差异,这种差异随时间的延长而逐渐减小,10 d 后几乎消除。但细菌群落却与此相反,在前期物料升温的过程中,像Sphingobacterium、Thermobifida 等具有木质纤维降解能力的优势菌属在各处理中基本呈现出相同的变化趋势,而中后期仅能在含菌剂的物料中被检测到。 同样,Liu[29]等通过高通量测序技术对园林废物堆体中的细菌多样性进行了全面分析,结果表明菌剂的施加改变了堆肥中期细菌的演替方向,在此阶段中各处理的多样性指数(Shannon)最大,物种差异最为明显,从而进一步验证了菌剂对生境微生物群落的影响。
2.2.2 不利因素的影响
总体而言,复合菌剂的施加赋予了微生物更高的能量代谢与物质降解效率[27,30]。 但是,堆肥系统中群落的组成通常也会受到许多不利因素的干扰而导致理想的演替方向出现偏差,最终致使所呈现出来的菌剂效果微乎其微,甚至完全失败[31]。 如Xi[32]等认为,菌剂与土著微生物之间的竞争关系将会引起某些功能菌株的衰退,从而导致优势种群的多样性降低、菌剂的作用效果减弱。 同样,Zhao[33]等指出物料中残留的抗生素也会抑制菌剂微生物的数量与活性,从而造成肥力的大量损失。 另外,更为关键的因素在于园林废弃物的本质是一种富含木质纤维素的堆肥材料。 其在未经任何处理的条件下,仅凭微生物的降解作用是很难打破木质素和半纤维素之间的屏障,而使纤维素从中释放出来并转化为可发酵利用的糖。 最终大部分功能菌株由于营养匮乏而导致种类和数量急剧减少,引起发酵失败[34]。 因此,研究者必须采取相应的措施消除不利因素的影响(见表2),以确保功能菌株能随着群落的正常演替而发挥积极作用。

表2 生境中不利因素的消除措施
2.2.3 温度变化的影响
高温好氧发酵是园林废弃物菌剂快速堆肥的典型特征。 在保证氧气含量充足的条件下,微生物群落将会随着温度的变化而不断进行自我调节与生态位的分离,以实现群落演替[12,29]。 而对于堆肥的不同阶段,功能菌株的种类和数量越多,木质纤维的降解酶系就越丰富,反应速率就越快[43]。因此在施加复合菌剂后,系统中微生物群落结构的变化趋势应尽量符合物料温度的整体走向,以确保每个阶段的生物活性都维持在一个较高的水平。
(1)升温阶段。 堆肥初期,微生物优先利用各种可溶性有机物(脂肪、单糖和碳水化合物)进行生长繁殖并释放热量,从而引起堆体温度的迅速上升。 在此阶段中,微生物的种类和数量极为丰富(细菌、放线菌、真菌),其中主要以中温、好氧型为优势菌群[43]。 虽然这些菌类都具备有机物的分解能力,但对于不同化合物的亲和程度却有所差异。 如在园林废弃物堆肥的升温阶段,主要的优势细菌为厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)、变形菌门(proteobacteria)等,这是因为它们能有效吸收糖类等易分解的物质[28-29,44]。 相比之下,真菌的多样性却远低于细菌,大致与放线菌相当。 其中主要包括子囊菌门(Ascomycota)、担子菌门(Basidiomycota)等,它们在降解纤维素、木质素方面表现出超强的效果[28,44]。
(2)高温阶段。当物料温度上升至50 ℃以上时,即可认为进入高温阶段[43,45]。 此时,生境中的物种多样性与丰富度发生了显著变化,它们整体朝着嗜热性的方向进行演替。 在此阶段中,嗜热性细菌和放线菌的代谢活动逐渐加剧,存在明显的优势种群地位。 而大多数真菌受高温抑制甚至死亡,仅部分耐高温的真菌(Thermomyces dupontii、Thermomyces lanuginosus、Thermophilic sporotrichum)开始大量增殖且对木质素、纤维素等复杂化合物表现出强烈的降解能力[46-47]。 而当温度继续升高至60 ℃以上时,生境中几乎所有的真菌完全停止活动,此时嗜热性细菌与放线菌(Acidothermus cellulolyticus、Bacillus subtilis、Rhodothermus marinus、Thermobifida fusca)占据主导地位,继而对一些难降解的有机化合物加以分解并开始腐殖质的形成[46,48]。
高温期是园林废弃实现快速堆肥并获取高品质腐料的重要阶段。 在此过程中,功能性微生物不仅代谢活跃,对木质纤维表现出超强的降解能力。 同时,高温等物理条件也促使了大分子复杂化合物的结构裂解,使得大量芳香类物质得以释放,为腐殖质的合成提供了更多前体,有利于后期生物途径(微生物合成、细胞自溶)与非生物途径(美拉德反应、多酚自缩合等)对腐殖质的积累[49]。因此有必要通过宏基因组学技术,全面获悉堆肥各阶段的群落演替规律,从而阶段性补足不同时期的功能性微生物(见表3)。 特别在高温阶段,通过添加合适比例的嗜热性微生物,这对有效解决园林废弃物堆肥周期长、降解不彻底的问题具有重要意义[35]。
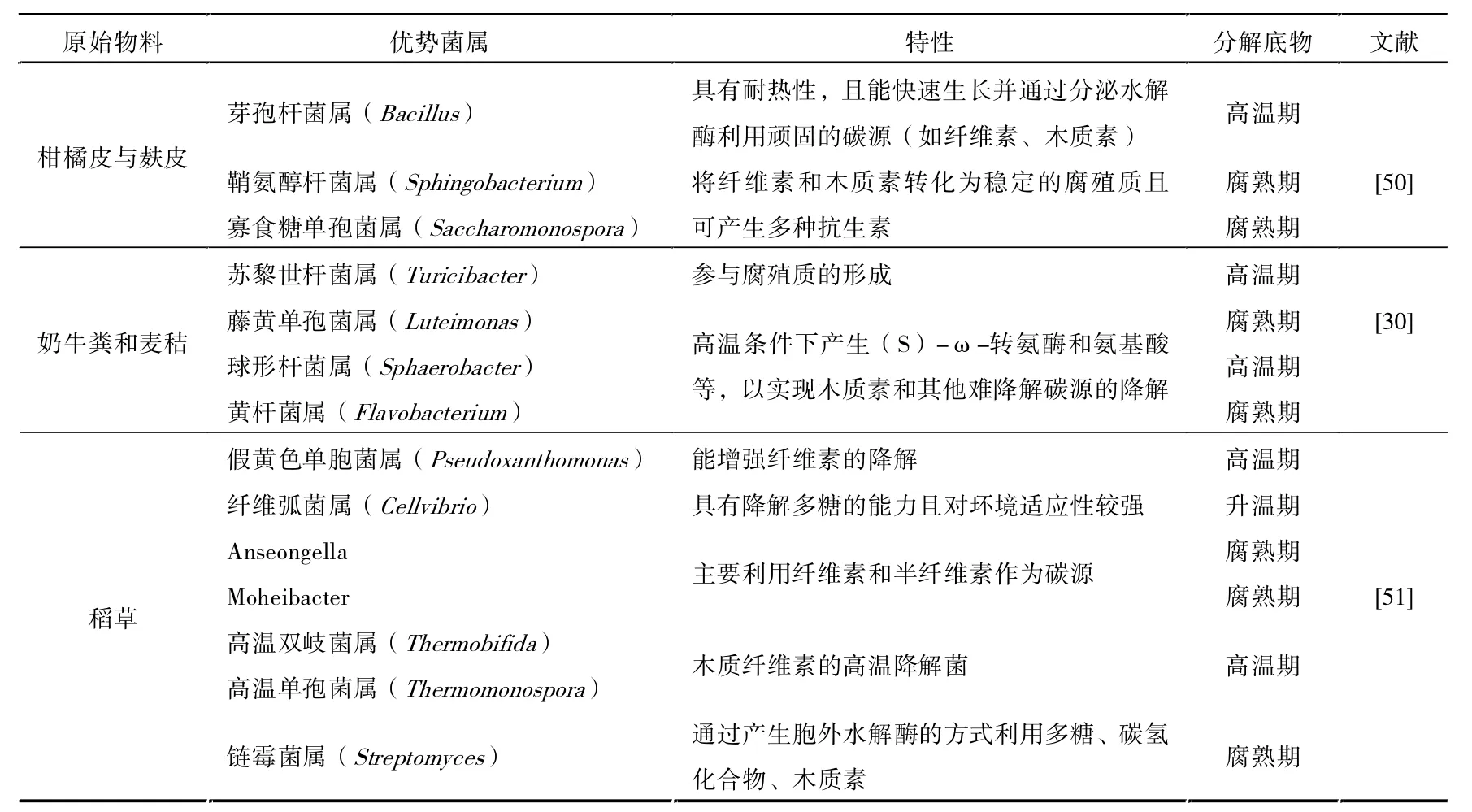
表3 不同物料中的功能微生物
(3)腐熟阶段。 当温度持续下降并稳定维持在40 ℃左右时,堆肥进入腐熟阶段[12]。 此时园林废弃物中可利用的有机物基本完成了矿质化和腐殖化, 体系中仅剩下一些较难分解的残留物以及新合成的腐殖质[3,15]。此时,由于营养的严重匮乏以及对温度环境的不适应,大量嗜热性微生物的多样性与丰富度开始逐渐降低。而那些经过高温期残留下来的中温细菌、真菌、放线菌又重新活跃起来并再次恢复优势地位,进而对上阶段的难降解物质继续分解、腐殖。
3 复配菌剂的群落结构的影响因素研究
如今,随着鉴定技术与数据处理能力的飞速发展,环境因素与物种之间的联系也越加明确。 研究者可通过对相关影响因子(如:原料含水率、C/N 比、氧气浓度、pH 值等)的定向调控,以确保复合菌剂的高效作用。
3.1 含水率
含水率是指示堆肥特性的一个重要参数,如果含水率过高,物料各组分间的空隙将会被水膜填充,导致堆体供氧不足而形成厌氧状态,并产生恶臭。 同样,过低的含水率也将严重阻碍微生物的生长繁殖、物质交换以及代谢速率,甚至导致整个堆肥工艺的失败[52]。目前大量研究表明,堆肥的起始含水率控制在50%~60%最为适宜[43,45]。 但由于园林废弃物中结构致密的木质纤维素对水分的持有能力较弱,往往会引起堆体底部积水,不利于好氧发酵[53]。因此还需通过翻堆或加入表面活性剂等措施加以改善[54]。
3.2 C/N
园林废弃物的C/N 比随组分、地域、季节而变化,整体呈现出高碳低氮的性质(见表4)。 理论情况下,微生物每消耗25 g 碳便需要吸收1 g 氮[43]。但当物料中的C/N 比过高时, 微生物则需要经过多次生命循环才能氧化掉多余的有机质,进而延缓了堆肥进程。 若C/N 比过低,则过量的氮素又将以氨气的形式流失,致使物料肥效下降。 通常来说,堆肥材料的初始C/N 比控制在25%~30%范围内最为适宜[12,55]。 因此对于园林废弃物的前期预处理,可以选择一些含氮量较高的经济来源物(见表5)进行调节。
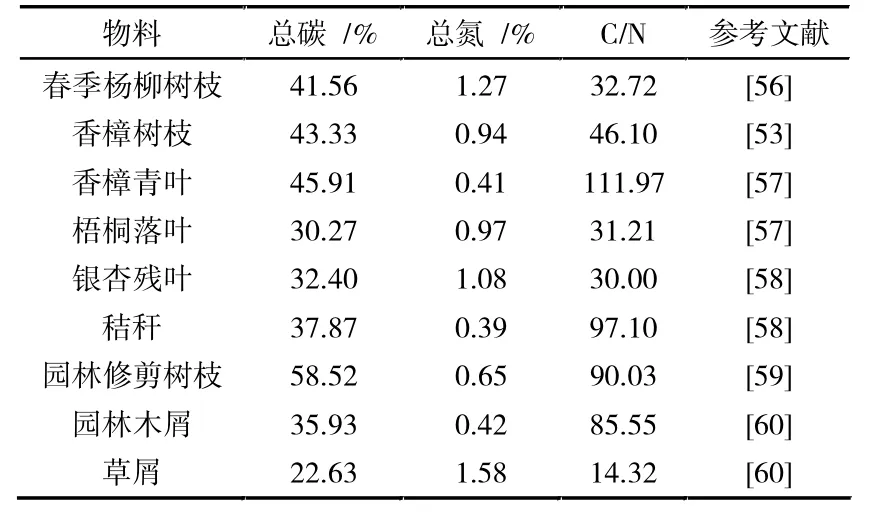
表4 常见园林废弃物的理化参数
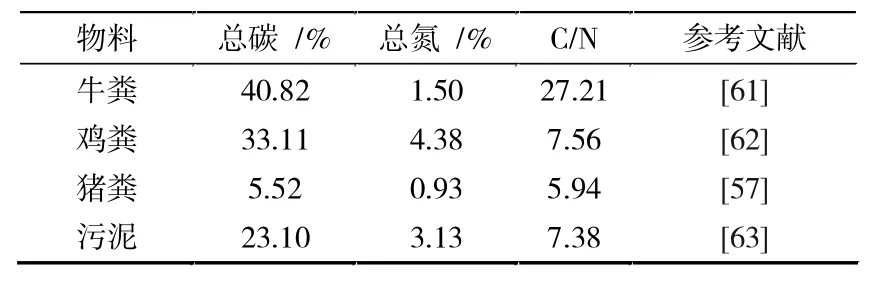
表5 常见高氮物料的理化参数
3.3 pH 值
pH 值是影响微生物代谢活动的重要因素之一,其大小随温度和时间的变化而变化。一般认为,中性或弱碱性为微生物堆肥发酵的最适环境[43,45]。当pH 值过高或过低时,可通过翻堆或加入一些中和药剂进行调节,以避免微生物的生长繁殖和有机物的降解受到抑制。 但由于堆肥系统本身存在一定的自我缓冲能力,故在实践操作中很少专门针对pH 值进行调控[52]。
3.4 氧气浓度
充足的氧气供给是实现园林废弃物高温快速堆肥的关键所在。 研究发现,堆体的溶氧浓度维持在8%~18%范围内最为适宜[64-65]。若通气太旺,则会引起微生物的活动加剧,致使营养物质以及水分严重流失。同时,热量的大量散失也将导致底物中的虫卵和病原菌无法有效杀灭;相反,若溶氧浓度低于8%时,堆体内部又将形成厌氧环境,从而产生恶臭并影响堆肥质量。 因此,研究者往往根据堆层中的氧气浓度和耗氧速率,以翻堆、表面自然扩散或强制通风等方式对系统中的溶氧浓度加以调控[52]。 但值得注意的是,在通风供氧的过程中,应充分考虑物料密度与空隙的最大饱和度,以避免造成温度的大幅度变化。
3.5 温度
温度是微生物活动强度的最好反映。 在堆肥初期,堆体中的嗜温菌较为活跃,能迅速利用物料中的可溶性有机物并释放出热量,引起温度的不断上升[66]。 当温度到达55 ℃左右时,堆肥进入高温好氧阶段。 此时嗜温菌的生长活动受到限制甚至死亡,而嗜热菌的繁殖进入激发态,一些复杂难分解的有机化合物(半纤维素、纤维素、木质素等)也开始逐渐降解[12]。 但高温不宜超过60 ℃,否则将导致大部分微生物的代谢活动停止,仅剩下部分嗜热菌[52]。 因此,一般认为高温阶段最好在55~60 ℃范围内,并维持3 d 以上,这样既能保证堆体微生物的丰富度和多样性,又能达到无害化处理的标准[52,67]。
4 结语
添加外源复配菌剂的堆肥化处理因其绿色、环保、经济效益大等显著优势,现已广泛实践应用于园林废弃物的资源化再利用。 但由于物料自身高C/N 比、亲水性差、透气性不足、高分子化合物(如纤维素、木质素等)难降解等特性,这对未来复配菌剂的模型构建、特性优化以及相关配套设备与工艺提出了更高的要求。相信随着研究的深入和产业化应用,复合堆肥菌剂能够为园林废弃物的科学化、绿色化、安全化处理提供强有力的支持,能够有效改善园林废弃物堆肥效率低的问题。
