黄土高原草地和刺槐根际AM真菌对刺槐和侧柏生长、土壤性质及叶片光合特性的影响
2023-09-14段文艳景若楠
段文艳,李 鑫,李 晴,景若楠,盛 敏
( 西北农林科技大学 林学院,陕西杨凌 712100)
土壤微生物趋于生境选择,生境不同其群落结构亦不同[1-2]。土壤微生物作为连接地上和地下生态系统的重要枢纽,其群落结构的变异对地上植被生长和地下土壤养分循环均具有重要影响[3-4]。丛枝菌根(Arbuscular Mycorrhizal, AM)真菌是重要的土壤生物成员之一[5],能与90%以上的陆生植物形成菌根,在促进植物生长、提高植物抗逆性、改善土壤肥力等方面发挥着重要作用[6-9]。已有研究表明AM真菌群落结构与生境条件(如宿主植物种类、土壤因子和环境因子等)关系密切[10]。对菌根真菌而言,原始生境决定了菌株特性,菌株来源不同其共生特性和抗逆性不同[11-12]。目前,AM真菌通过与植物根系侵染共生来促进植物光合作用、活化土壤养分、改善根系土壤微环境进而增强植物抗逆性的作用已被证实[13-14]。关于AM真菌的功能,目前多数研究仅关注一种或几种AM真菌的单一或简单复合效应[15-16],罕有研究探索AM真菌群落变异与其功能间的潜在关系。
黄土高原是中国水土流失最为严重的地区之一,为改善生态环境和防治土壤侵蚀,自20世纪50年代便于黄土高原开始营建了大面积的人工刺槐(Robiniapseudoacacia)纯林[17-18]。近年来,一些研究发现由刺槐单一树种组成的纯林可导致土壤退化、干化、生产力降低等诸多问题,限制了人工刺槐纯林的可持续经营和植被恢复后生态效益的发挥[19]。关于人工林生态系统稳定性差、功能和服务质量下降的原因,多数研究从环境因素[20]、林木特性[21]和人为因素[22]等方面进行了探索,而从微生物角度进行探索的研究报道较少[23]。Sheng 等[23]对黄土高原不同林龄刺槐纯林中AM真菌特性进行研究后发现,人工刺槐纯林对土壤中AM真菌孢子密度及其群落结构产生了显著影响,尤以35 a生刺槐纯林的影响最为突出。此外,刺槐和侧柏纯林控制黄土高原土壤侵蚀而导致的土壤退化被越来越多的研究者所关注,刺槐和侧柏等多树种混交林成为营造水土保持林的新趋势。Chen等[24]对黄土高原刺槐侧柏混交林进行研究后发现,刺槐的存在显著改变了侧柏植株的养分含量和AM真菌侵染特性。因此,本试验在此研究的基础之上,以采自草地(对照)和35 a生刺槐纯林地的土壤样品为菌剂,在盆栽条件下研究不同AM真菌群落对刺槐和侧柏植株生长状况、叶片光合特性以及土壤生物学特性的影响。研究结果可为揭示AM真菌群落变异与其功能间的关系以及“土壤-AM真菌-植物”的内在互作机制提供理论基础。
1 材料与方法
1.1 供试植物
供试植物为刺槐和侧柏,刺槐和侧柏种子均购于陕西省咸阳市杨凌金诺有限公司。选择大小一致的刺槐和侧柏种子,用5%的NaClO表面杀菌10 min,无菌蒸馏水冲洗5遍,将表面消毒后的刺槐和侧柏种子分别置于皿底铺有3层湿润滤纸和灭菌湿润河沙的无菌培养皿中,于28 ℃培养箱中催芽3~5 d,选择露白一致的种子移入盛有无菌育苗基质的育苗钵中,常规育苗管理30 d后选择生长一致的刺槐和侧柏幼苗,备用。
1.2 供试菌剂
供试菌剂为Sheng 等[23]研究中采集的草地土壤(简称草地菌剂)和35 a生刺槐林地土壤(简称刺槐菌剂),其土壤性质、AM真菌孢子密度及种属分布特征详见表1。供试菌剂的采样地点及采样方法简述如下。
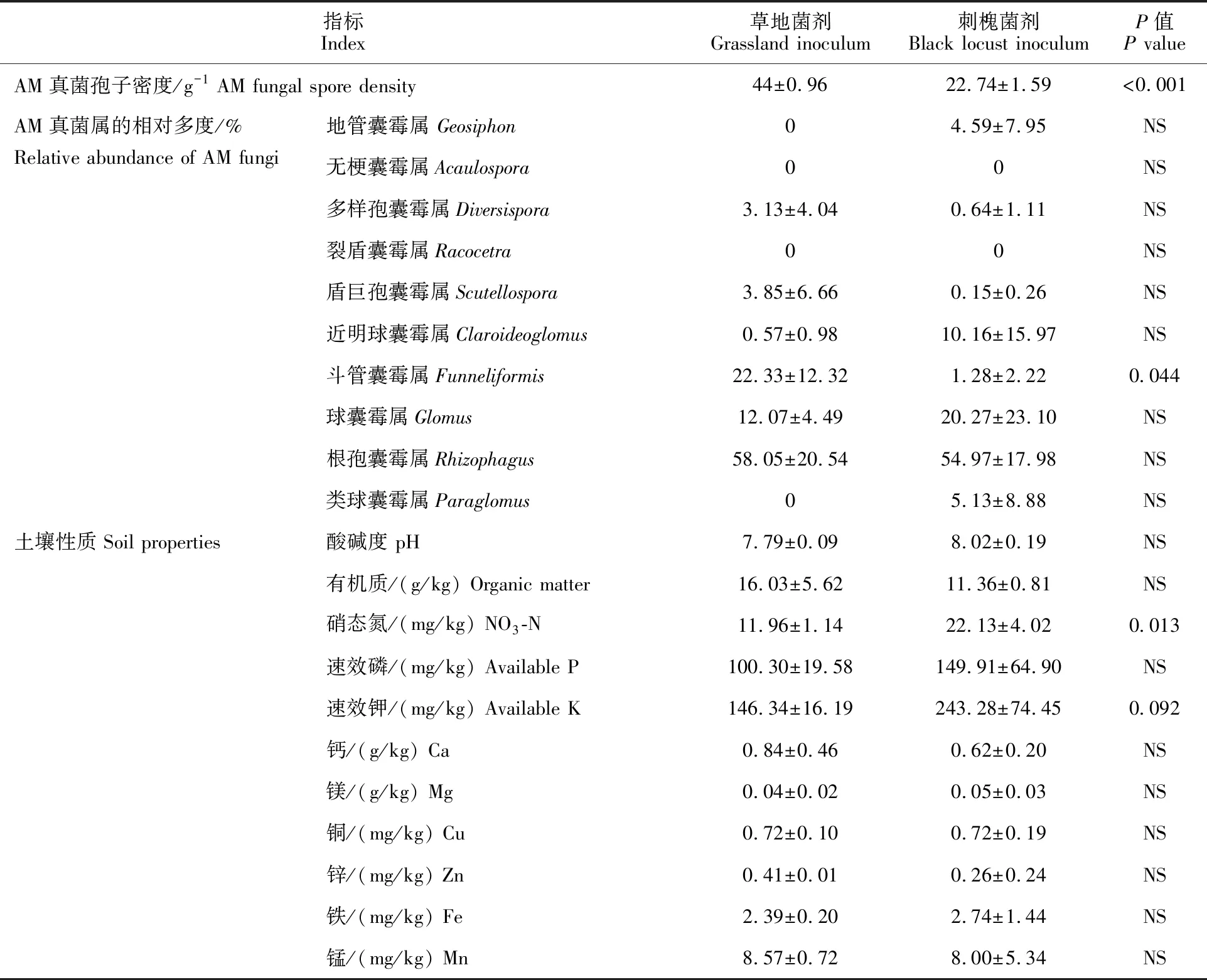
表1 供试菌剂的土壤性质、AM真菌的孢子密度及各属的相对多度
采样地点:所有供试菌剂均采自陕西省长武县中国科学院长武农业生态试验站的王东沟流域(35°12′N,107°40′E)。该流域地处黄土高原丘陵沟壑区,属暖温带半湿润大陆性季风气候,年均降水量584 mm,年均气温9.1 ℃,无霜期平均 171 d,土壤类型为黑垆土,母质为深厚的中壤质马兰黄土[25]。
采样方法:2013年11月,于王东沟流域选择坡向和坡度相似(阳坡,坡度约35°)的草地和35 a生人工刺槐纯林样地各1块,于各样地内分别设置3个20 m × 20 m 的样方;在各草地样方内,采用“S”形布点法在0~20 cm土层范围内采集土样约1 kg;在各刺槐林地样方内,随机选择刺槐5株,每株按东西南北4 个方位,去除枯枝落叶层后,在0~20 cm 土层范围内采集根围土约1 kg。
1.3 盆栽基质
2014年10月,再于2013年11月设置的草地样方内采集土壤样品作为本研究的盆栽基质;将所有盆栽基质过2 mm筛,高压湿热(121 ℃)灭菌2 h,取出放置一周,备用。
1.4 试验设计
以刺槐和侧柏作为供试植物,每种供试植物下设不接种AM真菌的对照(Non)、接种草地菌剂(GL)和接种刺槐菌剂(CH)3个处理,每处理重复6次。接种处理加入菌剂30 g/盆和50 mL菌剂过滤液,不接种处理施加等量灭菌菌剂和 50 mL菌剂过滤液。菌剂过滤液为草地菌剂、刺槐菌剂和水按1∶1∶10的体积比充分混合后利用双层滤纸过滤所得;菌剂过滤液的加入可保证各处理除AM真菌以外的其他微生物区系基本保持一致。
供试盆钵为150 mm×130 mm×150 mm的塑料盆(使用前先用0.1% KMnO4溶液浸泡2 h后,再用水冲洗、晾干后备用),每盆装盆栽基质2.0 kg,每盆种植刺槐或侧柏幼苗1棵。常规育苗管理6个月后,测定刺槐和侧柏的株高、地径、地上和地下部干质量、叶绿素相对含量、叶片光合气体交换参数、叶绿素荧光参数、土壤酶的活性、土壤中球囊酶素的含量、微生物量C、微生物量N以及根系的菌根侵染率。
1.5 测定指标及方法
1.5.1 株高、地径和生物量 株高和地径分别用直尺(cm)和游标卡尺(mm)进行测定。
地上和地下部干质量的测定:用自来水将植株地上(茎和叶)和地下(根系)部冲洗干净,再用蒸馏水清洗3遍,然后用吸水纸吸干表面水分后迅速放入烘箱110 ℃杀青30 min,再于80 ℃下烘干至恒质量,冷却至室温后称量。
1.5.2 AM真菌侵染率 参考Phillips等[26]的方法测定刺槐和侧柏根系的菌丝侵染率、泡囊侵染率和丛枝侵染率。
1.5.3 土壤酶活、微生物量及球囊霉素含量 采用高锰酸钾滴定法测定过氧化氢酶的活性[CAT,mg/(g·min)];采用3,5-二硝基水杨酸(DNS)比色法测定蔗糖酶的活性[SUC, mg/(mL·h)];采用苯酚钠-次氯酸钠比色法测定脲酶的活性[URE,μmol/(m2·s)];采用磷酸苯二钠比色法测定碱性磷酸酶的活性[ALP,mg/(mL·h)][27];采用氯仿熏蒸提取法测定土壤微生物量C(MBC,mg/kg)和微生物量N(MBN,mg/kg)的含量[28];采用Wright等[29]的方法测定易提取球囊霉素(EEG,mg/g)和总球囊霉素(TG,mg/g)的含量。
1.5.4 叶片光合特性 光合气体交换参数:利用Li-6400便携式光合作用测定系统(Li-Cor, USA)于9:00-11:30测定刺槐和侧柏自上而下数第2片完全展开叶的净光合速率[Pn,μmol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]、气孔导度[Gs,mol/(m2·s)]和胞间CO2浓度[Ci,μmol/mol)]。测定时使用6400-02B光源控制光强为1 000 μmol/(m2·s),CO2浓度为400 mol/mol,空气流速为0.5 dm3/min,叶室温度为25±1 ℃。每个测试点稳定2 min后读数,每个叶片连续记录3次,取平均值[30]。
叶绿素相对含量:利用CM-1000叶绿素含量测量仪(Spectrum, USA)测定刺槐和侧柏叶片的叶绿素相对含量(SPAD值),其测定时间与光合气体交换参数的测定时间同步。

PSII最大光能转化效率Fv/Fm= (Fm-Fo)/Fm
表观光合电子传递速率ETR = ΦPSII× PAR×0.5×0.84
其中PAR(photosynthetically active radiation)为光合有效辐射。
1.6 统计分析
采用R语言中的函数aov和duncan.test对植物生长状况、AM真菌侵染特性、根际土壤特性以及植物叶片光合特征分别进行单因素方差分析和多重比较;采用Vegan程序包中的函数adonis和rda分别进行多元方差分析(MANOVA)和主成分分析(PCA)揭示不同菌剂类型对刺槐和侧柏叶片光合特性的影响;采用Vegan程序包中的函数rda()和varpart()分别进行冗余度分析(RDA)和变差分析确定刺槐和侧柏生长状况的关键影响因素[34]。
2 结果与分析
2.1 AM真菌对刺槐和侧柏生长状况的影响
与Non刺槐植株相比,GL对刺槐植株生长无显著影响,而CH却显著降低了刺槐植株的地径、地上和地下部干质量;与Non侧柏植株相比,GL和CH均对侧柏植株具有显著的促生作用,且该促生作用在GL和CH处理间无显著差异(表2)。由此可知,不同AM真菌对刺槐和侧柏生长状况的影响亦不同。

表2 AM真菌对刺槐和侧柏生长状况的影响
2.2 AM真菌对刺槐和侧柏根系的侵染状况
分析结果表明,刺槐和侧柏根系的菌丝侵染率和泡囊侵染率在GL和CH处理间无显著差异,但其丛枝侵染率在GL和CH处理间具有显著差异(图1)。相对而言,GL更利于刺槐根系形成丛枝,而CH则更利于侧柏根系形成丛枝(图1)。

P<0.001:在0.001水平上差异显著。NS:差异不显著。下同
2.3 AM真菌对刺槐和侧柏根际土壤生物学特性的影响
研究发现,刺槐根际土中微生物量C、蔗糖酶和碱性磷酸酶活性、易提取球囊霉素和总提取球囊霉素含量在不同处理间无显著差异,而微生物量N、过氧化氢酶和脲酶活性则具有显著差异。其中,GL和CH的微生物量N显著高于Non(图2-A),CH的过氧化氢酶活性显著高于GL和Non(图2-C),Non的脲酶活性显著高于GL(图2-E)。
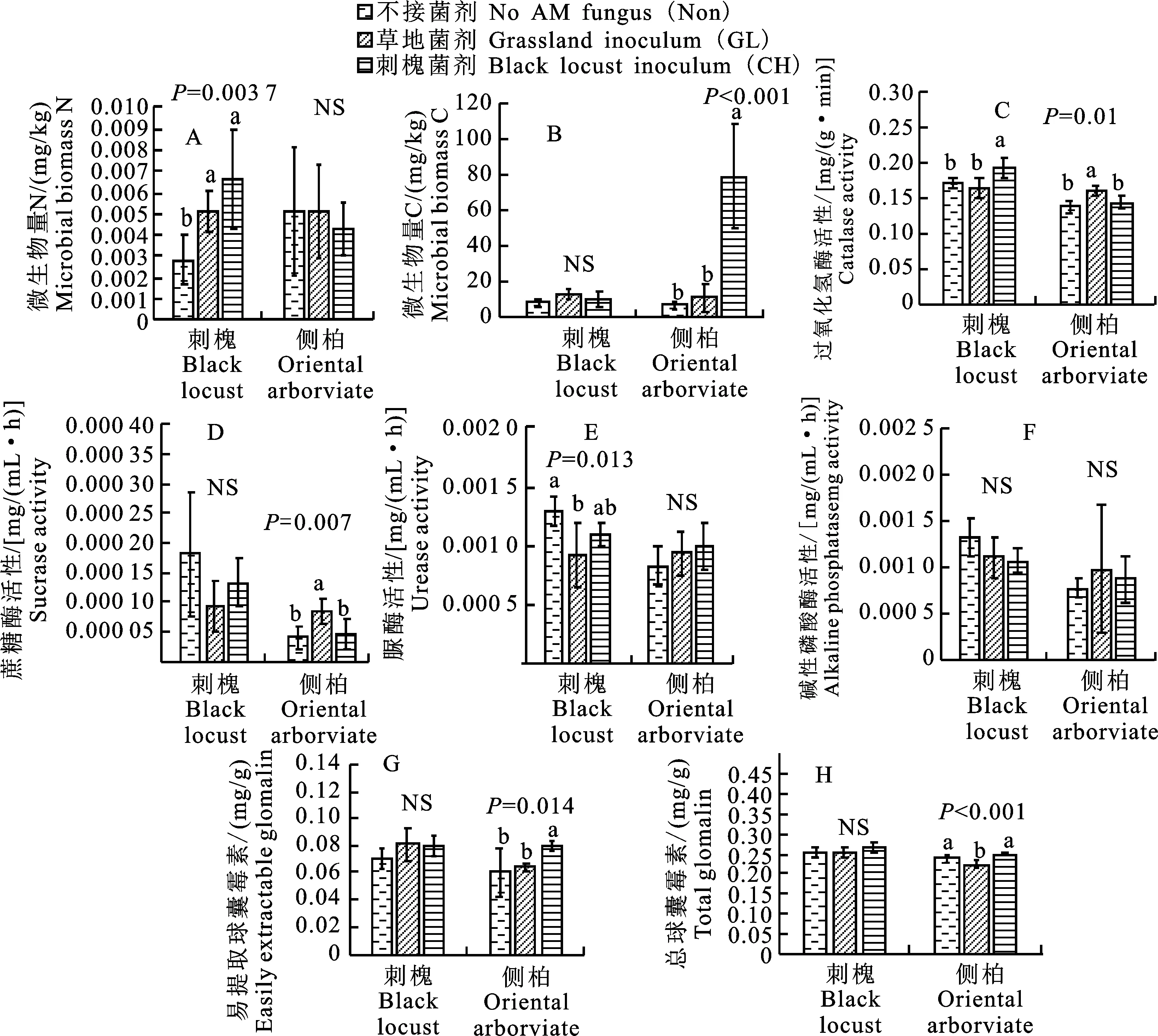
不同字母表示同一树种在不同处理间的差异显著(P<0.05);下同
研究还发现,侧柏根际土中微生物量N、脲酶和碱性磷酸酶活性在不同处理间无显著差异,而微生物量C、过氧化氢酶和蔗糖酶活性、易提取球囊霉素和总提取球囊霉素含量则具有显著差异。其中,CH的微生物量C和易提取球囊霉素的含量显著高于GL和Non(图2-B和图2-G),GL的过氧化氢酶和蔗糖酶活性显著高于CH和Non(图2-C和图2-D),GL的总提取球囊霉素含量显著低于CH和Non(图2-H)。
2.4 AM真菌对刺槐和侧柏叶片光合特性的影响
多元方差分析(MANOVA)结果表明,不同处理对刺槐(F=4.620,P=0.002)和侧柏(F=3.079,P=0.005)叶片光合特性具有显著影响。利用主成分分析方法(PCA)对刺槐和侧柏叶片光合特性进一步分析后发现,刺槐和侧柏的PCA1典型变量值在不同处理间具有显著差异(P刺槐=0.003,P侧柏=0.006)。其中,CH处理的刺槐叶片光合特性与GL和Non显著不同(图3-A);Non处理的侧柏叶片光合特性与CH和GL显著不同(图3-B)。

SPAD:叶绿素相对含量; Pn:净光合速率; Tr:蒸腾速率; Gs:气孔导度; Ci:胞间CO2浓度; Fv/Fm:PSII最大光能转化效率; ΦPSII:PSII电子传递效率; qP:光化学猝灭系数; NPQ:非光化学荧光猝灭系数; qN:非光化学猝灭系数; ETR:表观光合电子传递速率。下同
方差分析发现,刺槐叶片叶绿素相对含量、净光合速率、蒸腾速率、Fv/Fm、ΦPSII、NPQ、qN和 ETR在不同处理间具有显著差异,而其他光合特性指标则无显著差异。其中,Non的叶绿素相对含量显著低于CH和GL(图4-A);CH的净光合速率、蒸腾速率和Fv/Fm最高,Non次之,GL最低(图4-B、图4-E和图4-F);CH的ΦPSII和ETR最高,GL次之,Non最低(图4-G和图4-K);Non的NPQ和qN显著高于CH和GL(图4-H和图4-J)。

图4 AM真菌对刺槐和侧柏叶片光合特性的影响
方差分析还发现,侧柏叶片的净光合速率、气孔导度、胞间CO2浓度和Fv/Fm在不同处理间具有显著差异,而其他光合特性指标则无显著差异。其中,GL的净光合速率和气孔导度最高,CH次之,Non最低(图4-B和图4-C);Non的Fv/Fm显著低于CH和GL(图4-F);Non的胞间CO2浓度显著高于CH和GL(图4-D)。
2.5 刺槐生长状况的关键影响因素
选取受AM真菌处理显著影响的刺槐植株生长、菌根侵染、叶片光合特性及土壤生物学特性指标进行冗余度分析,分析结果表明丛枝侵染率、过氧化氢酶和脲酶活性、净光合速率、Fv/Fm、ΦPSII和ETR与刺槐植株生长状况(地径、地上和地下部干质量)显著相关。其中,丛枝侵染率与地下部生物量呈正相关,脲酶活性与地径和地上部干质量呈正相关,Fv/Fm与地下部生物量呈负相关,ΦPSII、ETR和过氧化氢酶活性与地径、地上和地下部生物量呈负相关。冗余度分析结果还表明,CH导致刺槐植株生长状况下降主要与其对刺槐根际土中过氧化氢酶活性、叶片净光合速率、叶绿素荧光参数Fv/Fm、ΦPSII和ETR的提升作用有关(图5-A)。

Above:地上部干质量; Under:地下部干质量; Dia:地径; Height:株高; CAT:过氧化氢酶活性; SUC:蔗糖酶活性; URE:脲酶活性; MBC:土壤微生物量C ; MBN:土壤微生物量N; EEG:易提取球囊霉素; TG:总球囊霉素; Arb:丛枝侵染率. ***表示在 0.001 水平下相关性显著; ** 表示在 0.01 水平下相关性显著;* 表示在 0.05 水平下相关性显著;. 表示在 0.1 水平下相关性显著
将与刺槐生长状况显著相关的土壤酶(过氧化氢酶和脲酶)、菌根侵染状况(丛枝侵染率)及叶片光合特性指标(净光合速率、Fv/Fm、ΦPSII和ETR)作为解释变量,利用varpart()变差分析函数比较土壤酶、菌根侵染状况及叶片光合特性对刺槐生长的贡献程度。结果表明,土壤酶、菌根侵染状况及叶片光合特性共解释了55.1%的变差,其中30.8%的变差为叶片光合特性单独解释,这表明刺槐植株生长状况的差异可能主要为叶片光合特性的变异所致(图6-A)。

图6 AM真菌、土壤性质和植株叶片光合特性对刺槐和侧柏生长状况影响的变差分析
2.6 侧柏生长状况的关键影响因素
选取受AM真菌处理显著影响的侧柏植株生长、菌根侵染、叶片光合特性及土壤生物学特性指标进行冗余度分析,分析结果表明微生物量C、过氧化氢酶活性、净光合速率、胞间CO2浓度以及Fv/Fm与侧柏植株生长状况(株高、地茎、地上和地下部干质量)显著相关。其中,侧柏植株各生长指标均与胞间CO2浓度呈负相关,与微生物量C、过氧化氢酶活性、净光合速率和Fv/Fm呈正相关。冗余度分析结果还表明,GL和CH显著促进侧柏植株生长主要与其对侧柏根际土中微生物量C和过氧化氢酶活性、叶片光合特性(净光合速率、胞间CO2浓度、Fv/Fm)的影响有关(图5-B)。
将与侧柏生长状况显著相关的土壤酶(过氧化氢酶)、微生物量C及叶片光合特性(净光合速率、胞间CO2浓度和Fv/Fm)指标作为解释变量,利用varpart()变差分析函数比较土壤酶、微生物量C及叶片光合特性对侧柏生长的贡献程度。结果表明,土壤酶、微生物量C及叶片光合特性共解释了61.3%的变差,其中叶片光合特性单独解释或与土壤酶、微生物量C共同解释的变差为52.6%,这表明侧柏叶片光合特性的变化可能是其植株生长状况差异的主导因素(图6-B)。
3 讨 论
3.1 不同AM真菌对刺槐和侧柏根系的侵染 能力
已有研究表明AM真菌的种属类别会对根系侵染特性产生不同影响[35]。本研究发现,刺槐和侧柏根系的丛枝侵染率在GL和CH处理间具有显著差异,而其菌丝侵染率和泡囊侵染率则无显著差异,这可能与草地菌剂和刺槐菌剂中AM真菌的种属组成有关。Knegt等[36]研究发现Rhizophagus侵染能力较强且能在宿主根内形成大量的菌丝和泡囊结构。而本文前期研究结果表明,草地菌剂和刺槐菌剂中Rhizophagus的相对丰度较高且二者间无显著差异(表1)[23]。因此Rhizophagus在草地菌剂和刺槐菌剂中分布的相似性可能是菌根化刺槐和侧柏根内菌丝和泡囊生长状况趋于一致的主要原因[37]。
Li 等[38]研究还发现Funneliformis可在刺槐根内形成大量的丛枝结构。本文前期研究结果表明,草地菌剂中Funneliformis的相对丰度显著高于刺槐菌剂[23]。草地菌剂中相对丰富的Funneliformis可能是菌根化刺槐根内丛枝侵染率较高的主要原因。然而,对侧柏而言,接种草地菌剂并未促进其根内丛枝的形成,究其原因可能与AM真菌的宿主特异性有关[39]。
3.2 不同AM真菌对刺槐和侧柏根际土壤生物学特性的影响
土壤微生物量可在一定程度上反映土壤微生物的数量、活性与功能[40]。本研究发现,在刺槐根际接种草地菌剂或刺槐菌剂均可显著增加根际土中微生物量N,这可能是因为刺槐对固氮微生物的富集作用和AM真菌对根瘤菌等根际微生物的促生作用使得土壤中微生物量N增加导致的[41]。而在侧柏根际仅接种刺槐菌剂可显著增加根际土中微生物量C。对于侧柏来说,草地菌剂或刺槐菌剂中的固氮微生物无法与侧柏根系作用形成根瘤,AM真菌成为影响侧柏生长的主要微生物,因此本研究中侧柏根际土壤微生物量N并未出现差异。研究中供试菌剂的AM真菌孢子密度为刺槐菌剂高于草地菌剂(表1),所以作为表征微生物数量的微生物量C其变化趋势与AM真菌孢子密度变化趋势相同,均为刺槐菌剂最大。
已有研究结果表明,AM真菌可显著影响宿主植物根际土中脲酶、过氧化氢酶和蔗糖酶的活性[42]。在土壤生态系统中,脲酶可将有机氮分解为无机氮供植物吸收利用,其活性的强弱常用于表征土壤氮素营养状况、土壤生产力及土壤质量等[43]。一些研究发现,接种Glomusmosseae或G.constrictum可显著增加刺槐根际土中脲酶的活性[44],而本研究却发现接种草地菌剂和刺槐菌剂均未显著增加刺槐或侧柏根际土中脲酶的活性。对于固氮植物刺槐来说,不接菌剂的处理中不存在AM真菌与固氮微生物共生提高土壤氮素这一途径,刺槐只能通过提高土壤中的脲酶活性来增加土壤氮素从而满足刺槐的生长发育,这可能是导致本研究中接种不同菌剂均未显著增加刺槐根际土中脲酶活性的主要原因。而非固氮植物侧柏对于氮素需求量少,脲酶活性也随之降低,因此在不同处理下的侧柏根际土中脲酶活性并未出现显著差异。过氧化氢酶可分解细胞产生的过氧化氢进而保护土壤微生物活细胞免受氧化胁迫,其活性的强弱可表征土壤肥力状况和土壤总的生物学活性[45]。已有研究表明接种AM真菌可提高植物根际土中过氧化氢酶的活性[46]。本研究亦发现在刺槐根际接种刺槐菌剂或在侧柏根际接种草地菌剂可显著增加过氧化氢酶的活性,过氧化氢酶活性的增加可在一定程度上缓解过氧化氢对土壤微生物和根系的毒害作用。蔗糖酶可分解土壤有机碳供植物和土壤微生物吸收利用[47-48]。本研究结果表明,在侧柏根际接种草地菌剂可显著增加蔗糖酶的活性,该酶活性的增加可加速侧柏根际有机碳的循环与利用,为侧柏生长提供碳源。综上,草地菌剂和刺槐菌剂对刺槐和侧柏根际土壤酶的活性均具有一定的影响,但其影响程度与宿主植物种类和AM真菌特性 有关。
球囊霉素是一种含金属离子的糖蛋白,主要存在于AM真菌菌丝体和孢子壁中[49],其常被认作是AM真菌对植物生长环境调整与适应的一种积极应答机制[50-51]。多数研究认为土壤中球囊霉素含量与AM 真菌种类及生长发育水平有关[52]。本研究发现,与不接种处理相比,接种刺槐菌剂可显著提高侧柏根际土中易提取球囊霉素的含量,球囊霉素含量的增加可能对维持侧柏根际土中有机碳平衡和土壤团聚体稳定性具有积极作用[53]。
3.3 AM真菌对刺槐和侧柏叶片光合特性的影响
已有研究表明,接种AM真菌在提升宿主植物叶片光合能力中发挥着重要作用[54]。叶绿素是植物叶片光合作用的载体,其相对含量可表征植株养分状况及叶片的光合能力[55];光合气体交换参数(净光合速率、蒸腾速率、气孔导度和胞间CO2浓度)可直接反映植株叶片的光合速率,叶绿素荧光参数(Fv/Fm、ΦPSII、ETR、NPQ和qN)则可为解释光合速率的变化提供依据[56]。具体而言,Fv/Fm表示光合中心吸收利用光能的最大效率,反应植物潜在的最大光合能力[57];ΦPSII反应实际光能转换效率,表征植物在稳态下的实际光合能力[58];ETR表示传递能量的效率与利用率[59];NPQ和qN 为 PSⅡ天线色素吸收的不能用于光合电子传递而以热形式耗散的光能部分[60]。本研究结果表明,对刺槐植株而言,接种刺槐菌剂提高了刺槐叶片叶绿素相对含量、净光合速率、蒸腾速率、Fv/Fm、ΦPSII和ETR,降低了叶片NPQ和qN,这一结果表明刺槐菌剂可能主要通过调节刺槐叶片的光合速率、PSII反应中心的反应速率和能量利用效率、光能的热耗散等来影响植株叶片的光合能力;对侧柏植株而言,接种草地菌剂和刺槐菌剂均能显著提高侧柏叶片净光合速率和Fv/Fm,降低叶片胞间CO2浓度,这一结果表明草地菌剂和刺槐菌剂可能主要通过调节侧柏叶片光合速率、光合中心吸收利用光能的效率及胞间CO2浓度来影响其叶片的光合能力。综上可知,刺槐菌剂和草地菌剂对刺槐和侧柏叶片光合能力均具有一定的影响,但其影响程度在不同菌剂间存在差异,究其原因可能与不同菌剂中AM真菌种属组成有关。
3.4 刺槐和侧柏生长状况的影响因素
土壤-植物-微生物互作是维持林地生态系统结构和功能的主要驱动力之一[61]。陈雪冬等[62]研究发现AM真菌-土壤-植物三者共同决定了黄土高原人工刺槐林的生长状况。本研究的冗余度分析结果表明,刺槐植株生长状况(地径、地上和地下部干质量)与丛枝侵染率、过氧化氢酶和脲酶活性、净光合速率、Fv/Fm、ΦPSII和ETR等因素有关,侧柏植株生长状况(株高、地茎、地上和地下部干质量)与微生物量C、过氧化氢酶活性、净光合速率、胞间CO2浓度以及Fv/Fm等因素有关;变差分析结果进一步表明刺槐和侧柏植株生长状况发生的变异均主要为其叶片的光合能力发生变异所致。
光合产物是植物生长发育的重要物质基础,光合作用的强弱可直接影响植物形态的建成[63-64]。已有研究发现,AM真菌作为广泛存在的土壤微生物对植物光合作用具有显著影响[65]。接种AM真菌能够显著提高植物叶片叶绿素的相对含量[66]、叶片光合气体交换参数净光合速率、蒸腾速率和气孔导度[67]以及叶片叶绿素荧光参数Fv/Fm、ΦPSII和ETR[68-69];同时可以降低胞间CO2浓度[70]以及叶片叶绿素荧光参数NPQ和qN[71]等。如前所述,本研究中也发现草地菌剂和刺槐菌剂对刺槐和侧柏叶片光合能力具有显著影响,且其影响程度在不同菌剂间存在显著差异,这一差异可能为菌根化刺槐和侧柏植株形态建成及生长状况发生变化的主导因素。
4 结 论
黄土高原草地和刺槐根际AM真菌的侵染能力及其对刺槐和侧柏生长状况、根际土壤生物学特性和叶片光合特性的影响均存在一定差异。AM真菌主要通过影响刺槐和侧柏的光合特性进一步导致植物生长出现了差异。就植物生长而言,草地AM真菌可促进侧柏生长,而对刺槐生长无显著影响;刺槐根际AM真菌可促进侧柏生长,而对刺槐生长则具有显著的抑制效应。这一结果表明草地和刺槐根际AM真菌群落差异以致其对宿主植物的促生功能发生了改变,且该促生功能的改变与宿主植物种类有关。综上可知,在黄土高原造林及刺槐纯林改造中做到“适地-适菌-适树”方能最大限度地发挥AM真菌的 功能。
