基于优化的最大熵模型预测中国高山栎组植物的历史、现状与未来分布变化
2023-09-11王晓帆段雨萱金露露王崇云彭明春王旭红马云飞
王晓帆,段雨萱,金露露,王崇云,*,彭明春,李 云,王旭红,马云飞
1 云南大学生态学与地植物学研究所,昆明 650504 2 云南大学生态与环境学院,昆明 650504
随着人类活动强度不断加大,全球气候变暖日愈加剧[1]。植物地理分布格局与气候密切联系,并且受到气候驱动[2—3]。气候变化已成为物种分布变化的一个日益重要的驱动因素,强烈影响物种的生态习性和地理分布、生态系统的结构组成和功能[4—5],从而导致生物多样性丧失和生态环境安全降低[6]。由于人类活动引起的气候变化的持续发酵可能将导致植物物种分布区减小甚至消失[7],因此,植物通常改变其生态分布来应对气候变化,例如向高纬度或高海拔地区迁移,以追随适宜的环境条件[8—9],否则,它们可能面临灭绝[10]。气候变化和人为干扰的威胁已经导致了种群的数量下降和灭绝风险。因此,研究不同时期气候变化背景下物种的潜在适生区分布格局变化趋势,对于制定合理的物种保护策略,实现生物多样性保护,维持人类赖以生存的环境尤其重要。
生态位模型(Ecological niche models,ENMs)可量化物种发生与各环境变量之间的相关性,从而描述某一物种的生态位或栖息地的适宜性[11],是一种用于解释气候等环境变量对于物种分布变化的有效工具[12]。生态位模型种类多,用途广。目前,生态位模型使用较多的有规则集遗传算法模型(GARP)[13]、生物气候分析和预测系统模型(BIOCLIM)[14]、随机森林模型(RFs)[15]、MaxEnt模型[16]等。在这些模型中,MaxEnt模型由于其建模直观、预测精度高、操作简便、解释力强等原因被国内外学者广泛应用在各个领域[17—18]。MaxEnt模型预测物种潜在适生区近年来不断增加,且广泛应用在不同的群落类型中。例如Liao等对中国西南地区的7种栎类植物的潜在分布以及驱动因素进行了研究,表明温度是影响高山栎组植物最重要的环境因子[19]。王琦等对中国弧隔鼠尾草亚属(唇形科)的分布格局进行研究,其结果表明降水量、年温变化范围和海拔是影响其分布的主要环境因子[20]。高健等基于MaxEnt模型对槭属鸡爪槭组物种在古气候、当前和未来气候空间分布格局及动态进行了研究,其结果表明鸡爪槭组过去时期呈现北扩,而未来则呈现两端收缩趋势[21]。Liu等基于MaxEnt模型对东北地区16种树种在气候变化下的潜在适生区进行了研究,结果表明日变化范围、年平均气温、气温年变化范围和低温热度是影响这些树种分布的主要影响因子[22]。MaxEnt模型取得了较好的预测效果,与实际分布区较为接近[23]。
高山栎组(Quercussect.Heterobalanus)植物属壳斗科(Fagaceae)栎属(QuercusL.)寒温性硬叶常绿阔叶树种,形成了分布于青藏高原东南缘和横断山地区典型的寒温性硬叶常绿阔叶林[24],集中分布于海拔2400—3600m陡峭的山峡等极端环境[25—26]。高山栎组植物的演化与扩散伴随着青藏高原的隆起抬升,其演化经历末次冰期等极端气候,是一种古地中海特殊的古老残余植被[27],具有重要的生态和经济价值,包括:①高山栎组植物具有“四素同构”的作用,高山上的水汽可在其作用下重新转换为雨雪,从而维持雪线的稳定,在青藏高原的冻圈生态学循环过程中有着重要作用[25,28];②对青藏高原地区的氧气供应、生物多样性维持、水土保持和碳储存等方面发挥着举足轻重的作用[29];③高山栎组树种极强的萌生能力,可作为煤炭的替代资源,保障了川藏农牧区几十年的薪材能源需求[28]。然而近年来由于气候变化和人类干扰加之缺乏可持续管理,该森林景观破坏严重呈现破碎化状态,形成林地分布具有大面积的线性边界,其分布范围及原生性生境正在不断缩小[30]。因此,高山栎组的演化、分布和扩散一直受到研究者的关注[31],包括系统发育[32]、群落特征[33]等方面,然而对于高山栎组植物在气候变化下的潜在分布鲜有报道。高山栎组植物作为古地中海古老植物,经历了漫长的演化历史,基于物种分布模型预测其潜在分布区空间变化格局及分析影响其地理分布的重要环境因子,对保护和抚育重建这一区域特有植被具有重要的科学意义。
不同气候背景不同时期物种的地理分布范围及变化趋势由于环境条件的差异,从而使得潜在分布区呈现出一定差异性。适生区预测模拟可揭示高山栎组植物未来变化趋势及主要的环境影响因子。因此研究基于优化的MaxEnt模型和ArcGIS软件模拟高山栎组植物从末次冰期以来即末次间冰期(the Last Interglacial,LIG)、末次盛冰期(the Last Glacial Maximum,LGM)、全新世中期(the Mid Holocene,MH)、当代(Current)、2041—2060(2050)和2061—2080(2070)年的地理分布和空间变化格局,以揭示影响高山栎组植物分布的关键环境因子以及变化特征,以期为高山栎组植物的资源保护、恢复及合理利用提供参考依据。
1 材料与方法
1.1 数据收集与处理
研究所需高山栎组植物的分布数据主要来源于全球生物多样性信息网络数据库(GBIF,https://www.gbif.org/)、中国数字植物标本馆(CVH,http://www.cvh.ac.an/class)等信息共享平台,结合野外实地群落调查的50个分布点,初步获得分布记录414条。以物种名、经度、纬度的格式整理为.csv格式。为避免物种分布点距离过近而引起的空间自相关影响,研究基于SDMtools工具箱在10km×10km范围内只保留一个分布点的原则,最终得到160个高山栎组植物有效分布点(图1),保存为.csv格式用于MaxEnt模型构建。
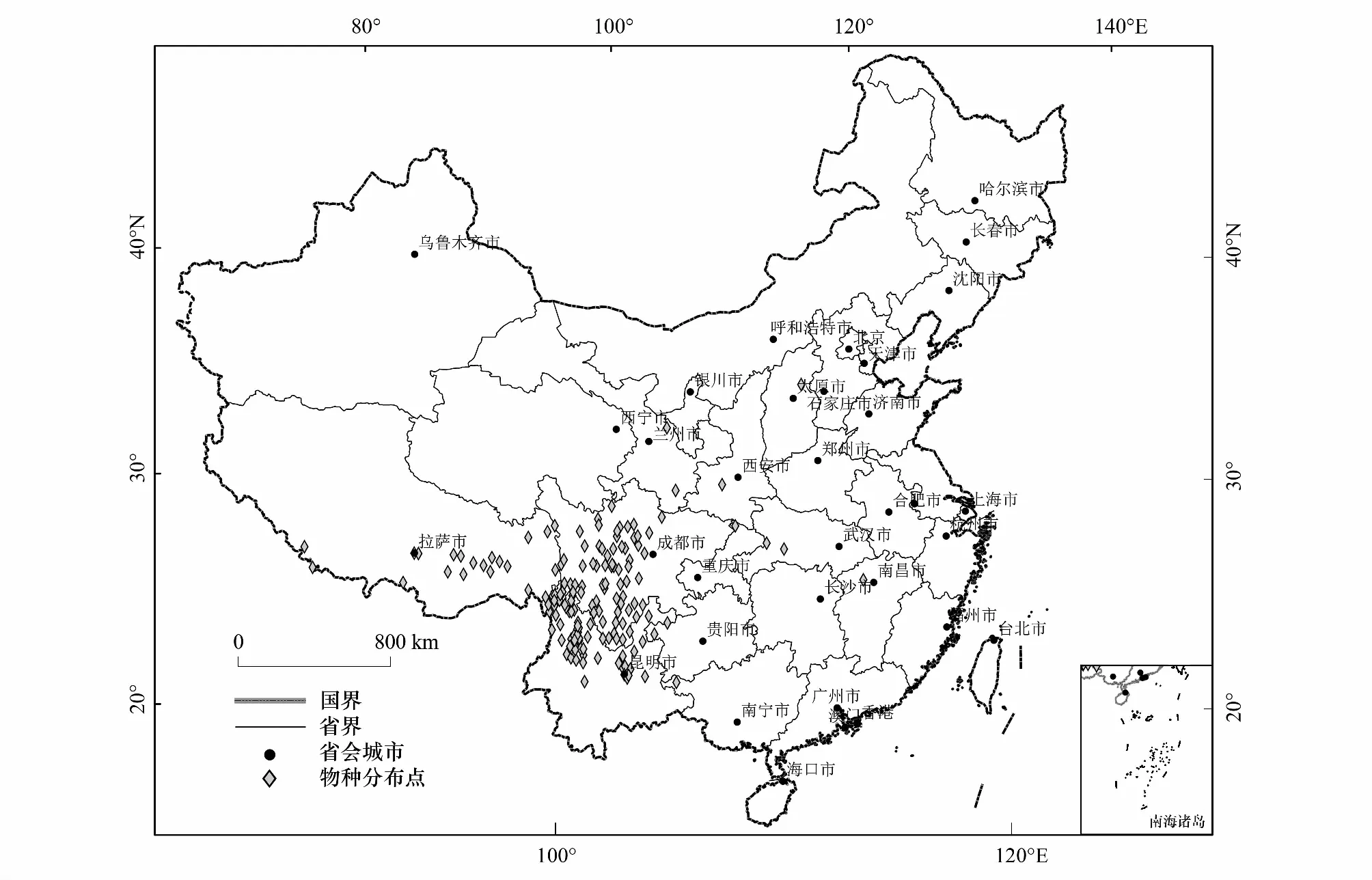
图1 筛选后的高山栎组分布记录Fig.1 Distribution points of Quercus sect. Heterobalanus after data cleaning
1.2 环境数据的获取与筛选
研究基于22个环境因子在6个时期,即末次间冰期、末次盛冰期、全新世中期、当代、2050s(2041—2060)、2070s(2061—2080)分别预测其分布区格局。环境数据包括19个生物气候因子以及3个地形因子。19个生物气候因子下载于WorldClim数据库(http://worldclim.org)。未来气候数据选择模拟中国气候效果较好的CCSM4模型下典型浓度路径(Representative concentration pathways,RCPs)[34],选取当前主流的3种不同浓度的排放路径(RCP2.6、RCP4.5和RCP8.5)来预测[35],RCP2.6、RCP4.5和RCP8.5分别代表未来低、中、高3种温室气体浓度排放情景。高程数据来源于地理空间数据云(http://www.gscloud.cn),坡度和坡向均由高程(dem)经ArcGIS 10.7重采样取得,所有环境因子经ArcGIS统一处理分辨率为30″(约1km×1km)。为避免环境因子间多重共线性造成模型的过度拟合,研究进一步根据模型训练的贡献率及各环境数据在R(4.1.1)中做Spearman相关分析的结果[36],剔除相关系数高于0.8且贡献率小(表1)的环境因子[37],最终筛选出12个具有统计学和生物学意义的环境变量进行建模(表2)。
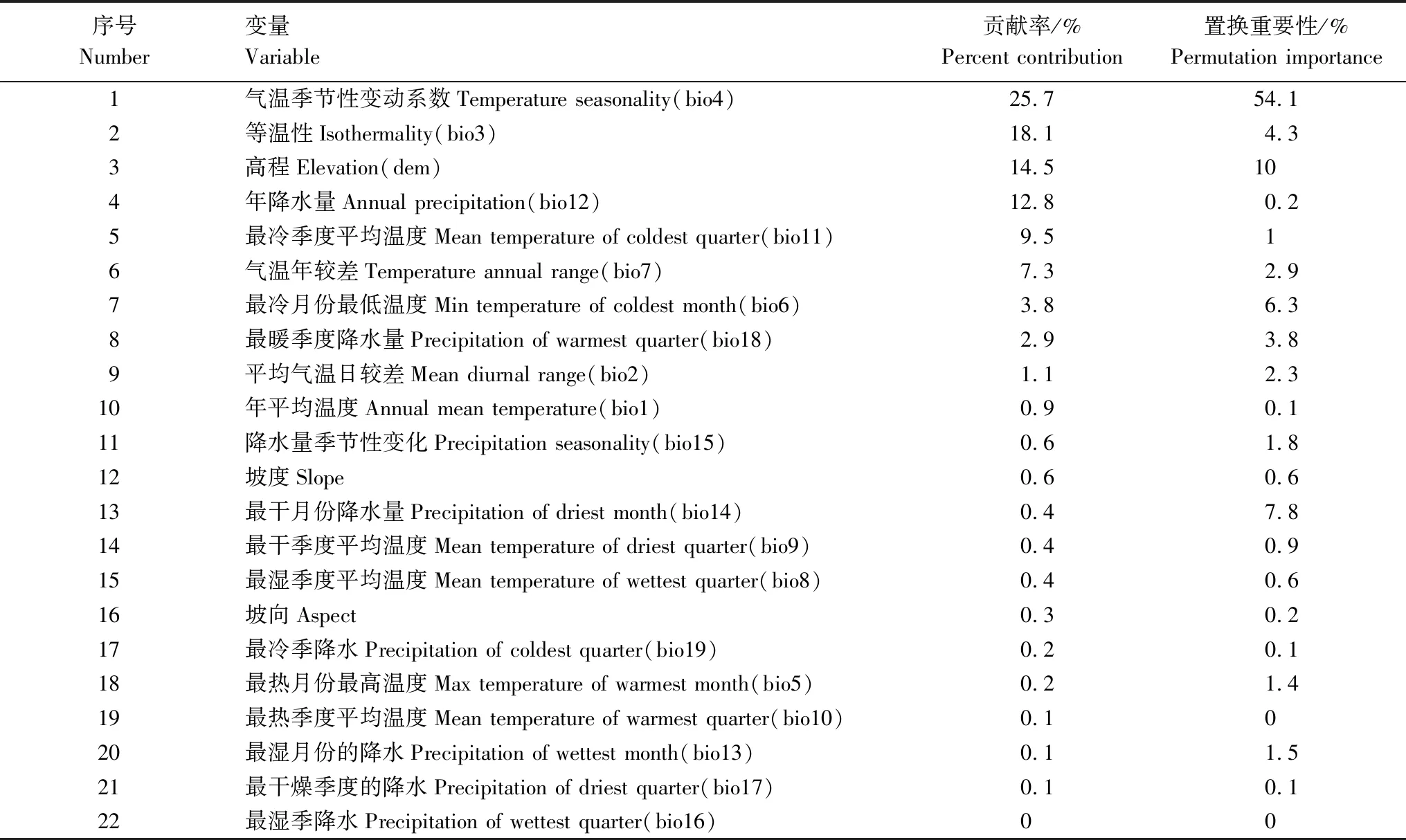
表1 环境变量对高山栎组分布的重要性
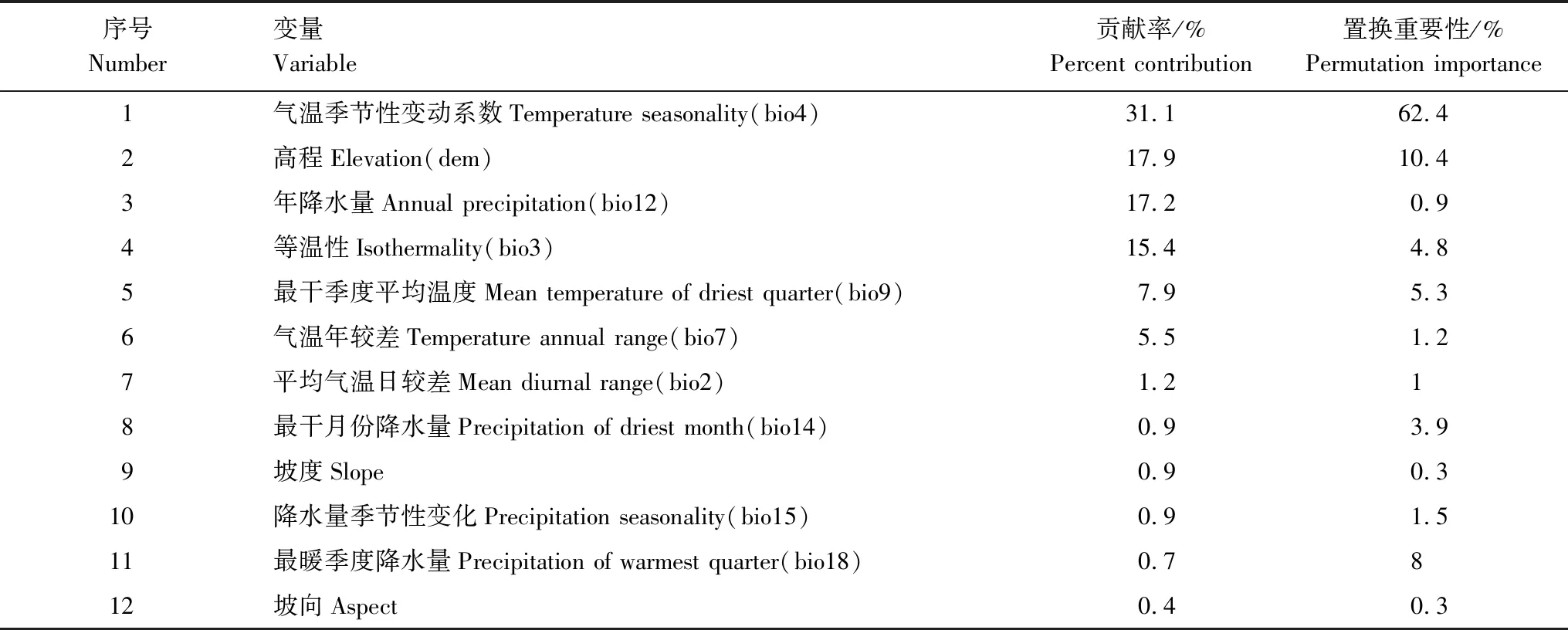
表2 MaxEnt模型各主导环境变量的重要性
1.3 模型建立与优化
研究运用ENMeval包(R4.1.1)对MaxEnt模型进行调优[38]。将范围为0.5—4、间隔为0.5的共8种调控倍频(Regularization multiplier,RM)与6个特征组合(Feature combination,FC),即:L、LQ、LQH、H、LQHP和LQHPT(L为线性、T为阈值性、Q为二次型、H为片段化、P为乘积型)进行参数之间的交叉组合[39]。之后在ENMeval包中将上述48种组合进行参数调测,最终选择delta.AICc=0,即最佳参数组合进行MaxEnt建模[40]。
1.4 模型评估
运用MaxEnt(V3.4.3)模型来预测高山栎组的潜在适生区分布。将当代分布点数据.csv格式和环境数据.asc格式文件导入软件,勾选刀切法,设置输出格式为Logistic形式,选取25%的分布数据作为模型的检测数据,75%的数据作为训练数据,勾选优化后的RM和FC值,重复10次,其他参数设为默认值进行建模。精度检验主要采取MaxEnt模型输出结果的受试者工作特征曲线(Receiver operating characteristic curve,ROC)下面积AUC值进行评估,AUC的取值范围为0—1,数值越大表示预测结果的可信度越高[41]。AUC值<0.7,预测结果差,0.7—0.8,预测结果一般,0.8—0.9,预测结果较好,>0.9,预测结果极好[42]。
1.5 适生区等级划分
将模型输出结果的平均值导入ArcGIS中,使用转换工具将其从.asc格式转换为Raster栅格数据。参考张明珠等在对长序榆(Ulmuselongata)研究中采用的分类标准[34],即采用平均逻辑值并结合实际分布区,将其适生区分为以下4个等级:P<0.2为不适生区、0.2≤P<0.4为低适生区、0.4≤P<0.6为中适生区、P≥0.6为高适生区,后统计不同适生区的面积。
1.6 物种适生区空间格局变化与质心分析
参考张殷波等的方法[43],在ArcGIS中将其适生区进行二值化,即分布概率<0.2的区域设定为不适生区,赋值为0;分布概率≥0.2区域设定为适生区,赋值为1,得到各个时期的非适生/适生二值图矩阵,定义0—0为不适生区,0—1为新增适生区,1—0为丧失适生区,1—1为保留适生区。计算不同适生等级下高山栎组植物在不同气候情景下与当代的面积变化、变化趋势与范围,获得其扩张、保留、收缩的面积以及地理范围。
基于上述二值图,运用SDMtoolbox工具箱计算模拟不同时期潜在适生区的几何中心位置变化,比较不同时期高山栎组核心适生区总体变迁趋势,反映不同时期环境变化对其分布的影响[44]。
2 结果与分析
2.1 模型优化结果及准确性评价
MaxEnt模型在模拟物种的潜在分布时会导致模型的过度拟合。研究调用ENMeval包基于物种分布点和环境数据在RM和FC的不同组合下进行交叉验证调优。由表3可以看出,当模型为默认参数,即RM=1,FC=LQHPT时,Dealt.AICc=135.164;当RM=2.5,FC=LQHPT时,Dealt.AICc=0,与默认参数相比,Avg.diff.AUC下降了约20.6%,Mean.OR10下降了约37.8%。表明当RM=2.5,FC=LQHPT时使得模型的复杂性和过拟合程度降低,模型精度更高。因此,研究选择RM=2.5,FC=LQHPT做为模型的参数进行建模。经参数优化后模拟得到受试者工作特征曲线下面积AUC平均值为0.960±0.015,表明模型预测结果可信度极高。

表3 MaxEnt模型ENMeval优化评价指标
2.2 影响高山栎组地理分布的主要气候因子
研究通过MaxEnt模型输出结果并基于刀切法的正则化训练增益贡献率、置换贡献率以及单因子响应曲线来揭示影响其生态地理分布的主导环境因子(表2、图2、图3)。从表2和图2中的贡献率和置换贡献率可以看出,气温季节性变动系数(bio4)、高程(dem)、年降水量(bio12)、等温性(bio3)的贡献率分别为31.1%、17.9%、17.2%、15.4%,累计贡献率高达81.6%,置换贡献率分别为62.4%、10.4%、0.9%和4.8%,累计达到了78.5%,表明以上4个环境因子是影响高山栎组植物分布的主导环境因子。
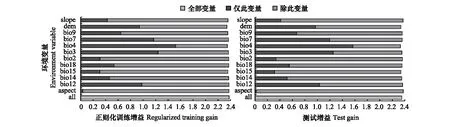
图2 主要环境因子的刀切法评价Fig.2 Evaluation of major environmental factors by Jackknife methodslope:坡度;dem:高程;bio9:最干季度平均温度;bio7:气温年较差;bio4:气温季节性变动系数;bio3:等温性;bio2:平均气温日较差;bio18:最暖季度降水量;bio15:降水量季节性变化;bio14:最干月份降水量;bio12:年降水量;aspect:坡向;all:全部变量
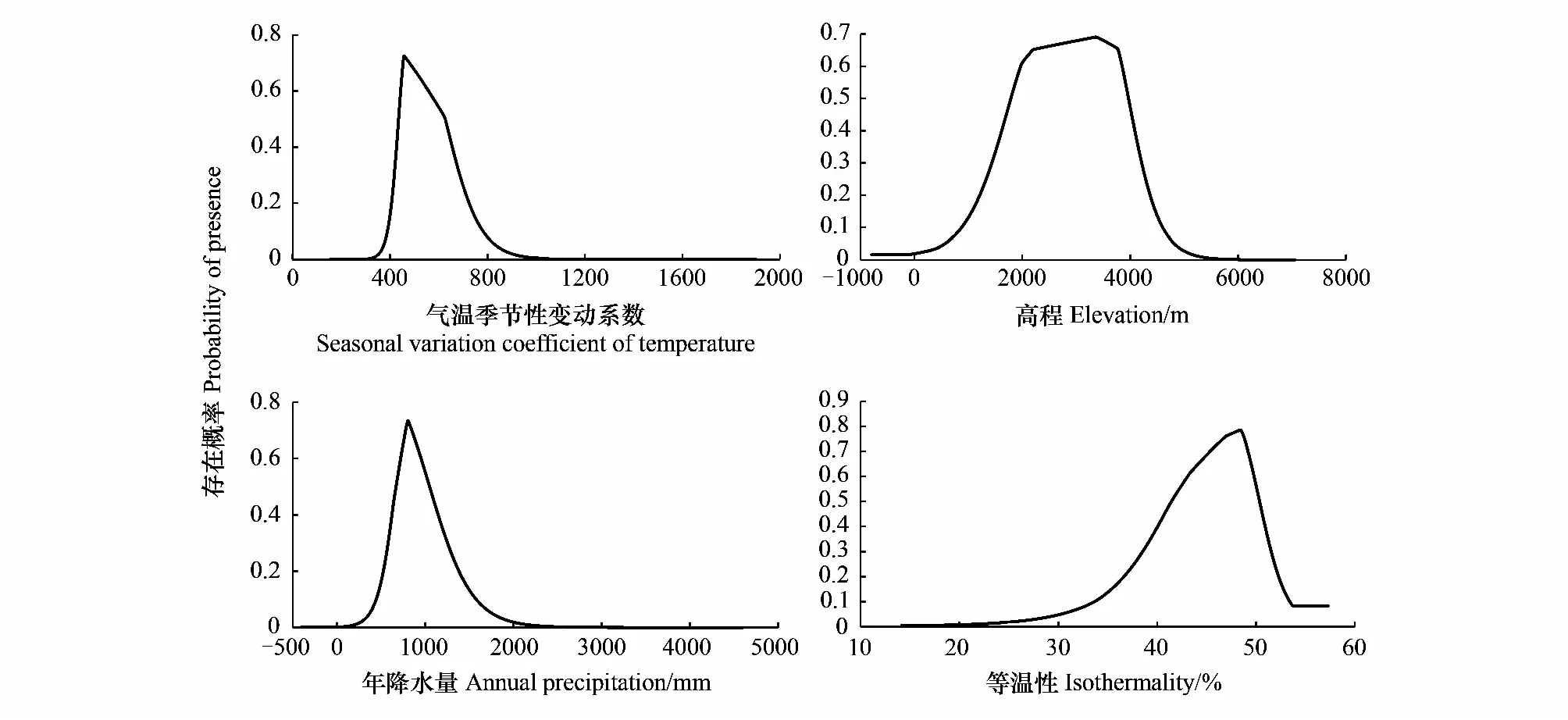
图3 潜在适生区与单因子响应变量的关系Fig.3 Relationship between potential suitable areas and single factor response variables
基于上述4个主导环境因子分别进行单因子建模,单因子响应曲线表征预测物种的存在概率随每个环境变量之间的变化关系,可以清晰揭示高山栎组植物的适生区存在概率与主导环境因子之间的相关变化趋势。通常认为存在概率大于0.5可视作最有利于物种的生长范围[45]。从单因子响应曲线(图3)可知,当气温季节性变动系数(bio4)小于340时,高山栎组植物存在概率几乎为0,其存在概率随bio4的增加而提高,当气温季节性变动系数为456时,高山栎组植物分布概率达到峰值,此时为其最适生存条件,之后随着bio4的继续增大,存在概率逐渐降低,当达到1068时,其存在概率再次接近于0。以存在概率大于0.5为最适生存范围,高山栎组植物气温季节性变动系数(bio4)的最适生存范围为435—625。与气温季节性变动系数(bio4)类似,高山栎组存在概率随高程(dem)、年降水量(bio12)和等温性(bio3)的增加而提高,达到最适生存峰值后随高程(dem)、年降水量(bio12)和等温性(bio3)的增加而降低,最适峰值分别为3366m、805mm、48.4。高山栎组植物高程(dem)最适范围为1792—3978m,年降水量(bio12)最适范围为670—1050mm,等温性(bio3)最适范围为41.5—50.3。
2.3 当代气候下高山栎组的潜在适生区
由图4可知,高山栎组植物主要分布区范围为25°6′32″—33°2′10″N,99°32′45″—102°39′4″E。集中分布于我国西南地区及藏东南地区。
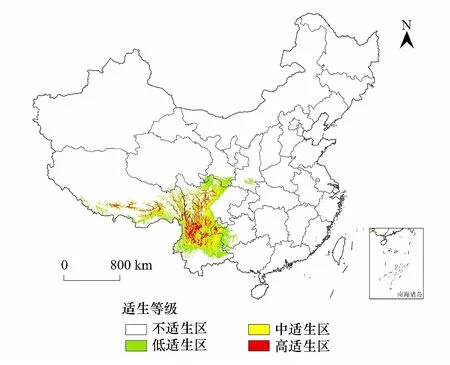
图4 当前气候条件下高山栎组在我国的潜在适生区 Fig.4 The potential suitable area of Quercus sect. Heterobalanus in China under current climatic conditions
总适生区:高山栎组总适生区主要分布于我国的云南省中部、北部,四川中部、西部,西藏东南部,贵州西部少部分地区,甘肃南部少部分地区以及青海南部极少部分地区,适宜生境总面积62.37×104km2。
高适生区:高山栎组植物最适宜生境主要分布于我国云南省昭通东南部、丽江市、大理东北部少部分地区、迪庆东部、曲靖西北部、楚雄北部少部分地区,贵州省毕节市西部,四川省甘孜东南部、凉山、攀枝花、阿坝南部,西藏昌都、林芝、山南少部分地区。高适生区面积10.47×104km2,占总适生区面积的16.79%。
中适生区:高山栎组中适生区多位于高适生区周围,主要分布在我国云南省中部的昆明市、楚雄州、昭通市中部、曲靖市中部、大理州、怒江北部、保山东部、红河州北部;贵州省毕节市中部;四川省凉山州东北部少部分地区、雅安北部和西南部少部分地区;西藏山南东南部分地区、林芝中部及东南部。中适生区面积20.94×104km2,占总适生区面积的33.58%。
低适生区:低适生区主要分布在我国云南省玉溪市、普洱市北部和中部、临沧东北部、保山中部、怒江南部、文山西北部、昭通东北部;贵州毕节西部、六盘水、兴义市西部和安顺西部少部分地区;四川乐山西南部、雅安西部、绵阳西北部、阿坝中北部、甘孜中部;西藏昌都东部少部分地区、林芝中部东南部和山南东南部少部分地区。低适生区总面积30.96×104km2,占总适生区面积的49.64%。
2.4 过去及未来高山栎组潜在适生区模拟
研究共选取6个时期来预测高山栎组的潜在分布。从表4和图5可以看出,从末次间冰期到当代,高山栎组总适生区从68.90×104km2(LIG)扩大到73.08×104km2(LGM),再缩小到68.31×104km2(MH),潜在总适生区面积在末次盛冰期达到最大,呈现先增加后减小单峰趋势。且其高适生区和低适生区呈现相同的变化趋势。中适生区则呈现逐渐降低的趋势。
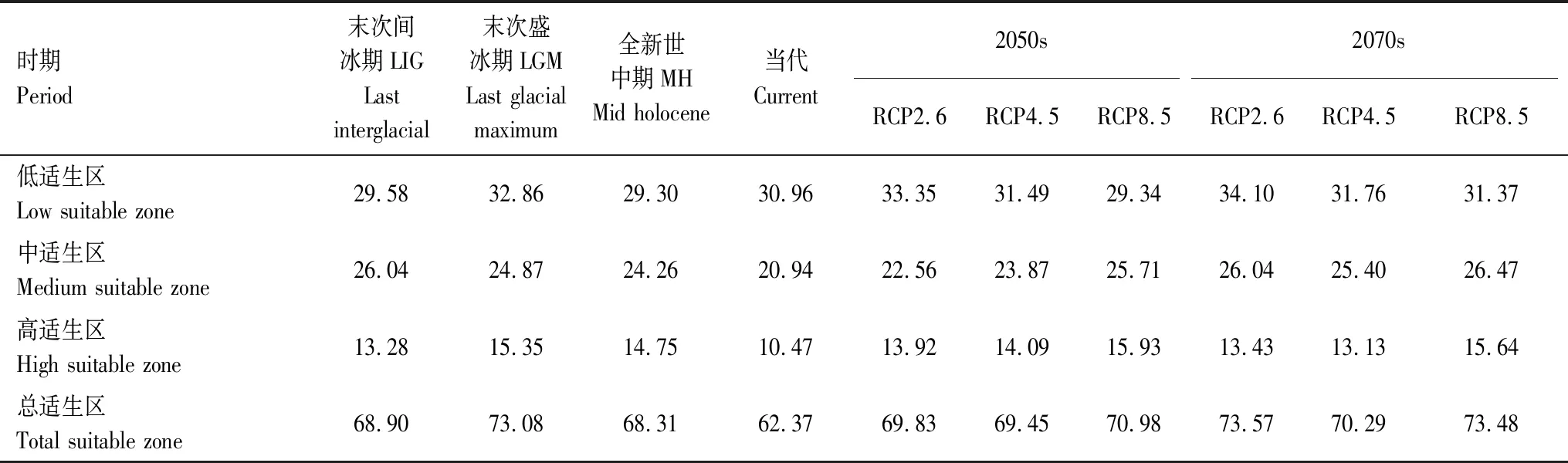
表4 不同时期高山栎组适生区面积/(×104km2)
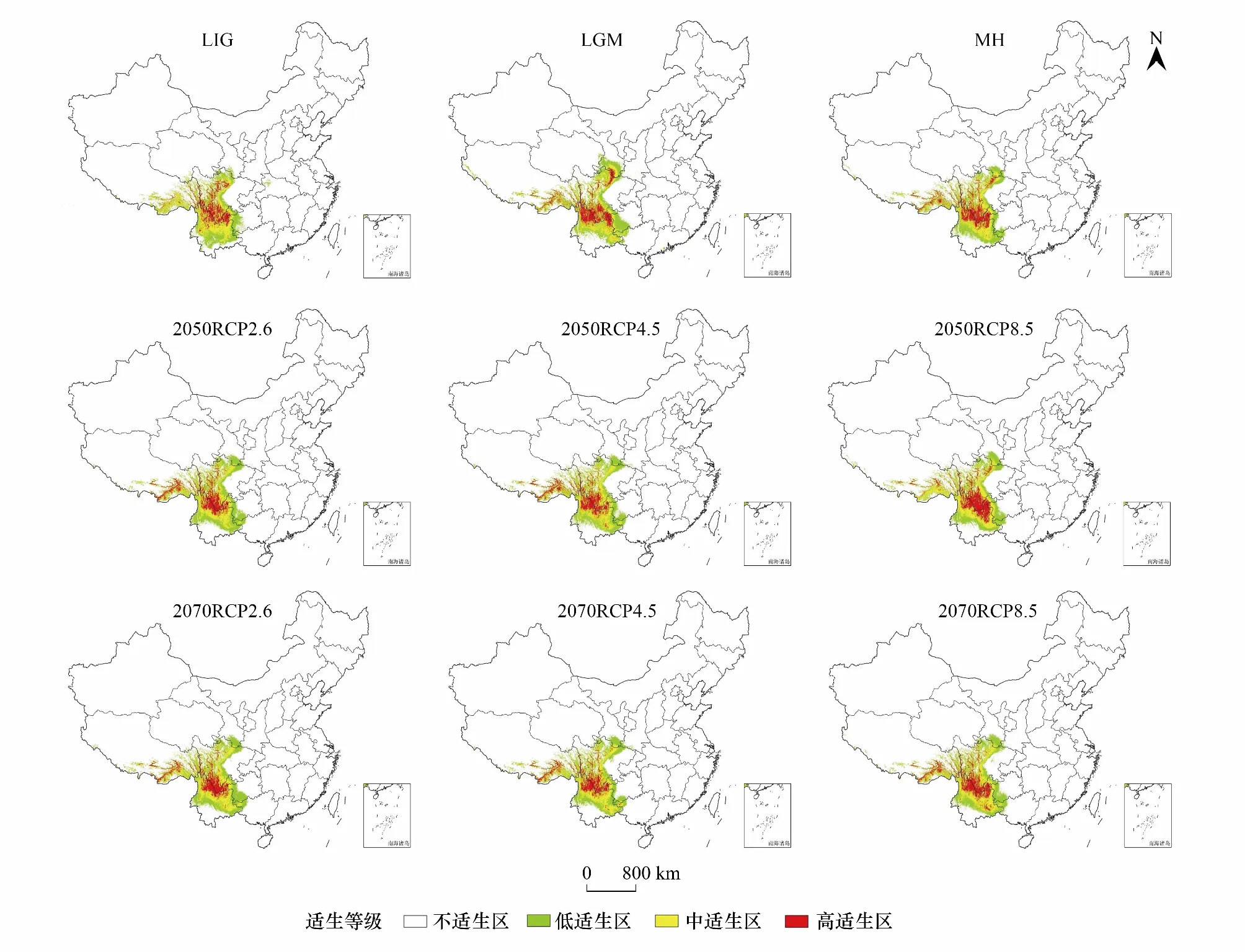
图5 不同时期高山栎组潜在适生区预测Fig.5 Prediction of potential suitable areas of Quercus sect. Heterobalanus in different periodsLIG:末次间冰期 Last interglacial;LGM:末次盛冰期 Last Glacial Maximum;MH:全新世中期 Mid Holocene;RCP:典型浓度路径 Representative concentration pathways;RCP2.6:温室气体低浓度排放情景 Low concentration greenhouse gas emission scenario;RCP4.5:温室气体中浓度排放情景 Medium concentration greenhouse gas emission scenario;RCP8.5:温室气体高浓度排放情景 High concentration greenhouse gas emission scenario
从表4及图5可以看出,从当代到未来,则呈现面积逐步增大的趋势。2050年三种情景下,总适生区面积都在69×104km2以上。其中2050sRCP2.6情景下,总适生区面积69.83×104km2,高适生区面积13.92×104km2,相较于当代分别增加7.46×104km2、3.45×104km2;2050sRCP4.5情景下,总适生区面积为69.45×104km2,高适生区面积14.09×104km2,相较于当代分别增加7.08×104km2、3.62×104km2;2050sRCP8.5情景下,总适生区面积70.98×104km2,高适生区15.93×104km2,相较于当代分别增加8.61×104km2、5.46×104km2。2070年三种不同的气候情景下,总适生区面积都在70×104km2以上。其中2070sRCP2.6情景下,总适生区面积73.57×104km2,高适生区面积13.43×104km2,相较于当代分别增加11.2×104km2、2.96×104km2。2070sRCP4.5情景下,总适生区面积为70.29×104km2,高适生区面积13.13×104km2,相较于当代分别增加7.92×104km2和2.66×104km2。2070sRCP8.5情景下,总适生区面积为73.48×104km2,高适生区面积15.64×104km2。总体来看未来两个时期不同情景相较于当代均有增加趋势,但各个时期内可看出高浓度排放路径相较于低浓度排放路径潜在适生区面积有降低趋势。
由图5可以看出,高山栎组最适分布区在不同时期有不同程度的差异,但主要分布在滇东北药山,滇西北白马雪山、浪都山、梅里雪山,云南、四川、西藏三省交界处的横断山脉,西藏东南部念青唐古拉山脉、唐古拉山脉以及巴彦喀拉山脉等山峡部分地区。另外,其潜在分布区在过去三个时期比较集中,而在当代可能由于气候变化以及人类干扰等影响使得潜在地理分布区则呈现破碎化斑块状态,在未来两个时期其适生区斑块破碎化态势有所缓和。
2.5 高山栎组潜在适生区空间格局变化
从表5和图6可以看出,与当代相比,从末次间冰期到全新世中期,高山栎组适生区面积空间格局呈现先增加后减小的趋势。末次间冰期新增面积为14.30×104km2,丧失面积7.77×104km2,其适生区增加率和丧失率分别为22.93%和12.45%,此时期面积新增地区主要集中在云南中部元江、红河、普洱,西藏林芝东南部等地区,其丧失区主要集中在甘肃南部成县,陕西秦岭部分地区,西藏西南部等地区。末次盛冰期新增面积最大,为18.54×104km2,新增率为29.73%,但丧失面积也最大,为7.79×104km2,丧失率为12.49%,总体变化率也最大,为17.24%。表明在末次盛冰期寒冷的气候条件下,高山栎组适生区面积发生大的变迁。模型预测新增的面积主要集中在云南、广西、贵州三省交界大娄山地区以及甘肃南部祁连山,此地区独特地势可能成为高山栎组适宜生长的避难所,而丧失区主要集中在云南中部海拔为1000—1500m地区以及秦岭地区。全新世中期面积增加率和丧失率最小,保留区面积相对较大,为55.04×104km2。主要丧失区在甘肃南部祁连山地区。
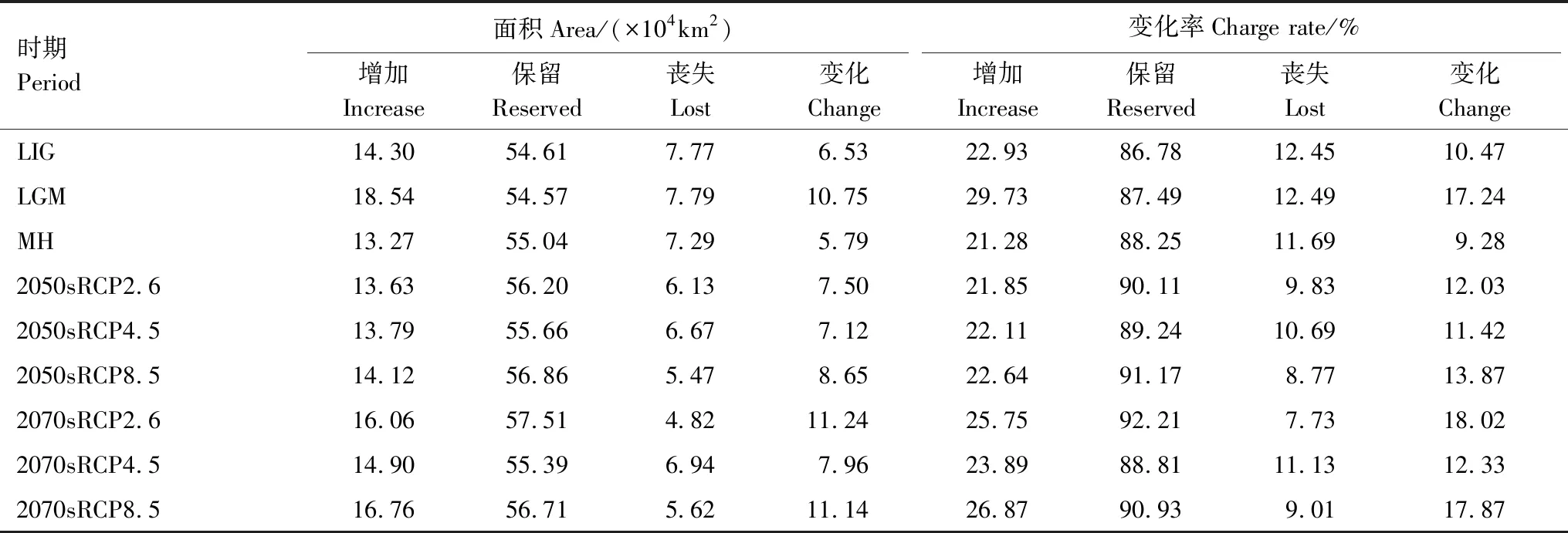
表5 不同气候情景下高山栎组适生区空间变化
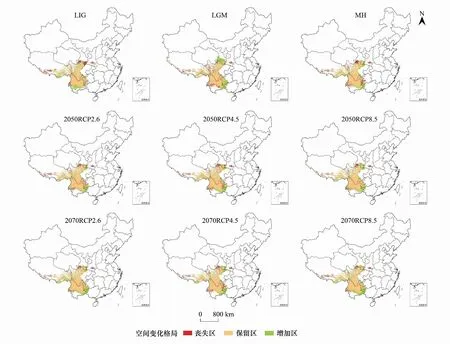
图6 不同气候情景下高山栎组适生区空间格局变化Fig.6 Suitable distribution changes of Quercus sect. Heterobalanus under different climate change scenarios
与当代相比,未来6种不同的气候情景下,2070sRCP2.6和2070sRCP8.5两种情景下新增面积最大,分别为16.06×104km2、16.76×104km2,新增率分别达到25.75%和26.87%,新增区域主要集中在云南、广西以及贵州交界处,甘肃南部和藏东南地区。丧失面积最小,仅为4.82×104km2和5.62×104km2,丧失率为7.73%和9.01%。其总体变化率也最大,分别达到18.02%和17.87%。2050sRCP2.6和2050sRCP8.5这两个情景下新增区面积分别13.63×104km2和14.12×104km2,新增率分别为21.85%和22.64%,丧失区面积分别为6.13×104km2和5.47×104km2,丧失率分别为9.83%和8.33%。2050sRCP4.5和2070sRCP4.5两种情景下丧失面积最大,分别为6.67×104km2和6.94×104km2,丧失率分别达到10.69%和11.13%,丧失区域主要集中在贵州省黔东南地区,陕西南部秦岭部分地区,云南省保山、德宏等地以及甘肃西南部分地区。其总适生区面积变化率最小,分别为11.42%和12.33%,表明这两个时期气候变化较小,对于高山栎组适生区影响不大。
总体来看,未来气候情景下,高山栎组潜在分布适生区集中性增强,破碎化程度降低,其分布丧失区主要集中在秦岭地区,云南西部德宏以及西藏西南部分地区,增加区主要向云南、贵州、广西三省交界处以及甘肃南部祁连山脉部分地区扩张,表明这些地区可能是其在未来格局变化的敏感区,应予以重视。
2.6 适宜生境质心的转移
研究将高山栎组潜在适生分布区以几何中心点来定义,以此方式来模拟在不同时期气候情景下质心迁移变化[43]。从表6和图7可以看出,末次间冰期(LIG)相较于当代质心位置偏南,迁移距离4.945km;末次盛冰期(LGM)质心位于当代的东北部,相对于当代迁移6.936km,且相对于其他时期与末次间冰期相距最远,这也间接表明从末次间冰期到末次盛冰期气候变化明显。全新世中期质心位于当代的东南部,与未来气候情景下质心位置相近,表明未来由CO2增加引起的气候变暖与全新世增温结果相似[46],迁移距离为3.629km。从末次间冰期到当代,总体呈现出先向东北迁移,然后向西南,最后向西北迁移的趋势,且当代海拔相较于过去三个时期最高,为3955m。2050年相较于当代质心向东南迁移,而2070年则在2050年的基础上继续向东南迁移,2070sRCP8.5情景下质心相较于当代最远,为5.908km。2050sRCP2.6情景下海拔较当代升高,2050年其他气候情景与2070年三种气候情景海拔相较于当代有不同程度的降低。
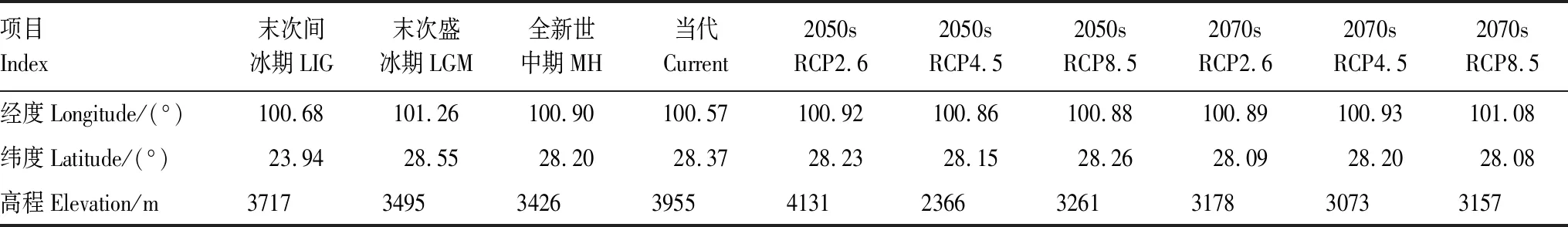
表6 不同时期高山栎组经纬度及海拔变化趋势
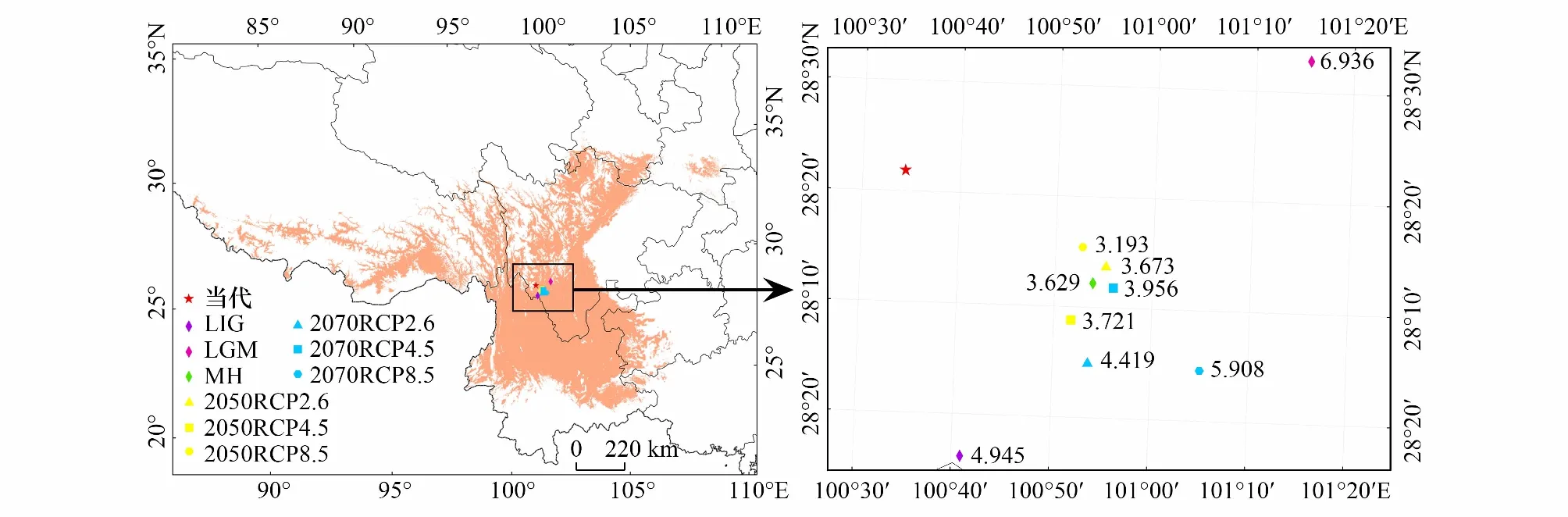
图7 不同时期气候情景下高山栎组适生区质心迁移位置/kmFig.7 Location of centroid shift of suitable areas of Quercus sect. Heterobalanus under different climate scenarios in different periods图中质心点旁的数字表示该点位置相对于当前质心点的距离
3 讨论
3.1 影响高山栎组分布的主导环境因子
MaxEnt模型已经成为生态学、保护生物学以及进化生态学研究的一个重要工具[47]。研究根据MaxEnt模型预测高山栎组植物潜在分布格局。通过刀切法训练增益、贡献率、单因子响应曲线揭示出影响高山栎组植物最主要的环境因子是气温季节性变动系数(bio4)、高程(dem)、年降水量(bio12)和等温性(bio3)。其中气温季节性变动系数(bio4)对高山栎组植物影响最大。由此可见,温度是影响高山栎组植物分布最重要的环境因子,其次为高程(dem)和降水量。
植物在区域尺度上的地理分布主要受气候的制约,其中水热条件起主导作用。Liao等在对中国西南横断山地区高山栎组7种植物的潜在分布及驱动因素研究中表明温度季节性是影响高山栎组植物最重要的影响因子[19],与研究结果完全吻合。Yang等在对喜马拉雅东部和横断山地区常绿硬叶栎的潜在分布进行研究时,表明温度是影响其大规模分布的主要因素,降水是其次因素[48]。由此可见温度相关变量对高山栎组的影响大于水分相关变量的影响。青藏高原和横断山区独特的地理和气候条件,受东亚和西南季风的影响导致湿度较大,使得高山低温的环境比干旱更有利的控制着高山植物[49]。植物必须在一定的温度范围内才能进行光合作用和呼吸作用,并且每种植物正常生长都需要一个最适宜温度阈值[50]。过低的温度可能会导致植物遭受冰冻和霜冻伤害,从而严重抑制它们的生长,例如通过膜损伤和光合作用减少[51]。高山栎组植物是青藏高原横断山脉的典型类群[52],其分布对于环境具有广泛的适应性,形态学的研究也表明高山栎组植物叶片具有很多与防寒保水有密切关系的生态特征,如叶片厚的角质层、被毛、具复表皮、木质化的表皮细胞壁和角质层以及较低的气孔密度等,这些生理生态特征共同提高了它们对于干冷气候的适应性,以使其适应高山环境[53]。其次为高程(dem),高程主要通过间接影响温度及降水影响物种分布格局[45]。通过单因子响应曲线可知适宜高山栎组植物生长的高程范围为1792—3978m,这与周浙昆等关于高山栎组演化的研究结果类似[25]。
3.2 高山栎组植物潜在适生区空间分布格局变化及质心分析
了解气候变化背景下物种潜在分布格局的变化,对于衡量气候变化对物种的影响和制定保护策略以维持生态平衡至关重要[54]。不同物种的分布受气候变化的影响不同。研究运用MaxEnt模型和ArcGIS软件对高山栎组植物不同时期的潜在地理分布进行模拟。揭示出高山栎组植物分布范围在25°6′32″—33°2′10″N,99°32′45″—102°39′4″E之间,这与Liu基于模型与实地调查的结果基本一致[55],表明研究基于优化的MaxEnt模型对高山栎组植物模拟精度高、效果好。气候是决定物种生存分布的最重要的因素,植物的分布特征是对气候最直接的反应[56]。研究从地理适生区分布图中可以看出,高山栎组植物主要分布在沿山脉走向的山峡中,这与Liao等的研究结果相吻合[19]。从末次间冰期到当代,其分布适生区面积呈现减小的趋势,末次盛冰期面积达到最大,高山栎组分布区在此时期发生了一定的扩张,这与叶利奇等对于横断山珙桐[57]的研究相符。推测这可能是因为喜马拉雅山脉的抬升阻挡了暖湿的印度洋气流,从而使得横断山地区变得寒冷而干燥,由于高山栎组植物特殊的生物学及生理生态适应机制,即使面对高寒的气候环境,对其影响也比较小[58],且此前的研究也发现,高山栎组植物具有相似的耐寒耐旱能力[48]。此外,在末次间冰期、末次盛冰期和全新世中期,高山栎组潜在适生区集中性强,而在当代,其潜在适生区斑块破碎化严重,这与张明珠[34]在对长序榆的研究结果相似,推测这可能是由于当代能源消耗量增加、CO2排放量增大、砍伐、开荒等人类活动加剧的生态破碎化有关[59]。在未来6个时期,高山栎组植物潜在适生区相较于当代在不同的气候情景下均有不同程度增加,可能高山栎组植物可以较好的适应未来气候条件。但在未来的分布格局中,2050s和2070s低浓度排放情景丧失率较小,而高浓度排放情景丧失率较大,推测可能是由于未来气候条件下,高浓度排放情景由于气温和降水变化剧烈且不规律,极端事件多,从而对高山栎组植物影响较大,导致生境破碎化加剧[60—61]。因此,我们的结果也表明,在未来全球气候变暖的大背景下,高山栎组植物将面临生存风险。
质心分析结果表明从末次间冰期到当代,其质心表现为东北—西南—西北的迁移趋势。相较于当代,未来质心向东南迁移。海拔整体表现为先升高再降低的趋势,且在当代最高。相关研究表明未来气候情景下,随着全球气候变暖,绝大多数动植物有着向高纬度及高海拔的趋势发展[62]。研究预测的潜在分布区质心则有着相反的趋势,即向低纬度和低海拔迁移的趋势。其西北方向分布区生境适应性在温暖气候将降低,分布西北界明显受到环境的限制[63]。历史上高山栎组植物随地势抬升逐步适应高山环境,未来气候变化中也可能向低海拔迁移扩散。同时,物种潜在地理分布并非仅局限于气候和地形因素,气候复杂性并不仅仅涉及到温度和降水,同时还影响到光照辐射强度,土壤C循环,臭氧层变化等;同时竞争、繁殖等生物因素、人类活动以及其他非生物因素物理障碍也会对植物生长产生影响[64]。因此,在未来的研究中可以综合植物本身的生理生化作用、生物之间的相互作用、生态系统变化以及人为因素等做出精准预测,以提高模型的精确度。
4 结论
研究基于ENMeval包优化的MaxEnt模型预测了高山栎组植物在过去3个时期,当代以及未来6种不同的气候情景下潜在适生区以及分布格局。
(1) 高山栎组植物在当前气候条件下潜在适生区面积62.37×104km2,占中国国土面积的6.5%,主要分布在我国的横断山脉及西藏东南部沿山脉走向的念青唐古拉山脉、唐古拉山脉以及巴彦喀拉山脉等山峡地区。
(2) 气温季节性变动系数(bio4)、高程(dem)、年降水量(bio12)和等温性(bio3)是影响高山栎组植物分布的主导因子,高山栎组植物最适生存概率出现在气温季节性变动系数(bio4)为435—625、高程(dem)1792—3978m、年降水量(bio12)670—1050mm、等温性(bio3)41.5—50.3。且气温季节性变动系数(bio4)是影响高山栎组植物最重要的环境因子。
(3) 历史时期高山栎组潜在适生区分布较为集中,当代破碎化严重,未来三个时期不同代表性排放浓度路径下其适生区均有不同程度的扩张,尤其在RCP2.6排放情景下扩增面积最大。但是高浓度排放路径RCP8.5情景短期内虽有利于高山栎组植物扩增,但是长远来看并不利于其生长发育。从质心变化来看,随着未来气候变化,高山栎组植物的质心由高纬度向东南部等低纬度扩散,从高海拔向低海拔迁移。
(4) 未来气候情景下,虽有利于高山栎组植物生长发育。但是人类活动也不断加剧,代表性CO2浓度排放也受到国家碳中和调控政策的影响,因此其在未来如何发展变化也将呈现一定的不确定性。当前气候条件下,高山栎组分布区零星分散,破碎化程度高,人类干扰程度大。因此,就其长远发展,应减少人为干扰。对于原生性群落,要针对不同情况加以保护,并通过植被抚育以增加其分布范围,充分地发挥其生态效益与经济效益。
