不同触地习惯对跑步时足弓、跖趾关节在体运动学及足底筋膜长度的影响
2023-09-02孙晓乐叶东强张燊张希妮傅维杰
孙晓乐 ,叶东强,张燊,张希妮,傅维杰*
为了更好地满足走、跑的需要,人类足部形态和结构出现了适应性变化,包括大脚趾内收、内侧纵弓及足底筋膜(plantar fascia,PF)的形成(Holowka et al., 2018;Mc-Nutt et al., 2018)。运动时,上述结构具有承担载荷、缓冲和稳定足部杠杆的重要作用(Welte et al., 2021)。其中,PF是三者发挥作用的结构基础,其横跨足弓,通过包裹跖趾关节(metatarsophalangeal joint,MTPJ)连接跟骨与5个近节趾骨(Bojsen-Moller et al., 1976)。运动时,PF在足弓弹簧(arch-spring)和绞盘效应(windlass mechanism)2种功能中具有重要作用。PF和足弓类似弹簧,在步态支撑早、中期,足弓受压而长度增加,PF随着足弓增长而被牵拉的同时可储存弹性能(孙晓乐 等, 2022)。在支撑中后期,足弓发生回弹,PF缩短,同时释放弹性能。这种压缩-回弹的过程被称为足弓弹簧机制(张燊 等, 2018; Welte et al.,2018)。此外,PF具有绞盘一样的功能,在步态的蹬伸阶段,MTPJ伸展使PF缠绕趾骨头,缩短跟骨与跖骨的距离(Fuller, 2000; Sarrafian, 1987),从而抬高并缩短纵弓,同时配合释放弹性能(Bolgla et al., 2004;Hicks, 1954)。由此可知,足弓、MTPJ和PF的运动是密不可分、相互影响的。
足底筋膜炎约占跑步损伤的10%(朱迪 等, 2015;Taunton et al., 2002)。了解足底筋膜炎的损伤机制,是减少损伤发生的前提。研究普遍认为,垂直地面反作用力(vertical ground reaction force, vGRF)的冲击负载(Van der Worp et al., 2016; Zadpoor et al., 2011)及足部肌肉力量不足(Cheung et al., 2016; Latey et al., 2014; Warren et al., 1987)是诱发足底筋膜炎的重要原因。因此,有可能降低vGRF冲击峰值的前掌着地(forefoot strike, FFS)跑步触地方式受到较多的关注(姜嘉怿 等, 2018;梅齐昌等, 2015;Crowell et al., 2011; Shih et al., 2013; Williams et al., 2012)。目前,有关不同跑步触地方式对PF和足弓影响的研究结果不一:有研究认为,FFS会增加足底内附肌的激活程度,可能会减少纵弓过度压缩以及PF的应变(McDonald et al., 2016);而Chen等(2019)和Lieberman等(2010)发现,相比后跟着地(rearfoot strike, RFS)的跑步触地方式,FFS时足弓形变较大,并拉伸PF使其过度负载。因此,针对不同跑步触地方式对PF和足弓的影响亟待进一步探究。
PF、足弓以及MTPJ位于足部皮肤深层,因此很难精确动态追踪运动过程中其运动学变化。现有研究大多采用运动捕捉技术(Caravaggi et al., 2009; McDonald et al.,2016)、超声技术(Caravaggi et al., 2009; Wager et al.,2016)、尸体实验(Erdemir et al., 2004;Fessel et al., 2014)和单平面透视技术(Fessel et al., 2014; Gefen, 2003),但无法了解运动过程中在体足部骨骼的三维运动。近年来,双平面荧光透视成像系统(dual fluoroscopic imaging system, DFIS)开始应用于运动分析和康复医学领域。DFIS不受皮肤、软组织伪影的影响,非侵入,对运动无干扰、精度高、重复性高,为精准分析骨/关节真实的在体运动提供了全新的途径(叶东强 等, 2021;张翠 等, 2019)。
因此,本研究采用高速DFIS探究不同跑步触地方式对足弓、MTPJ和PF运动的影响,深入分析跑步时的足弓、MTPJ和PF功能。研究假设:相比RFS跑者,FFS跑者的足弓和PF的形变量更大。
1 研究对象与方法
1.1 研究对象
选取30名健康男性跑者(表1),其中习惯穿鞋FFS和RFS跑者各15人(Kelly et al., 2017; Perl et al., 2012)。跑步触地方式依据DFIS所采集的足部荧光图像(Welte et al.,2021)以及GRF数据进行确定。要求受试者近4周的周跑量>20 km,近3个月内无下肢损伤史,优势侧腿均为右腿。本研究已通过上海体育大学伦理委员会批准,所有受试者在实验前需熟悉实验流程并签署知情同意书。
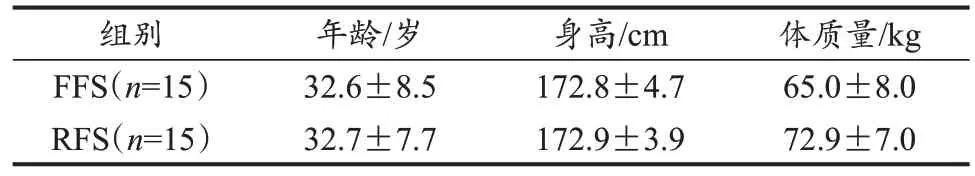
表1 受试者基本情况Table 1 Basic Information of Subjects
1.2 实验仪器
1.2.1 跖趾关节力量测试仪
采用自研的跖趾关节力量测试仪(刘宇 等,2015)采集趾屈肌力。该测试仪由底盘、踏板、座椅、拉力传感器构成。前期研究已对其进行了重测信度的检验,组间相关系数为0.874(Zhang et al., 2019)。
1.2.2 X线电子计算机断层扫描装置(computed tomography,CT)
采用64排128层CT机(SOMATOM, Siemens AG,德国)拍摄受试者仰卧中立位右侧足部的断面图像。层厚层距设为0.6 mm,体素的长、宽、高分别为0.488、0.488和0.625 mm,图像分辨率为512×512 dpi。
1.2.3 高速DFIS
高速DFIS由2组高速荧光成像系统(图1)构成,分别为2个产生X光的发射器与接收X光的图像增强器。发射器与增强器间的距离分别为131.8 cm和138.5 cm,增强器间的夹角为120°。X光发射器采用单脉冲模式(60 KV,63 mA),曝光速度为1/1 000 s,图像分辨率为1 024×1 024 dpi。采样频率为100 Hz。

图1 实验仪器设置Figure 1. Experimental Set Up
1.2.4 三维测力台
采用瑞士生产的Kistler三维测力台,型号为9260AA3(50.0 cm×30.0 cm×0.5 cm)。采集受试者跑步时的GRF数据,采样频率为2 000 Hz。
1.2.5 光栅计时系统
采用Witty-Manual光栅计时系统(Microgate,意大利),测量受试者通过跑道的时间来计算速度,从而控制跑速。
1.2.6 光栅传感器
采用GJ-2004光栅传感器,同步触发测力台和DFIS,最大限度减少拍摄时间以减少X光辐射量。
1.2.7 实验用鞋
本研究选取了一款常见的慢跑鞋作为实验用鞋,鞋中底由TPU和EVA构成,该鞋的前掌和后跟差为6 mm,单只42码鞋质量为290 g,且无足弓支撑结构。
1.3 实验流程
1.3.1 趾屈肌力测试
采用跖趾关节力量测试仪测试最大的趾屈肌力。测试过程中要求受试者坐在椅子上,固定膝关节、足前掌、足跟和足踝。测试期间,要求受试者双手抱胸尽力屈曲MTPJ以按压上抬30°的踏板(图2)。

图2 趾屈肌力测试示意图Figure 2. Schematic Diagram of Toe Flexor Strength Test
1.3.2 足部CT扫描
足部CT扫描时,受试者保持仰卧位,采用足部支具将足部固定于中立位。扫描范围从踝关节以上10 cm沿小腿横断面向下至离开足部支具结束。将扫描的图片保存为DICOM格式,用于足骨骼模型的重建。
1.3.3 高速DFIS系统调试
调整发射器与增强器的位置和角度,设置电压、电流以及曝光时间等拍摄参数,并采集孔阵钢盘和立方体标定架图像。采用XMALab 2.0.1(布朗大学,美国)对孔阵钢盘图像进行校正,根据畸变校正样条函数的算法(Fantozzi et al., 2003),校正每张足部荧光图像。立方体标定架图像用于在虚拟环境中重建三维空间。
1.3.4 跑步测试
受试者需更换实验用鞋和短裤,以3 m/s的跑速在跑步机上充分热身。热身后,实验人员为其讲解动作要求并进行示范,要求受试者熟练完成跑步动作。测试时,受试者采用自选的跑步触地习惯以3 m/s(±5%)的跑速通过测试跑道(710 cm×60 cm×47 cm),同时其身体会阻断光栅红外线而触发同步装置,启动DFIS和测力台采集受试者右脚的荧光图像及GRF数据。以右足踏上测力台标记位置(图1)为准,确保采集整个支撑期(从右足触地至离地)的足踝图像数据。为了最大限度减少X光的辐射量,采集1次有效数据(Welte et al., 2021)。以vGRF>15 N定义为足触地(Welte et al., 2018)。
1.4 数据处理
将足骨CT图像导入到Mimics 21.0软件(Materialise,比利时),采用图像灰度阈值分割和套索工具,建立跟骨、第一跖骨以及第一近节趾骨的骨骼模型。
将XMALab软件中生成的环境文件导入到Rhinoceros 6.0软件(Robert McNeel,美国)中,还原X光发射器和增强器的位置,而后导入校正后的荧光图像及骨骼模型。根据惯性解剖坐标系建立跟骨、第一跖骨以及第一近节趾骨坐标系,原点位于骨的质心,X-Y-Z轴与惯性矩张量的主轴对齐,并重新标记坐标轴,X、Y和Z轴分别代表内外、前后和上下方向。参考Welte等(2021)的研究,将PF简化为连接其起止点与籽骨接触点的2条纤维。起点位于跟骨内侧结节1/5处的2点,止点为第一趾骨内侧和外侧的2点,籽骨点位于内外侧籽骨最下端的2点。连接起点与籽骨点、籽骨点与止点,2条线段的平均长度即为PF长度(图3)。随后进行3D-2D配准,在三维虚拟环境中对骨进行平移和旋转操作,直到骨骼模型的投影轮廓与荧光骨骼轮廓相匹配(图4)。

图3 足底筋膜模型及坐标系Figure 3. Plantar Fascia Model and Coordinate System

图4 DFIS数据采集和处理Figure 4. DFIS Data Collection and Processing
1.5 实验参数
采用Rhinoceros软件中的生物力学插件计算跑步支撑期每帧足弓(第一跖骨相对跟骨的运动)、1stMTPJ(第一近节趾骨相对第一跖骨的运动)的6DOF数据(Welte et al., 2021),包括内外(X轴)、前后(Y轴)和上下(Z轴)3个方向的平移,以及屈伸(X轴)、内外翻(Y轴)和内外旋(Z轴)3个方向的旋转。其中,正值表示向外、向前和向上平移,以及伸、内翻和内旋;负值表示向内、向后和向下平移,以及屈、外翻和外旋。
对足弓、1stMTPJ的6DOF以及PF长度数据进行了滤波(20 Hz)以及时间标准化处理。同时,将整个支撑期分为支撑早期(0~20%)、支撑中期(20%~55%)、蹬伸期(55%~85%)和离地期(85%~100%)4个时期(Welte et al., 2021)。足弓6DOF、PF长度数据与足部静态中立位时足弓6DOF、PF的长度进行标准化处理。选取的参数包括:足弓、1stMTPJ在内外、前后和上下方向的平移距离,屈伸、内外翻和内外旋角度以及4个时期、整个支撑期的活动幅度;PF应变、功率、长度最大值以及最小值;其中,PF应变(Gefen, 2003)和功率(McDonald et al., 2016)计算公式如下:
1.6 统计方法
所得的参数值均采用平均值±标准差表示。采用Shapiro-Wilk检验正态分布,同时利用独立样本t检验(SPSS 20.0)比较不同跑步触地习惯对足弓、1stMTPJ的6DOF数据和PF长度的影响。显著性水平设定为0.05。
2 结果
2.1 趾屈肌力
FFS跑者的最大趾屈肌力显著大于RFS跑者[(3.45±1.05) N/kg vs (2.52±1.20) N/kg,P<0.05]。
2.2 足弓
本研究统计了每间隔10%时刻的数据,以及蹬伸期分隔点55%和85%,共13个时刻的数据。从足弓的6DOF曲线(图5)可发现,在支撑早、中期,第一跖骨相对跟骨先向外、向前和向上移动,随后在蹬伸初期向内、向后和向下运动,离地期回归中立位状态。转动状态下,跖骨相对跟骨先背屈、外展运动,在蹬伸初期达到最大,而后进行跖屈和内收运动。足弓在内外翻方向的运动幅度较小,呈现缓慢增加的趋势。
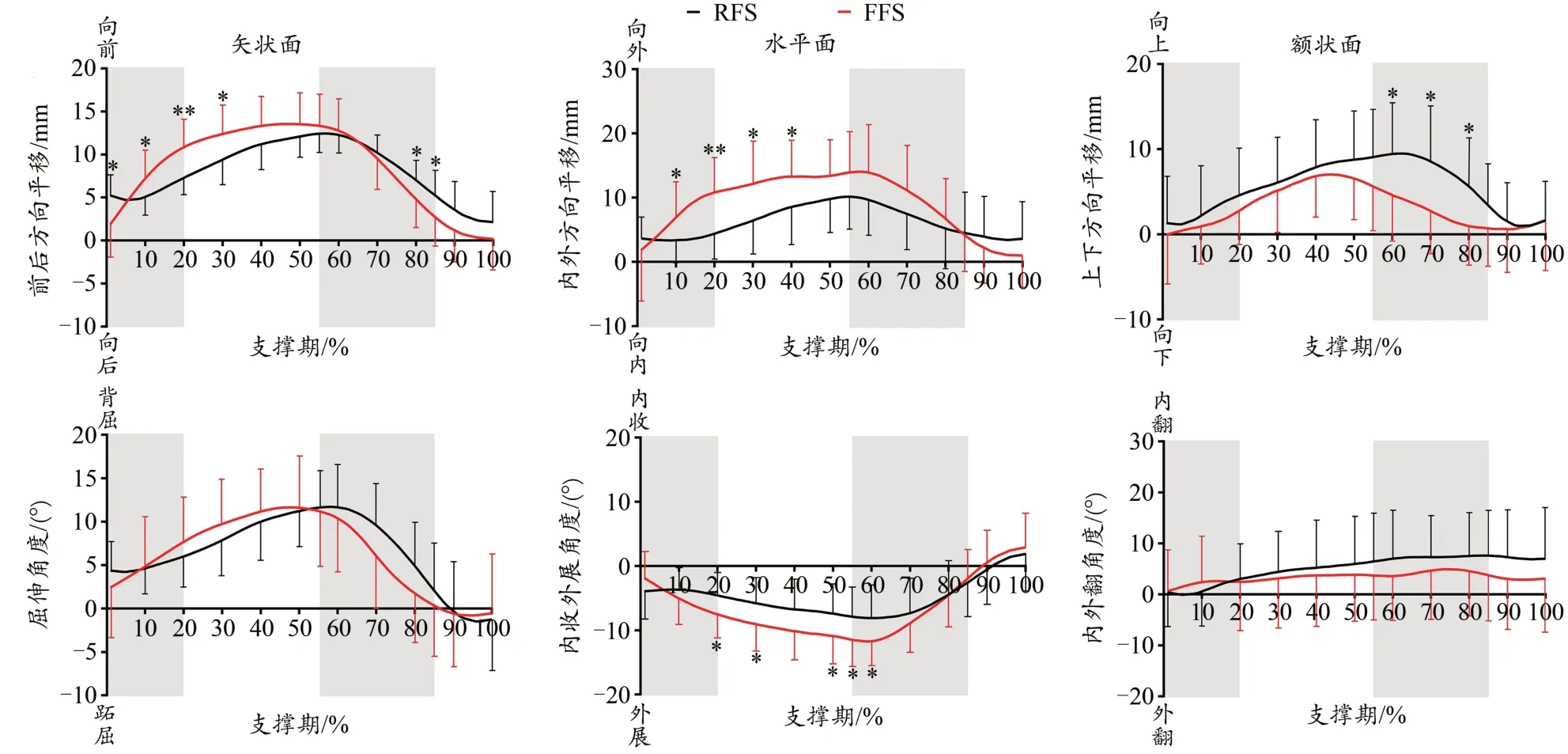
图5 不同跑步触地习惯对跑步支撑期足弓6DOF运动的影响Figure 5. Effects of Different Foot Strike Patterns on the Arch 6DOF Data during the Stance Phase of Running
平移状态下,FFS时跖骨相对跟骨向外的最大平移(P=0.011),以及支撑早期(P<0.001)、整个支撑期的(P<0.001)内外平移幅度以及支撑早期(P<0.001)、蹬伸期(P=0.005)和支撑期(P=0.004)的前后平移幅度显著大于RFS。转动状态下,FFS时跖骨相对跟骨支撑早期的屈伸活动度(P=0.003)、支撑早期(P=0.002)和蹬伸期(P=0.014)的外展活动度、最大外展角(P=0.023)显著大于RFS(图6)。

图6 不同跑步触地习惯对跑步支撑期足弓6DOF运动幅度和特征值的影响Figure 6. Effects of Different Foot Strike Patterns on the Range of Motion of Arch 6DOF Data and Characteristic Values during the Stance Phase of Running
2.3 第一跖趾关节
由1stMTPJ 6DOF运动曲线(图7)可知,平移状态下,趾骨相对跖骨先向内、向前和向下运动,在支撑中期达到最大值,随后开始向外、向后和向上平移。转动状态下,趾骨相对跖骨先屈和外旋运动,随后在蹬伸期进行伸展和外翻运动,其伸展运动与向上平移运动趋势是一致的。

图7 不同跑步触地习惯对跑步支撑期1st MTPJ 6DOF数据的影响Figure 7. Effects of Different Foot Strike Patterns on the 1st MTPJ 6DOF Data during the Stance Phase of Running
平移状态下,FFS时趾骨相对跖骨在支撑早期(P=0.009)、离地期(P=0.005)的前后平移幅度,支撑中期(P=0.001)、离地期(P=0.015)的上下平移幅度以及向上平移最大值(P=0.022)均显著大于RFS。转动状态下,在支撑早期(P=0.004)、中期(P=0.001)和离地期(P=0.012)的屈伸活动度,离地期(P=0.007)和支撑期(P=0.005)的外翻活动度,以及最大外翻角(P=0.048)均显著大于RFS(图8)。
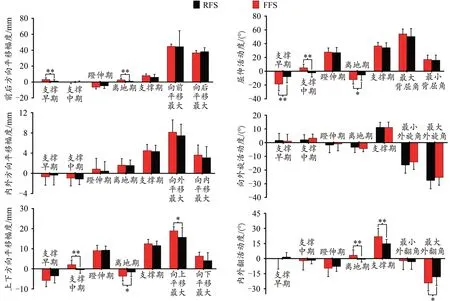
图8 不同跑步触地习惯对跑步支撑期1st MTPJ 6DOF运动幅度和特征值的影响Figure 8. Effects of Different Foot Strike Patterns on the Range of Motion of the 1st MTPJ 6DOF Data and Characteristic Values during the Stance Phase of Running
2.4 足底筋膜
在跑步支撑期,PF的应变先增加,蹬伸期达到最大值,而后逐渐减小(图9)。与RFS相比,FFS条件下支撑早期(P<0.001)、蹬伸期(P<0.001)以及支撑期(P=0.01)PF的长度变化量、最大应变显著更大(P=0.01)。此外,在支撑早期和蹬伸期,FFS条件下PF的功率显著大于RFS(图10)。
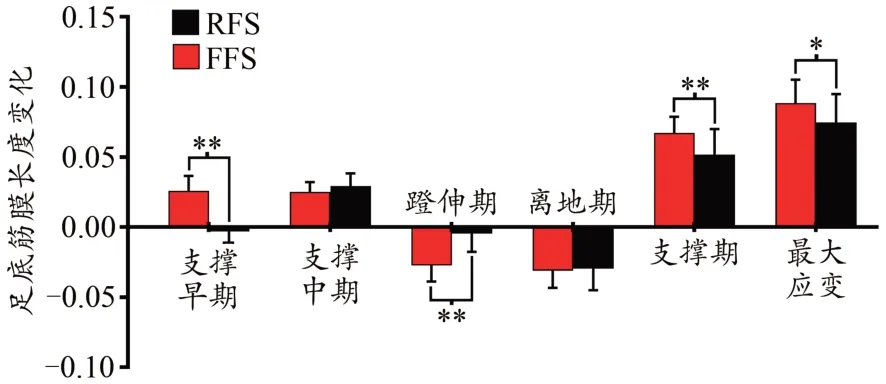
图9 不同跑步触地习惯对跑步支撑期足底筋膜应变和功率的影响Figure 9. Effects of Different Foot Strike Patterns on the Stain and Power of PF during the Stance Phase of Running
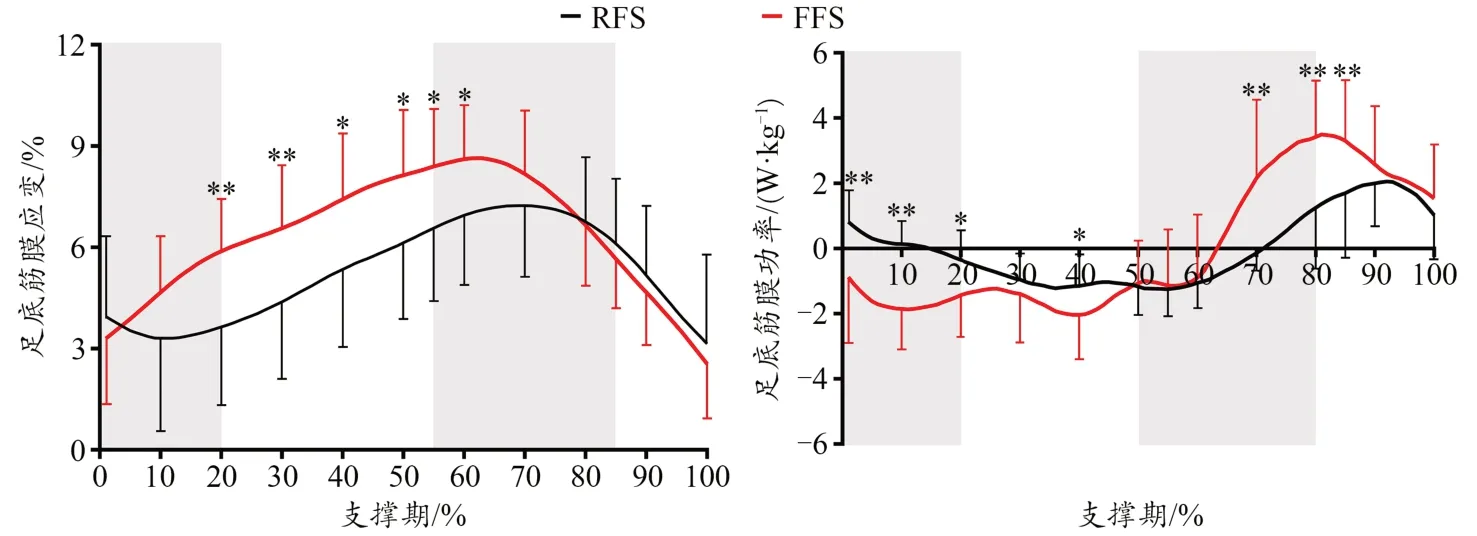
图10 不同跑步触地习惯对跑步支撑期足底筋膜长度变化量、最大长度和应变的影响Figure 10. Effects of Different Foot Strike Patterns on the Length Variation, Maximum Length and Strain of PF during the Stance Phase of Running
3 讨论
本研究假设得到验证,FFS显著增加了足弓(内外、前后、屈伸和外展活动度)和1stMTPJ(前后、上下、屈伸和外旋活动度)6DOF的变化以及PF的应变和功率,表明FFS可能强化了足弓和PF的力学特性。此外,FFS跑者的趾屈肌力显著大于RFS跑者。
本研究发现,FFS跑者的趾屈肌力显著大于RFS跑者,表明FFS使足部发生了适应性肌肉力量增强。有研究认为,足部肌力不足是足底筋膜炎发生的重要诱因,因足部肌肉不能在运动中控制足弓的形变,进而导致PF应变的过度增加(Cheung et al., 2016; Latey et al., 2014; Warren et al., 1987)。同时,趾屈肌力的增强可协调足后跟离地,使跑者获得较大的支撑反作用力,进而促使身体快速蹬离地面,潜在提高了跑步经济性(Maden et al., 2021)。
跑步时,足部纵弓类似弹簧,在支撑早、中期受压将机械能储存于足弓的弹性结构中,随后在蹬伸期弹性结构缩短,足弓释放能量(Kelly et al., 2017; McDonald et al., 2016)。Ker等(1987)通过尸体研究发现,足弓可储存并释放足够的能量以节省跑步代谢能,该观点在在体研究中也得到了证实(Stearne et al., 2016)。有研究指出,纵弓约提供跑步(跑速为4.5 m/s)所耗机械能的17%(Ker et al., 1987)。本研究发现,相对RFS,FFS增加了足弓在矢状面上的运动幅度,如支撑早期、蹬伸期和整个支撑期的前后平移幅度,以及支撑早期屈伸活动度显著增加,这表明FFS时足弓矢状面上的压缩量更大,而较大的足弓形变会储存更多的弹性能,同时衰减冲击。该结果得到了前人研究的支持(Perl et al., 2012),FFS条件下足弓形变更大,可能由于FFS时从触地开始足弓受压长度增大,而RFS时从全掌着地足弓开始负载受压。负载阶段,PF牵拉跟骨与跖骨,防止足弓过度形变而坍塌,并通过形变吸收部分足部冲击能量(Gefen, 2003;Snow et al., 1995)。在随后的蹬伸阶段,MTPJ的伸展产生绞盘效应,PF缠绕趾骨牵拉跟骨向跖骨运动,足弓长度缩短(Bolgla et al.,2004;Hicks, 1954)。
MTPJ作为下肢最末端关节,其伸展运动是通过踝关节跖屈肌配合足趾屈肌在远固定条件下收缩完成的(傅维杰 等, 2013),MTPJ屈曲角度的改变对跑步动作尤其是支撑后期的蹬伸效果产生重要影响(张发宁 等, 2022;Goldmann et al., 2011; Willwacher et al., 2013)。本研究发现,在整个支撑期MTPJ一直处于伸展状态,其运动趋势与前人研究结果一致(Welte et al., 2021),在此过程中,MTPJ主要以吸收机械能为主,基本不释放能量(Roy et al.,2006)。RFS条件下,支撑早、中期和离地期MTPJ的屈伸活动度和上下位移幅度更小(张发宁 等, 2021),较小的活动度可减小能量损失(Roy et al., 2006),而整个支撑期MTPJ的活动度无显著差异。因此,不同触地习惯对MTPJ能量吸收的影响还亟待进一步探究。本研究发现,在蹬伸期,FFS条件下MTPJ屈伸角度和向上平移值更大,这表明MTPJ伸展会进一步拉伸PF使其长度增大,绞盘效应更显著,同时也提高了MTPJ蹬伸力的传递。
对于PF,本研究发现,从触地开始PF应变呈现先增加后减小的趋势,表明PF的特性符合弹性结构的伸长-缩短期(stretch-shorten cycle,SSC)模式,在支撑早、中期PF储存弹性能(功率为负),在蹬伸期和离地期释放弹性能(功率为正)。PF这一特性可有效发挥“弹簧”作用,在不同负载条件下调节足部的弹性功能(McDonald et al., 2016)。此外,足弓矢状面的形变以及MTPJ的伸展也会影响PF的应变(Caravaggi et al., 2009; Flanigan et al., 2007)。本研究中,FFS和RFS跑者PF的最大应变分别为8.9%和7.5%。PF的最大应变决定了弹性能的效益(McDonald et al.,2016),FFS时PF最大应变显著大于RFS,这表明FFS跑者会从PF中获得额外的运动表现优势(performance benefit),从而提高运动表现。而Gefen等(2003)发现,从足全掌触地至离地阶段PF的应变为9%~12%,最大应变值的差异可能是由于受试者性别、测试动作的差异等引起的。本研究发现,在支撑期的前10%,RFS时PF出现缩短,而FFS时PF持续增长。FFS时PF这种“预负载”(preloading)会改变其张力和足弓长度(Iwanuma et al., 2011),同时可能有助于蹬伸期身体的向前推进(Pataky et al., 2008)。因这种“预负载”会减少PF胶原组织的卷曲现象,从而更早地强化足弓刚度,有助于蹬伸期足部传递更大的蹬伸力至地面(Caravaggi et al., 2009)。
本研究采用高速DFIS精准分析了不同跑步触地习惯下足弓、MTPJ以及PF的在体生物力学特性,但也存在一定的局限性:1)将PF简化为2条纤维,模型远端的部分线段会穿过籽骨内部,而实际上PF是起于跟骨结节缠绕籽骨,止于近端趾骨,并未穿过籽骨;2)本研究主要分析了足弓和MTPJ的运动学,未深入探究动力学数据;3)受试者均为健康男性跑者,不同性别、年龄以及足底筋膜炎患者的运动表现有待进一步探究。
4 结论
本研究发现,相比于后跟着地跑者,前掌着地跑者在跑步支撑阶段足弓的长度变化,第一跖趾关节的前后、上下和屈伸活动度显著增加,这表明前掌着地跑者足弓储存和释放的能量更多,跖趾关节的蹬伸效果更好。同时足底筋膜的应变以及功率显著增加,表明前掌着地跑者足底筋膜储存和释放的弹性能更多,具有更强的足底筋膜力学特性。此外,前掌着地跑者也具有更大的跖趾关节屈肌力量。
