碳氮磷化学计量比对土壤有机碳矿化激发效应的影响*
2023-08-24张睿媛秦树平胡春胜
张睿媛 ,袁 丹 ,秦树平 ,胡春胜**
(1.中国科学院遗传与发育生物学研究所农业资源研究中心/河北省土壤生态学重点实验室/中国科学院农业水资源重点实验室 石家庄 050022;2.中国科学院大学 北京 100049)
土壤有机碳库是陆地生态系统最大的碳库,其量级大约是大气碳库的3 倍(2400 Gt C vs.800 Gt C)[1]。土壤有机碳的矿化会释放大量CO2进入大气,其量级约是人为工业排放的7 倍[2]。由于土壤有机碳库规模巨大,其动态变化将对全球气候变化产生巨大影响[3]。因此,提升土壤有机碳库的稳定对于缓解全球气候变暖至关重要。
激发效应(priming effects,PE)是外源养分输入导致的微生物对土壤原有有机碳(soil organic carbon,SOC)降解速率的改变[4],是影响土壤碳库动态的关键机制之一[5-6]。长期以来,激发效应一直被认为会导致土壤碳的净损失,即正激发效应,根据环境条件和土壤类型的不同,激发效应可使土壤有机碳矿化速率最高提升400%[7-8]。然而,最近研究表明激发效应的方向也可能是负的,激发效应可使土壤有机碳矿化速率减少50%[9]。激发效应是一个复杂的过程,涉及不同种类微生物,受不同环境参数的影响。土壤特性、气候、土地利用方式、外源添加物质的性质及其添加方式都会影响激发效应的方向和大小[10]。
生态计量学结合生态学和化学计量学的基本原理,是研究生态系统养分循环及其生态效应的重要工具。碳氮磷作为生命的重要元素,在物质循环和能量流动中起着重要作用[11-12]。在土壤中,碳氮磷不仅是生物生长的基础,也是土壤有机质的关键组成元素[13]。因此,营养元素碳氮磷的含量及其化学计量比显著影响外源物质在土壤中的周转以及土壤原有有机质的矿化[14]。此外,碳氮磷化学计量比还调控了生态系统中土壤微生物群落的生长和活动。根据 “土壤微生物碳泵”理论[15],调控土壤微生物可利用的碳氮磷的化学计量比,在理论上可实现对土壤微生物量、群落组成与微生物周转速率进行定向调控,充分发挥微生物碳泵在提升土壤有机质方面的作用,通过“激发效应”和“续埋效应”调控土壤稳定有机碳库储量动态。
本文首先总结了碳氮磷化学计量比调控土壤激发效应的相关研究进展,接着分析了微生物作为激发效应的内源驱动力对碳氮磷化学计量的响应机制,然后总结出“共代谢” “微生物营养挖掘”和“化学计量分解”3 种有关碳氮磷化学计量比调控激发效应的机制,最后对碳氮磷化学计量比调控土壤碳矿化激发效应的研究不足与未来方向进行了总结和展望。
1 外源碳氮磷不同配施对激发效应方向与强度的影响
目前关于外源添加对土壤有机碳矿化激发效应的研究主要集中在外源碳氮配施、碳磷配施和碳氮磷配施,不同营养元素的配施对激发效应的影响也不尽相同(表1)。
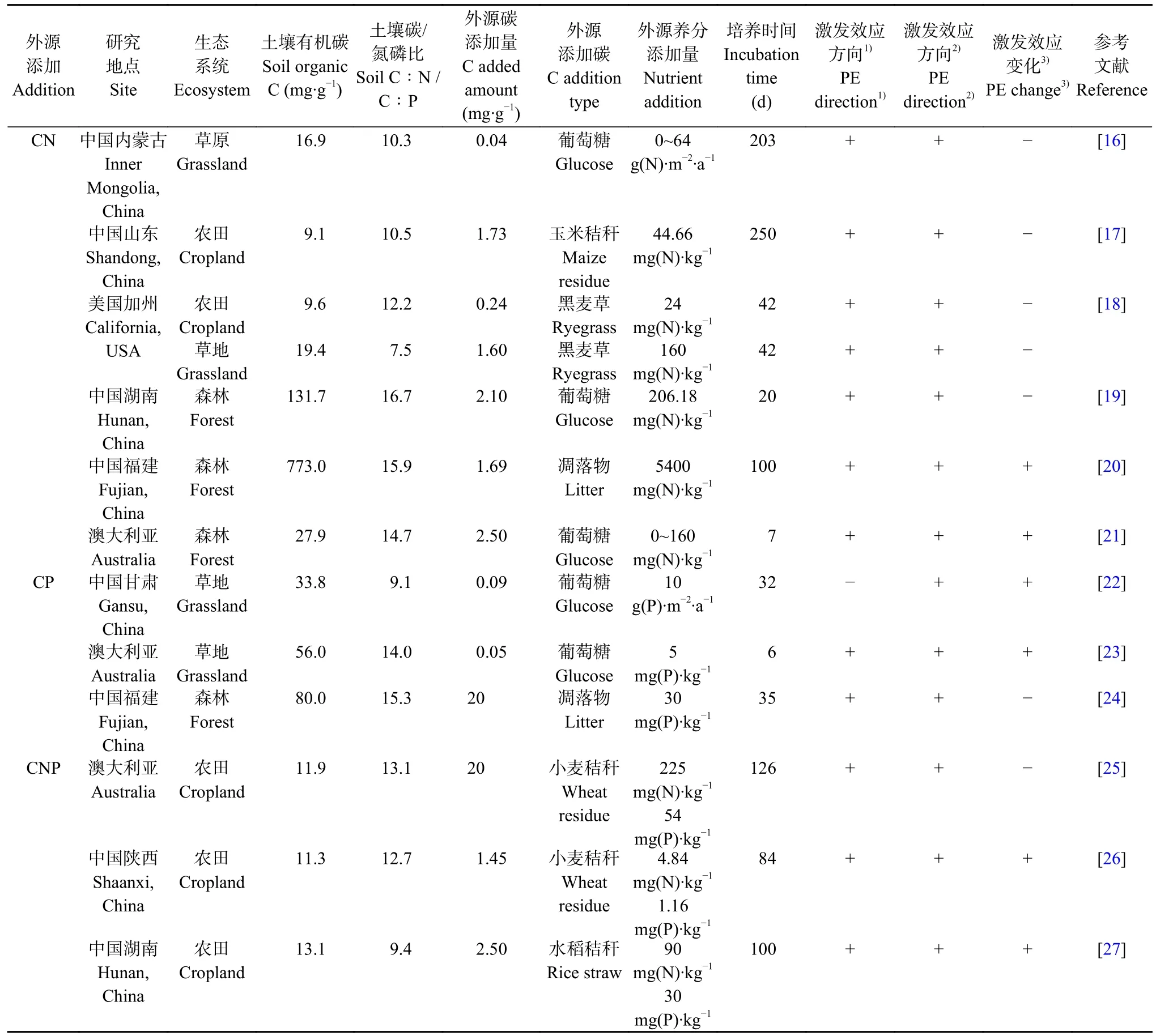
表1 外源碳氮磷输入对土壤碳矿化激发效应的影响Table 1 Effect of carbon,nitrogen and phosphorus input on the priming effects (PE) of soil carbon mineralization
1.1 外源碳氮配施对激发效应方向与强度的影响
激发效应的方向与强度和外源添加碳氮的微生物可利用性密切相关[28-30]。在较低的氮可用性条件下,外源碳氮配施使激发效应增强,微生物通过挖掘土壤中的有机碳以满足其对氮素的需求[31]。相反,在氮富集的情况下,微生物可以通过降低自身的碳氮比来响应外源养分的添加。因此,当提供低碳氮比的底物时,激发效应往往会降低,因为微生物不需要分解土壤有机质来获取氮素[29]。相关研究结果表明,外源碳氮配施下,正激发效应的大小随着氮添加量的增大而减小[16-20],这说明当土壤养分限制得到缓解时,土壤微生物不需要分解土壤有机质来满足其对氮的需求,同时减少了胞外水解酶的分泌,激发效应强度降低。但Zheng 等[21]发现在外源添加葡萄糖和氮素后,激发效应随氮添加量的增加而增加。这可能是因为氮加速了微生物内部代谢和周转,增加了分解土壤有机碳所产生的CO2;其次,外源碳氮添加所激活的微生物可以利用添加的氮产生水解酶,加速土壤难分解有机质的分解。一项全球Meta 分析表明,在微生物对本底氮挖掘占主导的土壤系统中,氮添加会降低激发效应强度,而在土壤本底氮素满足微生物需求的系统中,氮添加会增强激发效应[32]。
1.2 外源碳磷配施对激发效应方向与强度的影响
随着农业生产对磷需求的不断增加,磷肥管理成为农业系统的常见措施[33]。磷的微生物可利用性在调控土壤微生物活性方面起着关键作用,可以强烈影响土壤的碳周转[34-35]。现阶段关于磷添加对自然生态系统土壤碳周转的研究表明: 磷通过增加土壤微生物呼吸[36]、降低碳利用效率[37],或增加微生物对土壤本底碳的分解[38],促进了森林和草原土壤中的有机碳分解,这意味着磷可以调控土壤原有有机碳的分解,微生物对磷的需求诱导磷酸酶的产生[39],进而促进了土壤有机质的分解与磷的释放。外源磷添加会影响土壤真菌和细菌的群落组成[40-41],并影响碳氮磷获取酶的活性[42-43],这可能会改变土壤微生物间的群落结构,直接影响实际激发效应强度。在同一生态系统中,长期施磷土壤在外源添加碳后所产生的激发效应强度较不施磷土壤显著增加[22,44]。最近的新型双同位素标记方法(13C 和18O 示踪法)也揭示了磷可用性高的土壤比磷可用性低的土壤具有更强的激发效应[45]。除此之外,磷还可以通过影响土壤中氮的转化间接影响激发效应的强度,Mehnaz 等[23]发现,外源碳磷添加诱导了微生物生长的氮限制,使微生物通过挖掘土壤有机质中的氮来满足自身营养需求,增加了激发效应的强度。然而,在磷添加条件下,激发效应的强度则与外源碳质量有关,梅孔灿等[24]的研究结果表明,磷添加降低了低质量凋落物添加所产生的激发效应,缓解了土壤微生物的磷限制。
1.3 外源碳氮磷配施对激发效应方向与强度的影响
目前大多数研究主要聚焦在单一元素添加后土壤有机碳的分解,但在生态系统中碳氮磷通常是一起输入的[46]。Feng 等[32]的Meta 分析结果表明,68.3%的外源氮磷添加对土壤碳矿化激发效应的效应量低于零,即氮磷的添加降低了激发效应的强度。氮磷的添加可能会改变细菌、真菌的丰度和多样性,以及与碳周转相关微生物群落功能的变化,进而影响土壤有机碳矿化。具体的影响取决于土壤本底性质以及外源碳氮磷添加量。与单独输入纤维素碳相比,高量碳氮磷配施降低了30%~50%的正激发效应强度,且伴随着较低的细菌、真菌基因丰度[25];Mo 等[26]的研究结果也表明,在外源碳氮磷比例保持不变时,高量输入相较于低量输入激发效应强度显著降低,这说明微生物对本底土壤有机质的“营养挖掘”在外源养分可用性增加的情况下有所缓解。与之相反,有研究结果显示,碳氮磷配施对土壤有机碳矿化有积极或中性的影响[26,47-48],这可能是因为外源低水平的碳氮磷输入增加了土壤微生物的活性,使其分泌胞外水解酶以降解顽固的碳底物以获得自身所需的营养物质。
2 激发效应相关微生物对碳氮磷化学计量比的响应和调节机制
Kuzyakov[49]提出,激发效应是“活”有机物和“死” 有机物之间的相互作用,其中微生物是主要驱动因素。微生物通常喜欢高质量(低碳氮比)而非低质量的底物(高碳氮比)[50],因为后者需要增加胞外水解酶的分泌才能代谢[51]。外源添加高质量的底物会产生正激发效应,因为高质量底物会增加微生物的活性,额外的能量供应使微生物能够代谢原本无法接触的底物。但是,高质量的凋落物添加也可能对较老的土壤有机质分解产生负面影响,因为微生物会优先利用添加的新鲜有机物[9]。低质量凋落物添加的结果则更难预测,有研究结果表明,低质量的凋落物所引起的激发效应强度较小,因为添加的底物本身难以被微生物利用[33]。然而,其他研究结果表明,添加低质量底物会增强激发效应强度[52]。除此之外Fanin 等[53]的研究结果表明,外源添加高质量的凋落物会产生“瞬时”激发效应,当氮满足需求时迅速降低;添加低质量的凋落物会产生“延时”激发效应,当氮耗尽时,强度持续增加。
土壤有机碳周转相关微生物主要有4 种主要机制来响应资源化学计量: 1)通过非稳态行为(微生物改变其元素组成来响应资源组成的变化)调整其微生物量碳、氮、磷;2)改变特定的酶活性以获得限制性元素;3)通过调节元素利用效率(例如,碳利用效率carbon use efficiency,CUE),将超过其需求的元素矿化和排出;4)改变微生物群落组成[35]。微生物对碳氮磷需求的差异使得不同营养条件下的群落变化与碳循环的可预测变化有关。因此,需要将土壤碳周转的化学计量调控与微生物群落联系起来。
2.1 微生物量对碳氮磷化学计量比的响应
微生物可以将外源有机碳转化为微生物量并最终转化为残体物质,从而有助于土壤有机碳的稳定[54]。土壤微生物量是形成土壤有机质的重要组成部分,可调节有机物和养分的循环。虽然土壤微生物量仅占土壤有机质的一小部分,但它对生态系统中碳氮磷的循环做出了显著贡献。在农业土壤中,资源的碳氮磷化学计量相较于微生物所需的碳氮磷化学计量表现出更可变的特性[55]。因此,微生物会根据土壤营养水平改变其群落结构,支持适应资源碳氮磷化学计量的优势菌株,从而维持微生物量碳氮磷化学计量的平衡[56]。Huang 等[57]的研究发现,土壤碳氮磷化学计量与微生物量碳氮磷化学计量之间存在很强的线性正相关关系,但当土壤的原始养分状态可以满足微生物需求时,进一步的养分添加对微生物量的影响很小[58-59]。在热带和亚热带森林生态系统中,氮添加对微生物量碳氮没有显著影响,但在缺氮的温带森林生态系统中,氮添加提高了土壤微生物量碳氮的含量[60-61]。以前的研究发现,当碳限制被消除时,氮或磷是土壤微生物量的第二限制养分[62-63]。而单一的氮或磷添加通常对微生物生长没有影响,与单独秸秆还田相比,秸秆与氮磷的配施增加了微生物量碳,因为氮磷的可用性越高,微生物代谢活性也越高[64]。此外,有机肥与氮磷肥配施可以加速养分释放,从而减轻微生物群落的化学计量限制[56]。
与土壤资源碳氮磷化学计量梯度相比,微生物量碳氮比相对稳定,范围为8∶1 到12∶1[65-66]。最近的研究结果表明,与之相比,微生物量碳磷比变化范围较大,这说明微生物量磷的化学计量可塑性更大[67-68]。土壤微生物量碳磷比可以反映微生物量磷的有效性,作为磷供应受限程度的指标[69]。Fan 等[59]的研究结果表明,氮添加对土壤微生物量碳氮没有显著影响,但微生物量碳磷比显著增加,这表明施氮诱导微生物的磷需求增加。
2.2 土壤水解酶活性对碳氮磷化学计量比的响应
土壤微生物获取能量和养分的相对资源分配可以表示为碳氮磷水解酶活性的比率,通过结合生态化学计量理论和生态代谢理论来指示微生物代谢的局限性[70]。糖苷酶可以将纤维素和糖分解成溶解的有机碳,包括纤维二糖水解酶、α-葡萄糖苷酶、β-糖苷酶和β-木糖苷酶。而亮氨酸氨基肽酶和β-1,4-N-乙酰氨基葡萄糖苷酶负责通过降解蛋白质和几丁质来获取氮,磷酸酶通过水解有机磷释放出可用的无机磷。这几种水解酶已在与土壤养分循环相关的领域中得到了广泛的研究。此外,Moorhead 等[71]根据水解酶活性的比例计算酶活矢量角,更好地量化和可视化了微生物的代谢限制。多项研究表明,氮添加会显著降低土壤碳氮磷水解酶活性[72-73],从而抑制土壤中化学元素的转化以及养分的分解和释放[74]。其他研究则表明,氮添加对土壤水解酶活性有积极影响[75]。土壤碳氮磷水解酶活性对外源添加物响应的差异可能会导致酶化学计量比在氮添加后表现出不确定性[61]。此外,氮添加可以通过影响土壤酶活性来间接影响土壤微生物群落的养分限制。Xu 等[76]的Meta 分析表明在全球范围内,氮添加主要影响土壤微生物的碳磷限制,尽管氮添加增加了土壤碳磷水解酶的活性。Ma 等[77]的研究发现,森林生态系统中,磷的添加缓解了微生物的磷限制,增加了碳氮水解酶的活性。Liu 等[78]的研究结果表明,土壤水解酶活性与高营养浓度之间存在负反馈效应。
激发效应中涉及的酶活性意味着收益和成本[79],响应外源添加的酶活性变化可能导致土壤有机碳分解和激发效应的变化。然而,目前激发效应与酶活性变化之间的关系仍未可知。
2.3 微生物碳利用效率对碳氮磷化学计量比的响应
微生物碳利用效率(CUE)定义为用于微生物生长的碳相对于总碳摄取的比率,即微生物将底物碳转化为自身微生物量碳的效率[80],高碳利用效率表明微生物快速增长和较少的碳释放进入大气,低碳利用效率意味着相对较大的碳损失,较少的碳转化为微生物量[45]。养分限制是微生物碳利用效率的一个重要驱动因素,因为分解者代谢具有固定的化学计量限制[81],一般来说,增加养分可用性有望通过减少碳与养分的化学计量失衡来改善土壤微生物碳利用效率[25,80]。在养分丰富的土壤中,微生物通过减少碳与养分的化学计量失衡,对外源碳添加表现出更高的利用效率[81]。而在养分限制的土壤中,底物营养成分的差异往往决定了营养限制对微生物碳利用效率影响的大小和方向[45],例如当微生物利用高碳氮比的底物时,可能会出现碳利用效率下降,这是因为大量的碳投入导致有限的养分获取和大量的呼吸消耗[81]。然而也有研究表明,高碳氮比或对微生物碳利用效率没有影响[37,82]。Qiao 等[83]的Meta 分析结果表明,对于外源添加葡萄糖的土壤,氮配施对微生物碳利用效率没有影响,但是外源高分子化合物和植物残体添加的土壤中,氮配施分别导致微生物碳利用效率下降51%和53%。Song 等[84]的研究表明,微生物通过减少微生物碳利用效率来增加土壤激发效应强度,这是微生物对可用资源化学计量比的反馈。除此之外,微生物碳利用效率还受微生物群落组成的控制[45],因为不同的微生物群在分解和同化碳底物方面具有不同的效率。
2.4 微生物群落结构对碳氮磷化学计量比的响应
激发效应由两种功能性微生物群落负责,一种是利用顽固底物生长缓慢的K-策略者,另一种是代谢不稳定底物生长快速的r-策略者。微生物群落中r-和K-策略者相对丰度的变化是反映土壤微生物对压力源响应的有用指标[85]。K-策略者和r-策略者之间的平衡在某种程度上反映在真菌与细菌的比例(F∶B)或革兰氏阳性与革兰氏阴性菌的比例(G+∶G-)上,较高的比率被认为是K-策略者比r-策略者贡献更大[86]。Chen 等[28]的研究指出,K-策略者在分解低氮可用性的底物方面更具竞争力,而添加碳氮后的激发效应与r-策略者的主导地位有关。前人研究结果表明,在外源氮添加下,真菌生物量的减少和F∶B的降低会抑制土壤顽固碳的分解,导致葡萄糖和苯酚引起的激发效应强度降低[16,87-88]。而Bei 等[89]的研究结果中,秸秆与矿物质氮配施后,总磷脂脂肪酸(PLFA)的F∶B 值增加,13C-PLFA 分析进一步表明,矿质氮添加减少了秸秆来源的细菌,增加了秸秆来源的真菌和F∶B。先前的结果表明,磷有效性对土壤有机碳矿化的影响主要是由真菌通过改变群落结构和释放胞外酶实现的,磷添加导致K-策略者(特别是关键真菌分类群)的丰度和多样性的改变[90-91]。细菌和真菌在营养丰富和营养不足的土壤环境中具有不同的偏好[7]。真菌的碳氮比高于细菌,因此真菌在低营养条件下更活跃,因为它们的氮需求较低,碳利用效率较高[92]。但在营养丰富的情况下,例如外源秸秆与氮肥的配施,则降低了大多数真菌的丰度和多样性,但增加了细菌群(例如放线菌)的丰度[93-94]。低养分含量导致细菌和真菌物种间和内部对资源的激烈竞争,秸秆与氮配施通过减少微生物群落之间的竞争来改变负关联的共生模式[95]。迄今为止,细菌和真菌群体之间的竞争以及在农业生态系统有机质矿化过程中的主导顺序还存在相当大的不确定性[96-97]。
土壤有机碳周转相关微生物主要通过以上4 种机制来响应资源化学计量。虽然上述的4 种机制是并行运作的,但它们在空间、时间和生态系统尺度上的贡献仍未可知。土壤微生物量碳氮磷比率变化的相对灵活性限制了其非稳态行为,并且微生物群落在碳与营养比不同环境中的增长,不能仅仅通过非稳态行为来实现。因此,微生物元素利用效率和胞外酶分泌的补偿性调节有助于微生物响应资源的化学计量变化。
3 碳氮磷化学计量比调控土壤激发效应的假说
碳氮磷化学计量比对土壤碳矿化激发效应的影响是外源养分(碳氮磷)与土壤微生物、土壤本底有机质共同作用的结果,目前的研究主要从“共代谢”假说、“营养挖掘”假说、“化学计量分解”假说等方面来解释土壤碳矿化激发效应的强度与方向。
3.1 “共代谢”假说
“共代谢”是指土壤微生物在利用外源添加的易分解碳作为自身初始能量后,通过分泌特异性胞外水解酶降解同一介质中稳定性较高的有机化合物。土壤微生物种群的一个重要特征是它们对底物降解的酶特异性,即不同微生物分泌不同的酶来降解不同的有机物[51]。土壤微生物在分解顽固有机物的过程中,所分泌的水解酶同时也会分解土壤有机质。所以,外源添加物的生化多样性越高,微生物产生的胞外水解酶种类也就越丰富,所引起的激发效应也越明显[98]。Wu 等[99]的研究表明外源不稳定碳的添加,增加了微生物磷脂脂肪酸的含量以及β-1,4-葡萄糖苷酶和纤维二糖水解酶的活性,加速了土壤有机碳的矿化并产生了正的激发效应,且正激发的产生与微生物量和胞外水解酶的增加同步。Song 等[84]的研究结果表明在外源葡萄糖添加后,β-糖苷酶/(β-1,4-N-乙酰氨基葡萄糖苷酶+亮氨酸氨基肽酶)和β-糖苷酶/磷酸酶的比值增加,这将导致更多的碳水解酶进入土壤中,更多的水解酶在利用外源葡萄糖的同时增加了与本底有机碳接触的可能,导致更强烈的正激发效应。Börger 等[100]的研究结果表明,外源添加碳氮比相差较大的凋落物后,均产生了强烈的正激发效应,且碳氮比不同的两种凋落物(碳氮比为25.5和57.8)并不影响激发效应的响应,这说明此时的激发效应是由于微生物的“共代谢”。Shahbaz 等[101]的研究表明,在外源凋落物添加的培养试验中,培养后期激发效应的强度超过凋落物的分解强度,且土壤有机质衍生碳在微生物量中的增加与激发效应和酶活性的增加均显著相关,这表明实际激发效应的发生可能是由于微生物的变化,即从快速生长(r-策略者)变化为缓慢生长的种群(K-策略者)。由此可见,“共代谢”普遍存在于激发效应中。
3.2 “营养挖掘”假说
“微生物氮挖掘”假说是指在低氮可用性条件下,生长缓慢的K-策略者利用不稳定的碳作为能量来源,分解含氮的顽固有机物以此来获取自身所需要的氮素[30],产生正的激发效应。这意味着土壤氮限制有助于微生物分解顽固的土壤有机质以获取氮。Meier 等[102]的研究表明,过量的碳施入导致土壤微生物的养分限制(即氮限制),从而驱使它们利用不稳定碳作为能量来源,分泌更多的酶(氧化酶和氮水解酶)分解土壤本底有机质以获取有限的氮资源,进而产生了正的激发效应。Song 等[84]的研究表明,在添加葡萄糖后,土壤中的总溶解性有机氮显著增加,且β-糖苷酶与过氧化物酶的比值降低,这说明外源葡萄糖的添加诱导微生物利用土壤本底中的可用性氮有机质,促进土壤有机碳的矿化。“氮挖掘”假说强调的是微生物氮限制在激发效应过程中的主导作用,但是微生物的生长和繁殖也离不开其他营养元素,如近两年提出的“磷挖掘”假说[39]。
3.3 “化学计量分解”假说
“微生物化学计量分解”假说表明,当土壤本底和外源营养输入与微生物化学计量需求相匹配时,生长快速的微生物(r-策略者)活性最高,对土壤有机质的分解速率最快[28]。因此,“微生物化学计量分解”通常在微生物没有明显营养限制的情况下占主导地位[20]。“微生物化学计量分解”假设微生物化学计量是一种驱动力,这意味着所有微生物群落的生长增加,包括β-变形杆菌和寡营养菌(如酸杆菌和真菌)。高营养可用性可能会加速外源添加的新鲜有机物和本底有机质的矿化,从而延长激发效应的时间[7,28-30]。Zhang 等[20]的研究观察到,凋落物配施氮与单独凋落物添加相比引起了更高的激发效应,且微生物量和胞外水解酶活性均高于不加氮处理,这表明当外源添加满足微生物对养分的需求时,微生物活性最高。Bei 等[89]的研究结果表明,秸秆配施氮肥对微生物量碳无影响,但促进了激发效应的强度、秸秆分解速率和胞外水解酶活性,这与“微生物化学计量分解”假说一致。
“共代谢” “营养挖掘”和“化学计量分解”假说可以单独或组合运行(图1),具体取决于不同空间和时间尺度上土壤的碳氮磷可用性。最近的研究表明,主导机制随有机碳化合物、添加方式和分解阶段而变化[99-103],Chen 等[28]认为在外源碳氮添加后,“化学计量分解”和“共代谢”驱动初始激发效应阶段,而“微生物氮挖掘”驱动激发效应的后期阶段。其他研究则表明,在养分限制下,微生物对土壤有机质的“氮挖掘”可以主导正激发,但在微生物可用养分足够的情况下,无论是通过外源输入还是土壤本底有机质的矿化,“微生物化学计量分解”都可以随着时间的推移成为主导机制[25]。Song 等[84]的研究表明,外源葡萄糖添加后,所引起的正激发效应是因为“微生物共代谢”。在培养早期阶段,微生物分解复杂的有机物时分泌更多的氧化酶和氮水解酶以获得自身所需的氮,微生物进行“氮挖掘”产生正的激发效应,随着培养的进行,酶化学计量比的增加降低了微生物碳利用效率并增加了激发效应强度,此时的正激发由“微生物化学计量分解”所诱导。
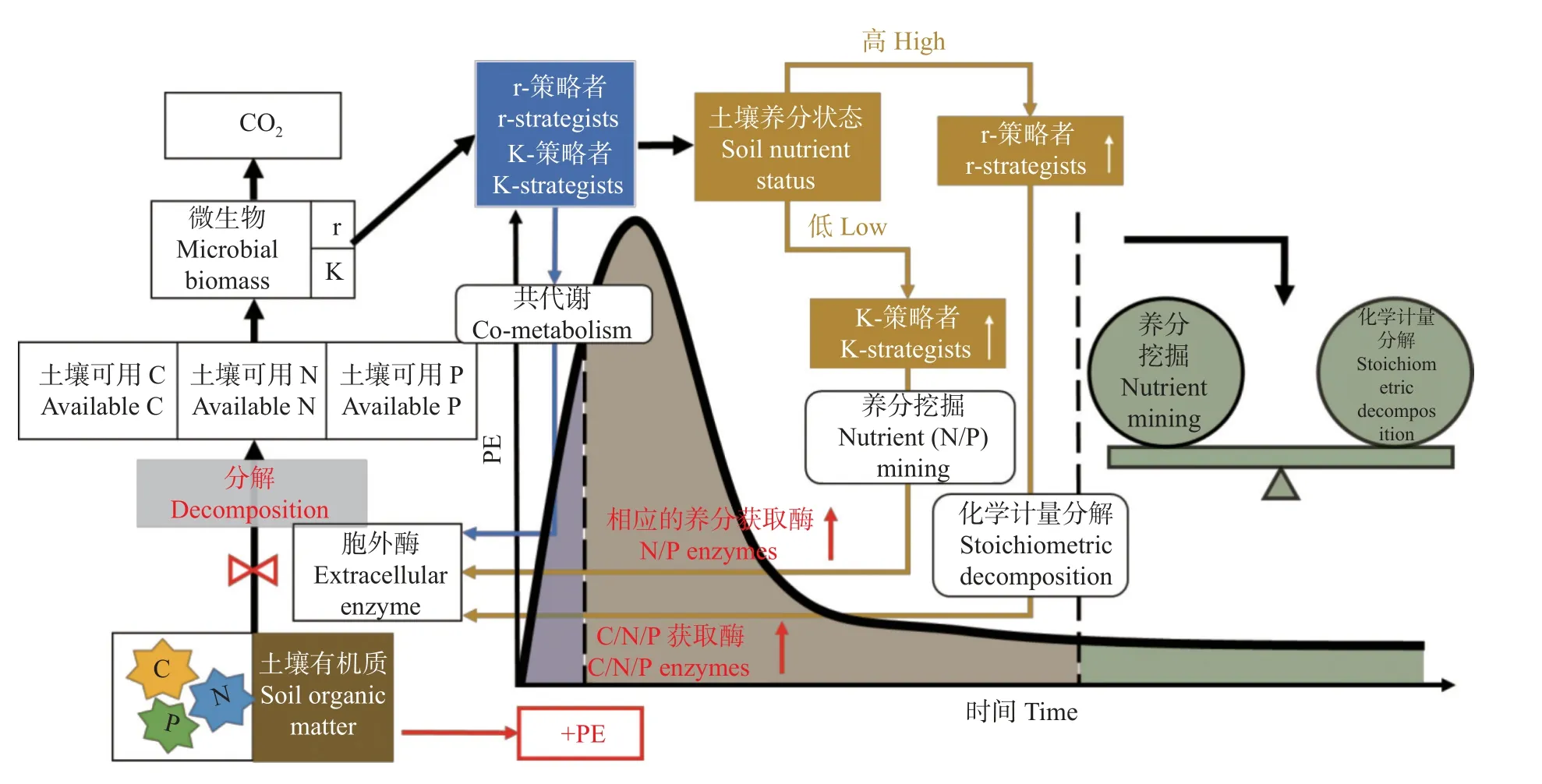
图1 微生物对土壤有机碳矿化激发效应(PE)的调控机制Fig.1 Mechanisms of microbial regulation of priming effects (PE) on mineralization of soil organic carbon
4 总结与研究展望
1)目前关于外源养分输入对土壤碳矿化激发效应的研究主要集中在单一养分的添加,对于碳氮磷协同配施方面的研究较少,未来急需着眼于农田生态系统中秸秆配施氮磷肥对于土壤碳矿化激发效应的影响,通过调节配施的比例,降低秸秆还田过程中通过激发效应损失的有机碳,提高秸秆还田增加土壤有机碳的效率。
2)微生物增殖过程本质上也是对外源碳氮磷等养分协同同化代谢的过程,通过调节土壤微生物可利用的碳氮磷化学计量比,在理论上实现对土壤微生物生物量、群落组成与微生物周转速率的定向调控,充分发挥“微生物碳泵”在提升土壤有机质方面的作用。如何优化碳氮磷输入比例、实现微生物碳泵储碳与负激发保碳的有机统一是未来需加强的一个研究方向。
3)激发效应产生的根本原因是外源碳氮磷输入后改变了土壤微生物可获得性养分的碳氮磷化学计量平衡,导致有些营养元素的亏缺或盈余,微生物为了恢复其合成代谢过程的碳氮磷化学计量平衡,会增强或减弱对土壤原有有机质的降解,进而产生正或负的激发效应。因此,未来急需将碳氮磷化学计量比调控土壤激发效应的理论运用于农田固碳减排的生产实践,减弱正激发效应,甚至将正激发效应转变为负激发效应,提高土壤固碳和碳存储能力,助力我国“碳达峰碳中和”战略的实施。
