上海大金山岛植被功能组成对草本植物、土壤动物和细菌多样性的影响
2023-09-11杨柏钰赛碧乐杨安娜张增可李曾燕王万胜杜运才于再平阎恩荣
杨柏钰,赛碧乐,杨安娜,张增可,李曾燕,张 立,牟 凌,王万胜,杜运才,刘 海,于再平,郭 超,阎恩荣,*
1 华东师范大学生态与环境科学学院,上海 200241 2 上海市金山区海洋海塘管理所,上海 201508 3 浙江舟山市普陀山林场, 舟山 316107
生物多样性是生态系统功能形成的基础[1]。在森林生态系统中,生态系统功能是由不同营养级生物多样性共同维持的[2—3]。植物多样性决定了生态系统生产力大小和物质返还量[4],土壤动物和微生物多样性驱动了凋落物分解和养分循环过程[5]。在此过程中,由于上行效应和下行效应,不同营养级或同一营养级内生物间也会相互影响[6—7]。例如,树木多样性既会通过改变群落结构组成、林下光线与水分资源分配而改变草本多样性[8—9],也会通过改变输入的叶片与枯木凋落物的数量与质量而改变土壤动物和微生物多样性[10—13];同样,由于负反馈作用,土壤动物和微生物多样性也会通过调节养分周转和土壤肥力,反过来影响草本和树木多样性[14—15]。因此,植物多样性与土壤动物和微生物多样性之间存在着密切的生态关联[16—17]。但是,我们对于不同营养级生物多样性相互关系及其驱动机制的理解仍处在探索阶段[18]。
过去二十多年中,生态学家探索了不同生态系统中草本或木本植物多样性对土壤动物及微生物多样性的影响[19—22]。研究发现,在温带草原中,由于食物资源的互补效应,随着植物多样性的增加,蚯蚓和线虫等土壤动物多样性显著增加[5];在干旱荒漠生态系统中,木本植物丰度促进了真菌等微生物多样性[20]。总体而言,虽然这些研究极大的提升了我们关于不同营养级生物多样性间相互作用的理解,但却往往从单一营养级的简单因果关联的角度出发,忽略了多营养级之间的相互反馈[6—7, 23]。同时,根据生物级联效应(cascading effect),上一营养级对下一营养级生物的作用过程可能同时包括了直接影响与间接调控[24]。然而,目前关于生物级联效应对多营养级多样性关系的影响仍没有明确的研究结论。
植物多样性对土壤动物和微生物多样性的影响主要取决于其返回土壤的凋落物基质的数量、质量与多样性[12],也就是说,植物多样性的功能属性组成是调控土壤动物和微生物多样性的主要因素之一[25—27]。研究表明,在热带和亚热带森林中,常绿与落叶树种的占比既会通过改变林下光斑大小,从而影响草本多样性[28—29],也会通过改变凋落物基质组成多样性,从而影响土壤动物和微生物多样性[10, 30—31]。同时,部分研究表明,树木物种多样性对土壤动物与微生物的影响较弱[32]。基于此,本文预测,相对于树木物种多样性而言,亚热带森林中落叶树木的占比(也就是功能组成)在调控林下草本、土壤动物与微生物多样性方面扮演更为重要的角色。由于不同营养级生物间既存在上行效应,也存在下行效应[6],加之级联效应所产生的直接与间接调控[24],本文提出科学预测1: 森林群落树木的功能组成(即落叶树木占比)既可以直接影响土壤动物和微生物多样性,也可以通过草本多样性的正向级联效应而间接调控土壤动物和微生物多样性。
另外,作为腐食食物链的主要构成,土壤动物和微生物群落间也存在着紧密的竞争、互惠合作与捕食关系[33—34]。先前基于亚热带森林的研究表明,土壤无脊椎动物可通过啃食树木木质残体而为微生物提供食物来源[11]。因此,本文进一步预测(科学预测2):群落中落叶木本植物占比的增加会加剧土壤动物与微生物之间的负向关系。具体而言,由于存在食物资源竞争关系,因此,随土壤动物多样性增加,微生物多样性下降。
上海大金山岛自1993年建立保护区起,封山育林30余年,探究植被功能组成和物种多样性对土壤动物及细菌多样性的调控作用,以及土壤动物和细菌多样性对植被物种多样性的反馈影响,对于揭示森林多营养级生物多样性的保育具有重要意义。综上,为检验所提出的科学预测,本研究以上海大金山岛包含不同落叶木本植物占比的13个植物群落为对象,在分析植物群落树木物种多样性、草本物种多样性、土壤动物和细菌多样性关系的基础上,探索土壤养分梯度上,群落中落叶树木占比对草本、土壤动物和细菌多样性的影响效应,从而为保护大金山岛的生物多样性提供理论支撑。
1 方法
1.1 研究区概况和样地选择
大金山岛(30°41′42″ N, 121°24′25″ E)地处上海金山区附近的杭州湾, 位于中亚热带北缘, 是金山三岛海洋生态自然保护区的核心区。大金山岛总面积为0.229 km2, 主峰高103.4 m, 其年平均气温15.8 ℃, 土壤为褐色山地黄壤, 降水充沛, 属海洋性气候。大金山岛植被从外貌水平可划分为草甸、灌草丛、落叶阔叶林、落叶常绿阔叶混交林和常绿阔叶林5个类型,群落物种组成中落叶植物按一定比例下降[35]。
2016年7月,选择灌草丛、落叶阔叶林、落叶常绿阔叶混交林和常绿阔叶林植被类型,根据分布面积大小,选择其中的典型植物群落建立了13个长期固定观测样地,落叶阔叶灌丛包括3个日本野桐(Mallotustenuifolius)群落样地和1个小叶女贞(Ligustrumquihoui)群落样地,落叶阔叶林包括麻栎(Quercusacutissima)群落、朴树(Celtissinensis)和丝棉木(Euonymusmaackii)群落样地各1个,常绿落叶阔叶混交林群落包括椿叶花椒(Zanthoxylumailanthoides)-柃木(Euryajaponica)群落样地1个,常绿阔叶林包括普陀樟 (Cinnamomumjaponicum)群落和红楠(Machilusthunbergii)群落样地各1个,青冈(Cyclobalanopsisglauca)群落样地3个。其中,由于小叶女贞群落分布范围小,样地面积为100 m2,其余群落类型的样地面积均为400 m2。另外,由于日本野桐群落和青冈群落分布面积较广,根据坡位和地形变化,每个类型各建立了3个同样面积的长期监测样地。
样地建成后,根据统计样方法对每个样地进行群落学特征调查,对于胸径大于1 cm,且株高大于0.5 m的所有木本植物,逐木确定物种名称,测量胸径、树高与冠幅。对于草本层,按照典型样地调查法,确定物种组成,记录各物种的个体数、平均盖度与株高。各样地的群落学特征见表1。
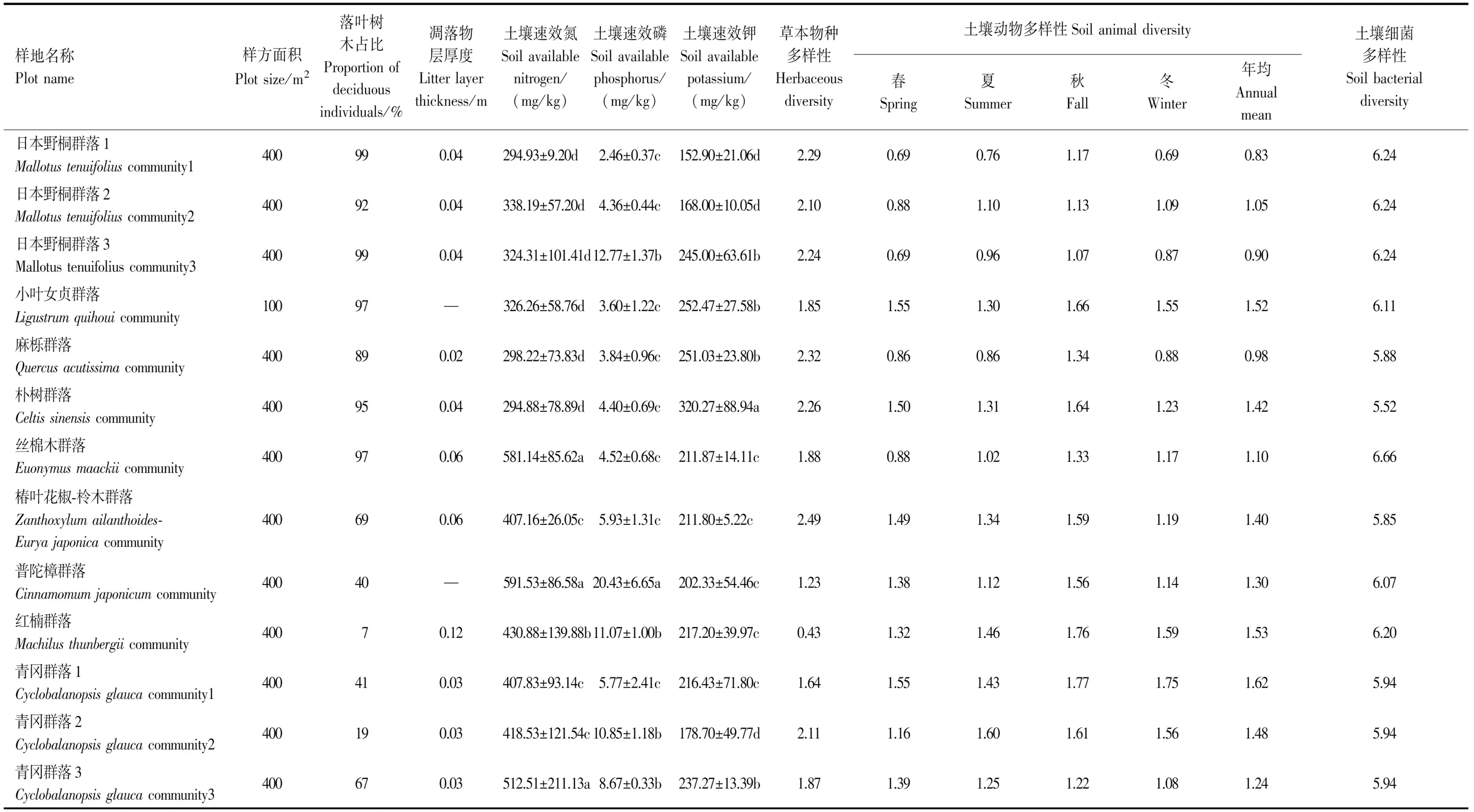
表1 大金山岛13个典型植被样地的基本特征
1.2 土壤理化因子测定及土壤无脊椎动物群落调查
在完成植物群落特征调查后,在每个样地中,随机设置8—10个采样点,利用土钻提取0—20 cm土样,带回实验室自然风干后进行土壤理化性质分析。样品风干后,去除石粒和根系等植物残体,用研钵磨碎,过60目网筛,用以测定土壤速效养分。土壤速效钾含量采用乙酸铵浸提-火焰光度法测定,土壤速效磷和速效氮含量分别采用氟化铵-盐酸浸提法和联合浸提-比色法进行测定[36]。
本文在2017年春季(3月)、夏季(6月)、秋季(9月)和冬季(12月),分四个季节调查土壤动物群落。采样时,在每个固定样地中,按照梅花桩法设置了5个采样点,利用直径为5 cm的不锈钢环刀,对表层土壤(0—10 cm)取样,然后将土柱放至自封袋,在4 ℃条件下运回实验室处理。在实验室,利用Tullgren漏斗法(25 w,72 h)分离中小型土壤动物[37]。分离出的土壤动物保存在75%的酒精中,利用体式显微镜(Leica, DVM6c, Germany)鉴定和计数。根据尹文英等,所有土壤动物鉴定至目水平[38—39]。
1.3 土壤细菌群落调查
在2017年夏季(6月)土壤动物取样的同时,也按照梅花桩5点取样法,采集土壤细菌调查土样。取样时,去除土壤表面叶片与枯枝凋落物,利用土钻收集表层0—10 cm土样,利用4℃保温盒快速运回实验室。在实验室,每个样地的5份土样混合,过2 mm筛,再选取3份,采用MoBio powerSoil DNA提取试剂盒(MoBio Laboratories, Carlsbad, CA, USA)提取土壤微生物DNA,并以16S核糖体rRNA基因V4区为目标区域,采用引物515F(5′-GTGCCAGCMGCCG CGGTAA-3′)和806R (5′-GGACTACHVGGGTWTCTA AT-3′) 进行PCR扩增。PCR反应程序为在94 ℃温度条件下扩增35个循环,其中包括在94 ℃温度条件下变性10 s以及在52 ℃温度条件下复性30 s,最后在72 ℃温度条件下延伸45 s,共计5 mins。最后保持72 ℃温度条件不变,将产物延伸10 mins。将得到的PCR产物纯化并在llumina Nova 6000平台上进行扩增子测序。使用Usearch v10软件将原始数据进行拼接和质控,使用代码“-fastq_mergepairs”合并双端序列,使用cutadapt软件去掉序列中的引物,将长度小于300 bp的序列和低质量序列舍弃,最后去除重复序列,得到有效拼接片段。随后使用UPARSE算法对超过97%相似度阈值的OTU (Operational Taxonomic Units)进行分类学分析,其中,通过检测并删除嵌合体,形成了最初的OTU表并比对SILVA 132数据库,采用RDP分类器(置信阈值为0.8)对每个OTU进行物种注释。为保持测序深度的一致,对样品的序列总数进行抽平处理,抽平后每个样品的细菌序列总数为30610条。最后,得到了OTU水平上的细菌群落组成与结构信息。
1.4 群落落叶成分比例和生物多样性指数计算
本研究选取落叶树木个体数占群落总个体数的占比(DP),用以反映群落中树木功能成分比例对不同营养级生物多样性的影响,计算依据公式1:
(1)
式中,Nd/e为每个样方中落叶树种/常绿树种个体数,N为每个样方中树木总个体数。
如公式2所示,本研究采用香浓维纳指数(Shannon-Wiener,H′)反映树木和草本多样性、土壤动物多样性及细菌多样性。
(2)
式中,Pi=Ni/N,Ni为第i物种的相对多度,N是所有物种的总个体数。
1.5 统计分析
首先,利用Pearson相关分析检验了土壤养分含量、树木物种多样性、落叶树木占比、草本物种多样性、土壤动物多样性与细菌多样性之间是否具有关联性。其中,前人研究表明[16,40],土壤速效养分常常集中分布在土壤表层(0—20 cm),且在相邻年际间差异较小,因此为了捕捉到群落整体的土壤速效养分情况对多营养级多样性的调控作用,本文将土壤速效养分、落叶木本植物占比、土壤动物多样性、微生物多样性放在一起探索其关联[16—17]。随后,为了探究林分不同生物类群多样性之间的相互作用,利用结构方程模型(Structural Equation Modeling, SEM)探究群落落叶树种比例如何通过改变草本层物种多样性而间接影响土壤动物多样性及细菌多样性。前人研究表明,虽然森林生态系统中上行效应与下行效应并存,但往往以上行效应为主[7]。因此,本文SEM中以木本植物与草本植物等低营养级物种多样性作为效应变量,以土壤动物与细菌等高营养级类群作为响应变量。同时,由于同一营养级间的相互作用,且占据空间资源的优势类群作用更大,SEM包括了木本植物多样性对草本物种多样性的直接影响路径,以及土壤动物多样性对细菌多样性的直接影响路径[7]。考虑到不同营养级间的级联作用,SEM纳入了木本植物通过草本物种多样性对土壤动物与细菌的间接路径[24]。另外,土壤养分是影响各营养级生物多样性的主要因素之一,本文分析发现,落叶树种占比与土壤速效磷显著相关,而与速效氮和速效钾无显著关系(详见结果2),因此,SEM结构中,土壤速效磷也被纳入作为影响各营养级生物多样性的效应变量。最后,由于土壤动物的季节变化较大,为更具代表性,本研究SEM选择年均土壤动物多样性(H′)。
模型分析过程中,依据Pearson相关分析结果作为模型变量筛选准则,选择与不同类群物种多样性相关关系显著(P<0.10)且相关系数值较大的变量进入SEMs。本文利用信息标准指数(AIC),卡方拟合优度检验(P),拟合优度指数(GFI)和近似误差的均方根(SRMR)等作为SEMs拟合优度的评估指标[41]。当AIC值较小,P> 0.05,GFI >0.90,SRMR<0.08时表明方程拟合结果较好且可信度较高。在对所有变量进行分析前,对数据进行了对数转换(ln)。所有的数据分析均在R4.1.2中进行。
2 结果
2.1 植物群落功能组成特征和土壤养分含量
如表1所示,13个群落的落叶木本植物占比变化范围较大(7%—99%),其中日本野桐群落1,3占比最高,小叶女贞群落、丝棉木群落和朴树群落次之,红楠群落最低。凋落物层厚度变化范围在0.02—0.12 m之间。土壤速效氮和速效磷含量在普陀樟群落最高,红楠群落次之,日本野桐群落1最低。土壤速效钾含量在朴树群落最高,小叶女贞群落和麻栎群落次之,日本野桐群落1最低。
2.2 落叶木本植物占比、草本物种多样性、土壤动物多样性与细菌多样性间的关系
由表2可见,土壤速效磷含量与落叶木本植物占比显著正相关(P<0.05),土壤速效氮和速效钾含量与落叶木本植物占比无显著相关性(P>0.10)。土壤速效养分也与各生物类群多样性无显著关联性(P>0.10)。
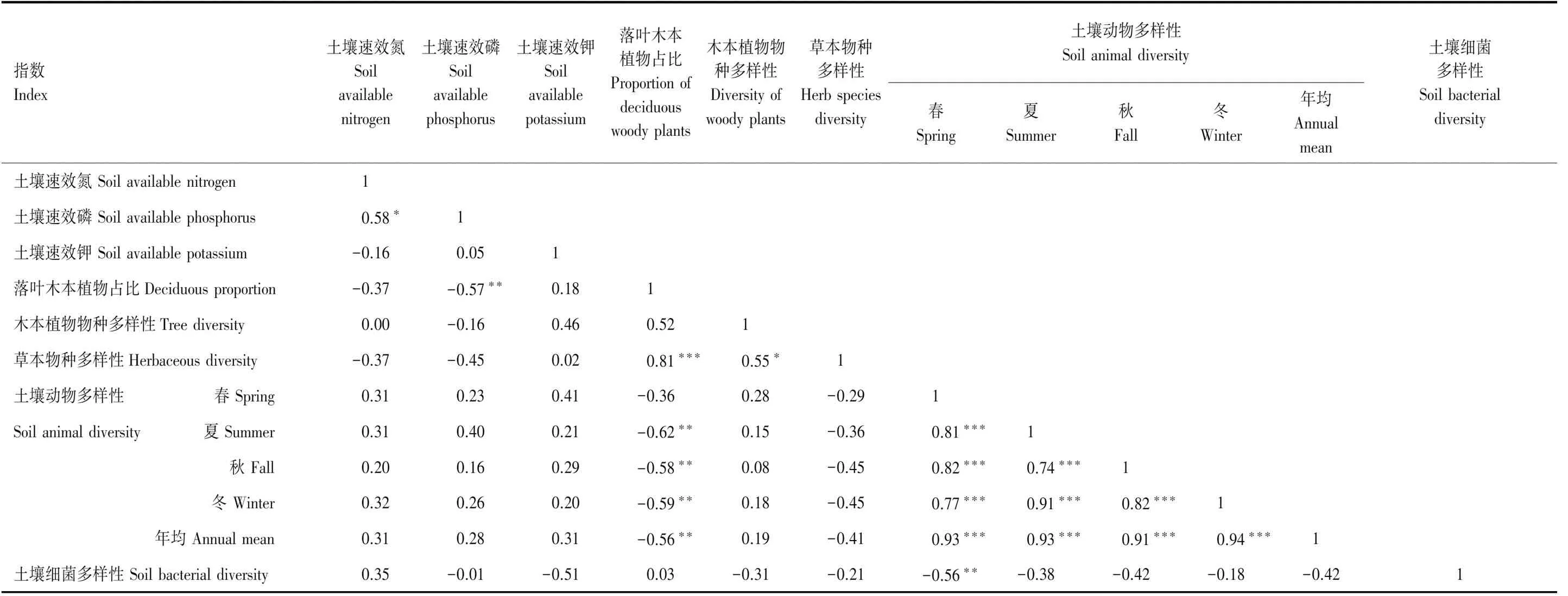
表2 土壤速效养分、落叶木本植物占比和物种多样性、草本物种多样性、土壤动物和细菌多样性之间的相关关系
落叶木本植物占比与草本物种多样性显著正相关(P<0.01),与土壤类群多样性(夏、秋、冬季和年均)显著负相关(P<0.05),但与春季土壤动物类群多样性和细菌多样性无显著相关性(P>0.10)。木本植物多样性仅与草本物种多样性显著正相关(P<0.10),而与土壤动物和细菌多样性无显著相关性(P>0.10)。草本物种多样性也与土壤动物和细菌多样性无显著相关关系(P>0.10),土壤动物多样性与细菌多样性间无显著关系(P>0.10)。
2.3 群落落叶木本植物占比对土壤动物和细菌多样性的直接和间接影响
SEM方程模型拟合效果较好(AIC=-8.438;P=0.845; GFI=0.988; SRMR=0.022),分别解释了群落落叶树木占比、草本层多样性、土壤动物多样性与细菌多样性变异的33%,66%,32%和34%(图1)。如图1和表3所示,土壤速效磷含量的增加显著抑制群落落叶树木占比(P<0.05),而对草本层、土壤动物与细菌多样性无显著影响(P>0.10)。落叶木本植物占比的增加显著促进草本层多样性,显著抑制土壤动物多样性(P<0.05);虽然落叶木本植物占比对土壤细菌多样性无直接影响,但间接通过增加草本层多样性而显著抑制细菌多样性(r=-0.47,P<0.05)。此外,草本多样性与土壤动物多样性均会直接抑制细菌多样性(P<0.10)。需要指出的是,本研究也拟合了草本物种多样性与土壤动物多样性间的作用路径,但模型被拒绝,间接说明草本物种多样性对土壤动物多样性的直接影响远远小于落叶木本植物占比的影响。
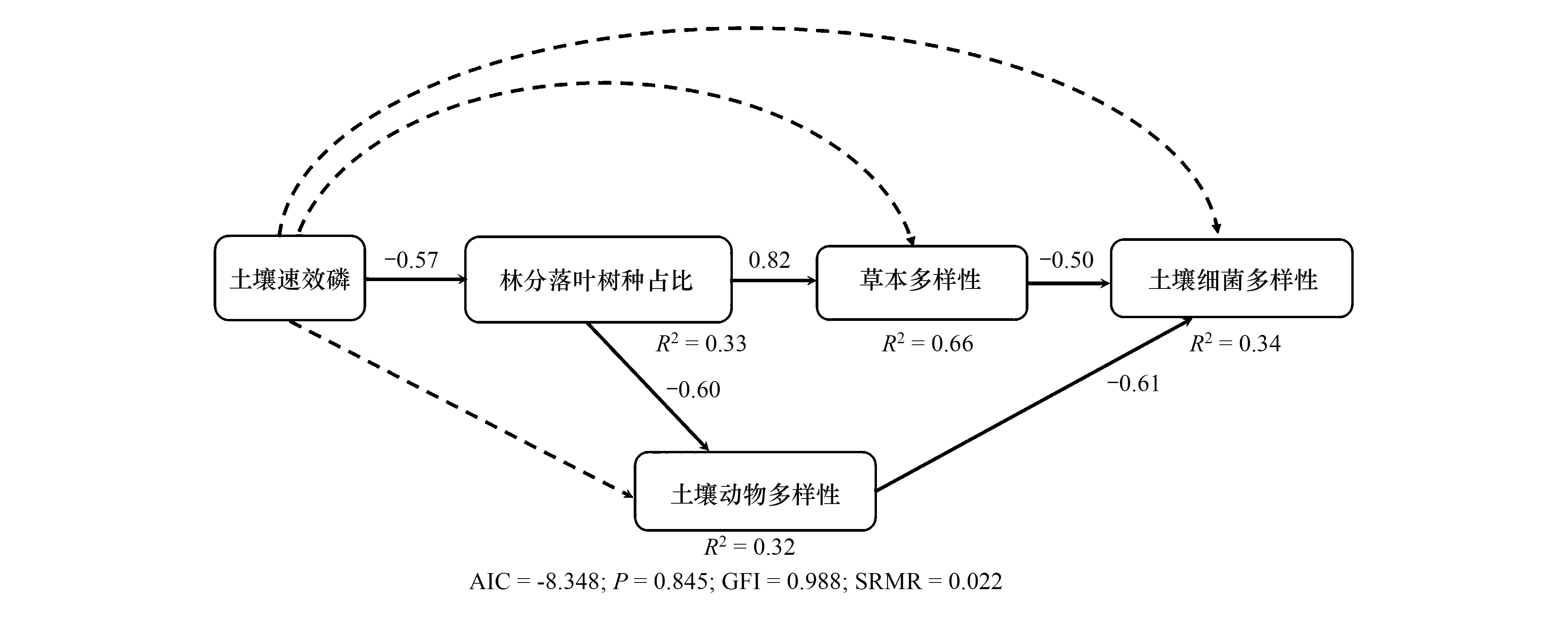
图1 群落土壤速效磷和落叶木本植物占比对草本、土壤动物与细菌多样性的直接与间接影响Fig.1 Direct and indirect effects of soil available phosphorus content and proportion of deciduous woody plants on herbaceous species diversity, soil animal diversity and soil bacterial diversityAIC:信息标准指数;P:卡方拟合优度检验;GFI:拟合优度指数;SRMR:近似误差的均方根

表3 土壤速效磷、群落落叶树木占比对草本、土壤动物与细菌多样性的影响路径和效应值
3 讨论
本研究发现,大金山岛植被中,落叶木本植物占比不仅对草本层物种多样性和土壤动物多样性分别产生直接的正和负效应,也会通过增加草本植物多样性而间接降低土壤细菌多样性。该结果支持本研究的科学假说,即:相对于树木物种多样性而言,亚热带森林的落叶树木占比(也就是群落落叶种功能组成)在调控林下草本、土壤动物与细菌多样性方面扮演着更为重要的角色。事实上,大金山岛位于中亚热带北缘,地带性植被类型为常绿落叶阔叶混交林和落叶阔叶林,与低纬度的典型常绿阔叶林不同,落叶物种是大金山岛植被组成的主体成分,是群落结构和生态过程的主要塑造者[42]。本文从多营养级角度证实了落叶木本植物占比对森林不同营养级多样性关系的调控作用,落叶树种占比是亚热带东部海岛生态系统不同营养级生物多样性维持的主要驱动因素[43]。
落叶木本植物占比与土壤动物多样性间的负向关系、草本物种多样性与土壤细菌多样性的负向关系以及土壤动物和细菌多样性间的负向关系均反应了低营养级生物对高营养级生物密切的上行调控作用[7]。落叶木本植物占比对土壤动物多样性的负效应可能是源于选择效应(selection effect)[5, 44—45],即:虽然落叶植物的群落优势效应可返还大量基质质量高的凋落物到地表,但降低了凋落物基质多样性,这样的格局有利于喜食高质量凋落物的专性类群的生长与繁殖,而不利于维持食性更为多样的土壤动物类群[46]。结合本研究中木本植物多样性与土壤动物多样性间无显著关联的结果,可以得出落叶木本植物占比所主导的选择效应决定了土壤动物多样性的分布格局。
本研究发现,落叶木本植物占比并不直接影响土壤细菌多样性,而是间接通过增加草本多样性而抑制土壤细菌多样性。一方面,该结果说明,落叶木本植物占比对土壤细菌多样性的上行调控更多来自于其促进草本植物多样性之后的生物级联效应[7],即:间接效应,另一方面说明,草本植物多样性是塑造大金山岛土壤细菌多样性的主要驱动力。本研究中,落叶木本植物占比与草本物种多样性存在积极的关联格局,其主要原因是,随着落叶木本植物占比的增多,尤其在非生长旺季,群落内的透光性增强,这样可促进阳性草本植物的适合度,从而增加草本植物物种多样性[28—29]。本文在调查中发现,相较于青冈群落样地1,2,3,落叶木本植物占比较高的野桐群落1,3及小叶女贞群落的阳性植物居多,例如络石、酢浆草与求米草等。同时本文也发现,木本植物物种多样性的增加会显著促进与草本植物物种多样性,这也从另一个侧面说明,木本植物物种多样性的增加会增加林下光资源的异质性,从而有利于维持光适应策略更为多样的草本植物[8, 47]。那么,随着草本物种多样性的增加,土壤养分资源消耗过快,加剧了不同细菌种类之间的养分竞争[48],使得细菌多样性下降,因此草本物种多样性与土壤细菌多样性间表现出了负向关系。
落叶木本植物比例增加所导致的林下草本植物繁荣不仅加剧了不同土壤细菌间的养分资源竞争,也可能会加剧土壤动物与细菌的养分竞争[11, 34]。本研究发现,土壤动物与细菌多样性间存在显著的负相关关系,这一格局说明,土壤动物与细菌间不仅仅存在互利作用,即正相关关系,也存在资源竞争格局[7, 11]。但在这里需要指出,土壤动物对细菌多样性的抑制作用也可能与土壤动物与细菌的食性有关。由于本文土壤动物的鉴定水平仅为目水平,无法深入探讨不同食性的土壤动物与细菌之间的资源竞争格局。综上,落叶木本植物占比的增加不仅会显著改变群落内部的光线供给性,从而影响草本物种多样性格局[28],更会通过改变凋落物基质多样性而调控土壤养分周转和供给等功能,从而改变土壤动物和细菌多样性格局[10, 30—31]。
4 结论
本研究表明,落叶木本植物占比是大金山岛植被系统不同营养级生物多样性的主要塑造者,其通过选择效应显著抑制土壤动物多样性,但会通过增加林下透光性而提升草本多样性,并通过促进草本多样性而间接减少土壤细菌多样性。该研究结果有助于深入理解中亚热带北缘森林生态系统多营养级生物多样性的维持机制。需要指出,考虑到本研究土壤动物的鉴定水平,加之本研究结构方程模型分析显示,土壤速效养分含量并未对土壤动物和细菌多样性产生直接和间接影响,而只影响落叶木本植物占比,因此,未来有关土壤动物和细菌多样性负向关系的维持机制需要使用分类水平更高的多营养级生物多样性数据来进行更深入的探索。最后,本研究仅从数量关系角度揭示了各营养级生物多样性间(alpha多样性)的关联性,未来应考虑利用控制实验等手段确认落叶木本植物占比在森林生物多样性(alpha, beta多样性)维持中的生态机制。
