日军细菌战与健康人力资本∗
2023-08-17周柏旭赵劲松余津嫺
周柏旭 赵劲松 余津嫺
一、引言
在战争中使用生物武器对经济发展的影响有多大? 一些研究认为战争对人口和经济发展转变造成了深远的影响,尤其是与战争相关的瘟疫大流行(Alfani,2013;Besouw 和van Curtis,2021;Voigtländer 和Voth,2013)。例如,Alfani (2013)发现17 世纪战争引发的鼠疫流行导致意大利人口锐减和政府财政能力减弱,并最终使经济发展停滞。也有研究认为,1350—1700 年间欧洲持续的战争虽然导致了瘟疫大流行和人口锐减,但也使得城市化加剧和工资率上升,最终推动了“欧洲的崛起”(Voigtländer 和Voth,2013)。普遍的观点认为战争总是伴随着大量平民的伤亡,然而一些特殊战争的影响却较少得到关注。依据史料记载(军事科学院,2015),日本侵华战争造成中国军民伤亡3 500 万人以上,其中平民占绝大多数。日军实施的细菌战也使得大量平民染疫而亡,但由于细菌战会向周围扩散,具体的人口损失较难估计(刘庭华,2015;袁成毅,2010)。因此,关于细菌战这类生物武器使用对经济发展的影响亟待研究。
本文研究了日军细菌战对健康人力资本的影响。已有文献主要通过历史上的战争冲击来识别战争对健康人力资本的影响,研究的问题主要集中在不同国家和地区的战争经历对儿童健康状况的长期影响上(Akresh 等,2012;Minoiu 和Shemyakina,2014;Singhal,2019),结论一般认为早期经历过战争的儿童会在成年后有更低的身高、受教育年限和收入水平(Guo,2020;Saing 和Kazianga,2020)。与本文较为相关的研究来自Li 和Koulovatianos (2020),他们发现中国1937—1950 年的战争对个体的肺功能有长期负面影响。还有一些文献讨论了一些特殊的战争对健康人力资本水平的负面冲击。例如,Le 等(2022)发现越南战争(1955—1975 年)中,美军使用的有毒“橙剂” 显著增加了当地平民的血液类患病率和残疾率;Arcand 等(2015)研究了安哥拉的地雷对儿童身高和体重的负面影响;Saing 和Kazianga (2020)检验了美军在1969—1973 年间向柬埔寨投放的炸弹对健康和教育人力资本的长期负面影响。相比之下,尚缺乏对战争中生物武器使用导致的健康人力资本损失相关研究。本文将第二次世界大战中日军在中国战场上进行的细菌战作为准自然实验,回答以下研究问题: 细菌战通过何种渠道对地区人力资本水平造成负面影响? 这种影响范围有多大? 是否是战时大规模平民伤亡的主要因素之一?
在侵华战争期间,日军曾在中国多地实行细菌战。不同于常规武器,日军的细菌炸弹作为一种生化武器不仅会造成被攻击者伤亡,还可通过人—人传染,或人—物—人传染造成大面积感染。根据联合国数据,1 克天花病毒制成的生物武器可致10 万至100 万人死亡。①互联网档案馆,https://web.archive.org/web/20220710063429/;联合国网站,http://www.un.org/chinese/peace/disarmament/biological.htm。各类细菌的特点不同,造成的损失也有所不同。臭名昭著的日军731 部队就曾在中国东北地区的平房建有专门研究各种致命性细菌的实验室,并在多地进行人体与战场实验(陈致远,2014)。王希亮(2012)提到,731 部队研究成员金子顺一的论文表明,一公斤鼠疫跳蚤投放在第二次感染时的人数最多可达24.36 万人,说明细菌战具有扩散性。尽管目前关于日军细菌战对健康损失的案例研究较多,但在全国范围内的因果研究仍然缺乏,这为细菌战的健康损失调查带来了挑战。
结合日军细菌战种类、地点以及战区内外瘟疫流行的历史数据,本文检验了日军在侵华战争中实施的细菌战是否导致了疫情的扩散,并在此基础上讨论了细菌战核心区域总体健康水平会受到何种影响。本文发现: 第一,霍乱、伤寒和鼠疫细菌战均导致了周边相对应瘟疫的流行,分别可扩散至周边250 千米、50 千米和25 千米范围内;第二,鼠疫和伤寒细菌战的扩散效应显著发生在战区,而霍乱细菌战在战区和非战区都可扩散至周边250 千米内;第三,离三类细菌战中心每近100 千米,生存率相较于战前会显著下降大约8 个百分点。
我们的研究对以下三个方面的文献有所贡献: 第一,丰富了战争对人力资本影响的相关研究。本文在全国范围内检验了细菌战对健康人力资本的影响,弥补了关于生物武器对健康影响实证研究的不足。而且,Becker (2007)曾强调像流行病这类影响人类生存率的因素对健康人力资本有重要影响②Becker (2007)通过1918 年大流感的例子说明,若流行病的传染率较小,疫苗研发成本将大于收益,是否值得研发还需慎重考虑。,我们关于细菌战对疫情扩散范围和生存率的研究可为相关流行病的疫苗研发和公共卫生政策制定提供参考。第二,我们的发现还可与近年来一支强调战争和瘟疫对经济发展影响的文献进行对话(Besouw 和Curtis,2021;Glaeser 和Shapiro,2002;Voigtländer 和Voth,2013)。Voigtländer 和Voth (2013)认为古代欧洲战争和黑死病流行冲击重塑了不同地区的人口比例,城市提升了对劳动力的需求,工资率上升,农村人口向城市转移,最终促进了城市化发展。而我们发现不同的细菌战所造成的疫情范围有所不同,战区与非战区的影响范围也有差异,不同的瘟疫冲击对经济发展的影响可能会有差别。我们还发现细菌战对农村和城市地区的生存率影响并没有太显著的差异,那么城市“安全港” 效应仍值得进一步讨论(Glaeser 和Shapiro,2002),这与Besouw 和Curtis (2021)的发现是一致的。因此,战争和瘟疫的影响范围对经济发展起着决定性的作用。第三,本文通过对日军各类细菌战与同时期对应的疫情关系进行因果推断,回应了已有文献在这一问题上的争议。许多研究都认为日军细菌战在中国战场中致死的人数在数百万以上,但对两者间的关系并没有很详细的说明(刘庭华,2015;谢忠厚,2016;袁成毅,2010)。这些估计数字大多来自不完全的档案记载,而且当时中国的防疫机构并不健全,难以证明这些病菌传染来自细菌战。一些区域在抗战前就爆发过瘟疫,瘟疫流行与战争之间是否存在因果关系也存在争议(曹树基,2012;周东华,2020)。本文的研究丰富了学界对此问题的相关讨论。
二、历史背景和数据
(一)日军细菌战
1931 年“九•一八” 事件后,日军开始建立细菌战研发和实验基地。在占领中国东北地区大量领土后,日军细菌战创始者石井四郎于1932 年在今黑龙江省五常县设立了最初的细菌实验场,紧接着于1933 年在哈尔滨南岗区设立了一个新的细菌研究所(陈致远,2014)。1936 年,为侵犯苏联作准备,石井四郎又在哈尔滨平房建立了一个大规模的生化武器生产根据地,即臭名昭著的“731” 部队(张树军和李忠杰,2016)。
1937 年全面抗战爆发后,日军陆续在中国战场上投放大量生物武器。除了731 部队,日军在各占领区还成立了细菌战支部。1933 年,日军在今沈阳市的“关东军临时病马收容所” 开始研究针对牲畜及农作物的细菌武器,亦称为第100 部队,以探求通过其他生物来传染人的方法。其生产的细菌包括鼻疽、炭疽、牛瘟、羊痘、锈菌和斑驳菌(陈致远,2014)。在一次实验中,这类细菌武器通过飞机投放撒播,受试的300 头牛羊全部死亡(张树军和李忠杰,2016)。至1945 年,第100 部队的细菌生产能力已经可以大面积感染牲畜及农作物。其他细菌战部队还包括在北京设立的华北“甲” 1855 部队,南京的华中“荣” 1644 部队,和广州的华南“波” 8604 部队。已经有大量研究显示,这些部队在731 部队的协助下对各地实施过细菌战,如1940 年的浙江细菌战、1941 年的常德细菌战、1942 年的浙赣和绥西细菌战以及1943 年的鲁西细菌战等(谢忠厚,2016)。随后,这些地区爆发了大规模瘟疫流行,例如谢忠厚(2016)提到福建永安城每遭到日军机轰炸一次,当地就随之流行一次瘟疫。这说明日军很可能通过飞机撒播的方式向未被占领的地区实施细菌战,这被称为“雨下法”。甚至,日军在占领新加坡后也设立有细菌战部队,被称为南洋“冈” 9420 部队。这支部队的生产能力仅次于731 部队,并参与了在滇西的细菌战,包括鼠疫和霍乱等(陈致远,2014)。根据《井本日志》的记载,在浙江细菌战实施过程中,目标及攻击法可灵活,攻击可重复,如“攻击温州由气象因素决定”。石井四郎甚至在细菌战攻击金华或绍兴、诸暨等地时开始并未与参谋们达成一致(陈致远,2018)。因此,细菌战地点选择具有一定的随机性。从空间上来看,细菌战主要集中在华北与浙江地区,散发于东北地区、沿海省份和滇缅边境,一些地区如丽水被细菌战多次攻击。内陆省份也未能幸免,如湖南常德就在1941 年遭受过鼠疫攻击(陈致远,2018)。总之,日军自全面抗战爆发开始,在中国各地实施过多次细菌战攻击,华北地区和浙江地区是遭受攻击次数最多的区域。
本文关于细菌战的数据主要来源于《日本侵华细菌战研究报告》(谢忠厚,2016)、《侵华日军在中国实施的霍乱细菌战研究》(罗运胜,2020)、《侵华日军在中国实施的鼠疫细菌战研究》(陈致远,2018)、《侵华日军细菌战纪实》(郭成周和廖应昌,1997)和《日本侵华细菌战》(陈致远,2014)。这些研究通过严谨的考据考证,从众多一手史料中摘录了各次细菌战的发生概况。我们根据上述资料中提供的文本,人工整理出各次细菌战发生地点和时间。①例如,1942 年2 月28 日《解放日报》报道: “1 月,敌‘扫荡’ 定县撤退之时,投放老鼠跳蚤甚多,经我军用显微镜检查及化验结果,断定此为带鼠疫菌的病鼠。” (转引自谢忠厚,2016)在这些研究的基础上,我们对每一次有记录的细菌战进行了交叉验证。②目前,日本方面并未提供关于细菌战的全面且系统资料,仅有1949 年伯力审判中的少量证词。
日军在细菌战中主要使用的细菌为霍乱、鼠疫和伤寒,因此本文仅考察此三种类型的细菌战是否导致了相对应的疫情扩散。③尽管相关资料也提及痢疾、疟疾、肺痨、回归热和炭疽的细菌战类型,但这类细菌战记载的次数太少,且通过多处资料比较后无法进行交叉验证,此处留待进一步研究。有史料和研究指出,日军大规模使用了三类细菌武器。例如,陈致远(2014)提出,《井本日记》中记载了日军用飞机投撒了70 公斤伤寒、50 公斤霍乱和5 公斤鼠疫跳蚤。陈致远(2018)还发现,虽然日军在实验室里研究的细菌武器以鼠疫、炭疽和鼻疽菌居多,但在战场上仍以霍乱、鼠疫和伤寒菌居多。我们搜集到有明确记载的霍乱、鼠疫和伤寒细菌战各59 次、57 次和28 次。其中,霍乱细菌战在1938—1945 年都有不同程度的实施,1943 年更是达到了最高峰22 次。鼠疫细菌战自1940 年以来均有实施,在1942 年高达25 次。伤寒细菌战自1938 年开始实施,但到1943 年及以后便没有明确的记载。
(二)抗战期间的瘟疫流行
抗战爆发后,各地的瘟疫流行逐渐增多,日占区的瘟疫尤其多。河南省《信阳县卫生志》曾记载,“日军入侵本县,人民四处逃难,……平昌关一带发生霍乱” (转引自龚胜生,2019)。又有江西省《冈上镇志》写道,“霍乱流行随日军频繁轰炸及骚扰,村民纷纷向抚州、南城、吉安、泰和、峡江、赣州一带逃难,几乎十室九空,很多难民在流亡过程中死于鼠疫、霍乱等病” (转引自龚胜生,2019)。除鼠疫和霍乱外,也有记载表明伤寒流行与日军有关,例如,内蒙古自治区《鄂温克族自治旗志》记载,当地三人因反抗日军欺辱被捕,其中一人被注射病菌后放回家,不久全家四人染伤寒病而死,后又蔓延至南辉河、伊敏河流域,死亡牧民达二百四十余人(转引自龚胜生,2019)。1938年8 月31 日的《申报》也曾报道,“日本占领下之区域发生严重霍乱” (转引自龚胜生,2019)。因此,在战争期间,各地发生的瘟疫流行与日军进行的细菌战存在重叠的情况。
本文使用的疫情基础数据来自《中国三千年疫灾史料汇编》(龚胜生,2019),该书以编年体形式,汇集了公元前674—公元1949 年各类史料中的疫情记载。在此书的基础上,我们整理出1880—1949 年的13 418 次县级疫情记载,并对各类疫情进行分类,主要包括霍乱、鼠疫、伤寒、天花、回归热、痢疾、疟疾和白喉。图1 为1880 年至1949 年主要疫情记载数的时间趋势。④关于霍乱的记载还包括其他名称,如“虎烈拉”、“虎疫”、“吊脚痧” 和“吊足痧”。鼠疫虽分为腺鼠疫和肺鼠疫,但书中除少数记载外,基本上只有“鼠疫” 的记载,并无详细的分类,因此都归类为鼠疫。伤寒与副伤寒在临床上不易区分,统一归类为伤寒。痢疾还包括“赤痢”、“痢疫”。可以看出,自1937 年全面抗战开始,各类疫情数在全国范围内呈现流行加剧趋势。1945 年全面抗战结束之后,各类疫情次数才开始逐渐减少,1949 年的记载数量与战前的1930 年大致相当。

图1 主要疫情记载数(1880—1950 年)
三、变量与识别策略
(一)核心变量
本文构建了县级的瘟疫流行和细菌战面板数据,以检验全面抗战期间(1937—1945年)瘟疫发生与日军细菌战之间是否存在因果关系,并在此基础上进一步分析细菌战与生存率之间的关联。县级行政区划数据来自《中华人民共和国县级以上行政区划沿革»(中华人民共和国民政部,1986)中1949 年的地图,共包含2 690 个县级区划单位,基本上覆盖了本文所研究的地理范围。①相较于哈佛大学中国历史地理数据库(CHGIS)在1911 年的数据,此数据的行政区划年份更接近抗战时期,因此更适合本文研究。
1.细菌战与瘟疫
本文的因变量为瘟疫是否发生的虚拟变量,即某县在相应年份中有瘟疫记载则赋值为1,否则为0。核心解释变量为某县受细菌战的影响程度。由于细菌战产生的影响会以细菌攻击地为中心向外扩散,我们使用某县与细菌战中心的最短直线距离作为衡量指标。具体衡量方式为: 首先,根据细菌战中心位置,借助ArcGIS 软件,划分出距离细菌战中心的不同区段范围;随后,生成不同区段范围内的虚拟变量,即若某县落入相应区段范围内则赋值为1,否则为0。②例如,0—25 千米的虚拟变量中,某县若落入细菌战中心25 千米范围内,则赋值为1,否则为0;在25—50 千米的虚拟变量中,若某县落入25—50 千米内,则赋值1,否则为0。由于细菌战的影响很难超过500 千米范围,因此我们也将分析样本限制在离细菌战中心500 千米的区域范围内,这一区域已包含至少1 813 个县,具有较好的代表性。③由于本文分别检验了霍乱、鼠疫和伤寒三种不同类型的细菌战,因此各自中心周围500 千米范围内所包含的县数量不同。
2.生存率
本文利用出生队列规模增长率作为生存率的代理变量。由于无法获取民国时期省级区域以下的人口数据,我们根据Meng 等(2015)和Chen 等(2020)的做法,利用人口普查微观数据抽样得到的各区域在不同年份的出生样本容量,即出生队列规模,来衡量细菌战核心区域对健康水平的影响。出生队列规模是一个衡量生存率的良好指标,因为它隐含了生育率、婴儿死亡率和后期死亡率等因素(Meng 等,2015)。
本文采用的人口普查数据来自国际微观共享整合数据库(Integrated Public Use Microdata Series,IPUMS)①参见: https://www.ipums.org。公布的地级市的1990 年中国人口普查微观数据,抽样方式为分层整群。本文利用了其中1919—1945 年的出生队列数据。出生队列规模能否反映各区域在各年的总体情况,取决于抽样调查在各区域和各出生年份中是否均衡抽样。为此,本文比较了出生队列规模与《中国1990 年人口普查资料》(国务院人口普查办公室和国家统计局人口统计司,1993)中普查总人口数据,两者在时间和空间上高度相关。②因篇幅所限,本文省略了出生队列与普查总人口相关性图,感兴趣的读者可在《经济科学》官网论文页面“附录与扩展” 栏目下载。“附录与扩展” 中图A1 (a)为出生队列规模与普查总人口在各出生年份的趋势,两者趋势几乎一致;图A1 (b)和图A1 (c)分别为两者在出生年份和省份中的相关性,说明IPUMS 提供的数据在抽样时也考虑了时间和空间上的均衡性。因此,本文利用出生队列规模增长率来衡量生存率具有一定的合理性。
此外,我们还搜集并计算了其他指标来反映健康人力资本,以及检验人口迁移可能造成的偏误。第一,我们利用出生队列规模计算了就业人口生存率、教师就业人口生存率以及小学毕业人口生存率,来反映长期中的健康人力资本质量。第二,我们人工整理了1936 年和1953 年两次县级全国人口普查中的总人口数,并计算出1936—1953 年总人口增长率。③1936 年人口数据来自《户口统计》(中华民国内政部统计处,1938),但不包括当时被日本占领的东北地区。虽然抗战胜利后的1946—1948 年间有过三次全国户口统计,但由于内战爆发,统计范围远不能覆盖全国,且许多数字为旧有材料的估计(侯杨方,2001)。第三,人口迁移也可能导致生存率的这一指标的偏误,因此我们剔除了人口流动较大的城市地区样本作为稳健性检验。④另外,曹树基(1997)曾提到,1949 年海外移民只占人口总数的2.5%。乔启明(1945)也发现1929—1933 年间的中国乡村人口迁移率仅为4.5%,且大部分在县内迁移。
3.其他控制变量
我们还考虑了一些地理及社会经济因素对瘟疫流行的影响。首先,农业自然产出会影响人类营养健康水平,进而影响对病菌的抵抗能力。我们选取Galor 和Özak (2016)的卡路里土地适宜度指数(Caloric Suitability Index,CSI)来衡量各县潜在的农业产出。⑤可从此网站获取: https://ozak.github.io/Caloric-Suitability-Index。第二,海拔和地形崎岖度可能会影响携带病菌的个体迁徙路径和速度,从而对疫情的传播范围产生影响。因此,我们计算了各县的平均海拔和地形崎岖度,数据分别来源于美国航空航天局(NASA)⑥可从此网站获取: http://www.webgis.com。与Nunn 和Puga (2012)。第三,人口密度是影响疫情传播的重要因素之一,因为人口密集地区容易出现人际交往频繁的情况,从而促进细菌的传播。我们利用中华续行委办会调查特委会(2007)于1920 年调查的1 703 个县的人口数,作为研究抗战前人口统计的重要参考。第四,我们还利用《中国抗日战争史地图集》(武月星,1995)中的数据检验了战区与非战区的异质性。主要变量统计描述如表1 所示,所有样本均限制在离各细菌战中心距离500 千米内,时间跨度为1887—1945 年。
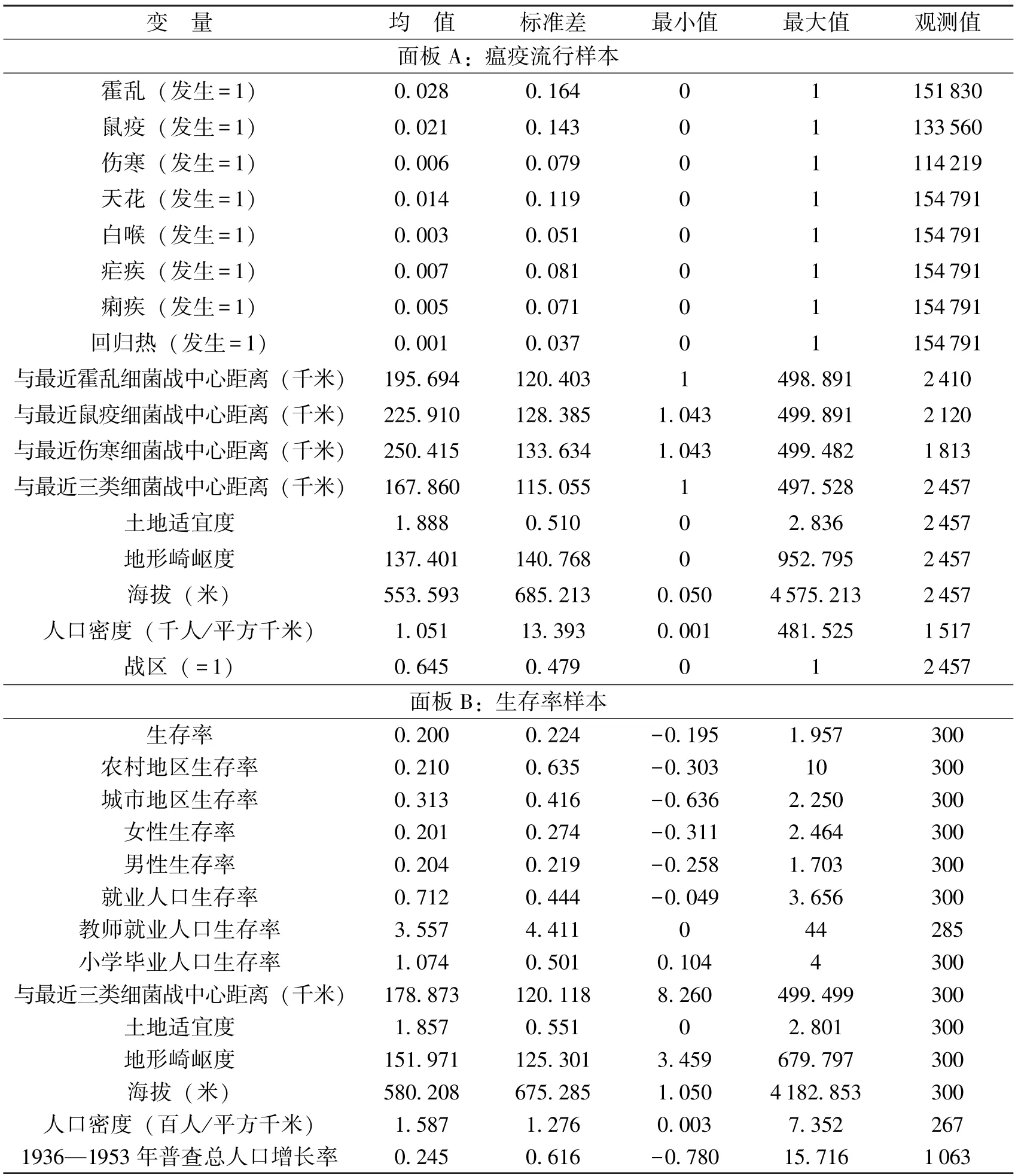
表1 主要变量描述统计
(二)识别策略
识别日军细菌战对同时期疫情扩散的因果效应面临两个方面的挑战。第一,目前明确记载的霍乱、鼠疫和伤寒细菌战只有144 次,且一些县城被多次和多种细菌攻击,相较于本文样本中2 690 个县级区划单位来说数量太少。如何选取控制组,即与实际细菌战攻击地点相似但实际没有受到攻击的区域,就成为一个需要解决的问题。第二,在受到日军细菌战攻击的地点中,有些地方可能同时处于自然疫源地,这可能会对因果关系造成一定程度的干扰和混淆。比如,福建、广东和广西等省份在抗战前一直是鼠疫流行地区,而抗战时期的鼠疫流行是自然爆发还是因日军细菌战这一问题也一直受到历史学界的关注,且已经成为较有争议的问题之一(曹树基,2012;周东华,2020)。
针对上面两个挑战,本文参照了Dell 和Olken (2020)的反事实估计方法来构建处理组和控制组,以及反事实组。第一,根据传染病会通过人—人或人—动物—人向周围扩散的特点,本文将距离细菌战中心依次渐远的县作为控制组。若能观察到离细菌战中心越近的县疫情发生的概率越高,那么说明细菌战与疫情扩散有较强的因果关联。第二,为了排除细菌战中心也可能是自然疫源地的情况,我们将全面抗战爆发前50 年(1887—1936 年)的各类疫情流行发生情况作为反事实组。具体而言,1887—1936 年县城并未受到细菌战攻击,因此可以用于建立反事实组,以帮助消除因自然疫源地而产生的干扰和混淆因素,更准确地评估细菌战对疫情传播的影响。①Dell 和Olken (2020)应用该方法估计了19 世纪中期荷兰殖民者在印度尼西亚爪哇岛建造的甘蔗工厂是否对当地经济发展产生持续影响。由于历史甘蔗工厂的样本数据较少,他们利用现代调查的截面数据构建了历史甘蔗工厂的反事实工厂,即适合殖民者建造但实际没有建造甘蔗工厂的区域,通过估计反事实工厂所在地与真实工厂所在地经济发展的差异,从而分离出真实工厂的影响效应。若细菌战中心周围各县在全面抗战期间与抗战前50 年的疫情发生情况有显著差异,仍然能够说明日军实施的细菌战与战时疫情流行扩散有因果关联。
基准回归方程如下:
其中,epidct为某县瘟疫是否流行的虚拟变量,若c县在t年发生瘟疫则取值为1,否则为0。等式右边为c县到最近细菌战中心地点的距离,用于度量c县受细菌战的影响。例如,是指若c县与最近的细菌战中心距离在0—25 千米区段范围内则为1,否则为0。以此类推,区段i的选取还有25—50 千米,50—75 千米,……,375—400 千米。基准组为400—500 千米,即样本中离细菌战中心最远的县。②细菌战中心周围各区段范围内县样本的分布请见《经济科学》官网“附录与扩展” 中图A2,可见除了离霍乱细菌战400 千米外,各区段样本数量分布较均匀。受关注的系数γi可被解释为,与较远的地方相比,离细菌战中心第i区段范围的县发生疫情的概率变化程度。在基准回归中,为了尽可能精确识别各区段的扩散效应,我们选取较小的25 千米间距区段进行分析。③我们在稳健性分析中选取了不同的反事实组(见“附录与扩展” 中图A3)、限制样本(见“附录与扩展” 中图A4)和改变区段间距(见“附录与扩展” 中图A5)等不同的模型识别,结果依然稳健。Xc是一系列控制变量,包括土地适宜度、地形崎岖度、海拔和人口密度。是各县与最近同一个细菌战中心距离的固定效应,这能够确保处于相同最近细菌战中心的各县被比较,而不是与其他细菌战中心的县比较。νt为年份固定效应,εct是误差项。
我们通过式(1)分别估计全面抗战期间(1937—1945 年)的系数与战前50 年(1887—1936 年)的系数,两者点估计的差分即为细菌战对周围县疫情发生概率的影响。由于战前50 年前的反事实组与抗战期间处于相同的位置,通过两者差分可在一定程度上消除一些随位置变化但无法观察到的影响,例如一些自然疫源地的影响。不仅如此,本文还参考Dell 和Olken(2020),计算出一个p值: 全面抗战前的系数绝对值分布大于全面抗战期间对应的的系数绝对值的比例。较小的p值表示受细菌战影响的县在抗战前不太可能有此类瘟疫流行,且战前与战后差异较大。①Dell 和Olken (2020)的方法与双重差分模型的思想一致。此方法不仅在空间上比较了细菌战影响的差异,还在时间上比较了差异。与双重差分类似,我们也报告了战前的“平行趋势”,即战前50年细菌战周围各区段里细菌战对瘟疫流行影响的系数核密度分布,如图2 左图所示。
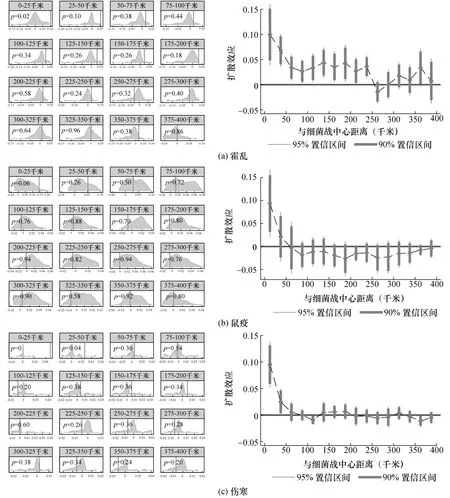
图2 基准回归: 霍乱、鼠疫和伤寒细菌战的扩散效应
四、细菌战对瘟疫流行扩散的影响
(一)基准回归结果
图2 为通过式(1)估计出的霍乱、鼠疫和伤寒细菌战对周围地区疫情发生概率的影响。首先,图2 中左分图为与细菌战中心距离各区段在全面抗战前50 年各年的估计系数分布,垂直线为全面抗战期间估计系数。若系数分布集中于0 值附近,说明细菌战中心在战前瘟疫流行的可能性不大。若p值较小,则表示全面抗战前后的瘟疫发生概率差异较大。图2 中右分图中的各区段系数则是分别通过全面抗战期间与战前50 年的样本分别计算出估计系数,再相减得到两者的差分估计值。这些估计系数表示,在消除了可能的自然疫源地影响后,与较远的地区相比,离最近细菌战中心不同区段范围内各县疫情发生的概率大小。
从图2 (a)可以得知,霍乱细菌战对周边地区霍乱疫情有显著的影响。图2 (a)左分图显示,在战争前各区段范围内估计值都集中在0 值附近,说明战前细菌战中心周围地区较少出现霍乱疫情。而且,与其他区段相比,50 千米范围内区段的p值均较低,说明在最靠近细菌战的中心区域,抗战前后差异较大。因此,我们无法拒绝霍乱细菌战造成了周围霍乱瘟疫扩散的假设。图2 (a)右分图显示,在霍乱细菌战中心250 千米内的县发生霍乱疫情的概率有显著上升。在最近的25 千米区段内的县发生霍乱疫情的概率最大,大约会上升10%。从表1 的描述统计可知,离霍乱细菌战中心500 千米范围样本内的霍乱瘟疫发生的概率均值为0.028,因此0—25 千米区段内的县受到细菌战影响而发生霍乱疫情的概率为均值的4 倍左右(=0.1/0.028),表明这些县受细菌战影响非常大。而在逐渐远离细菌战中心的250 千米区段内,霍乱疫情发生的概率下降至大约3%。在250 千米外,霍乱疫情发生的概率不再显著,说明离霍乱细菌战较远的地区不太可能发生霍乱疫情。因此,日军在中国实施的霍乱细菌战显著导致周围250 千米范围内地区的霍乱疫情扩散。
图2 (b)部分显示了鼠疫细菌战对周边地区鼠疫疫情的影响。首先,图2 (b)左分图显示,在距离细菌战中心较近的区段里,抗战前的估计系数分布并不总是在0 值附近而是稍微右偏,说明细菌战中心周边地区在战前的鼠疫疫情也可能受到自然疫源地的影响。尽管如此,在25 千米区段范围内的p值仍然很小,只有0.06,说明在最近的25千米内鼠疫疫情发生概率在战争前后差异较大。而在逐渐远离细菌战中心的区段中,抗战前后的差异不再显著。从图2 (b)右分图中也可以看出,只有25 千米区段范围内的县发生鼠疫疫情的概率较大且显著。具体来说,在离鼠疫细菌战中心最近的25 千米内,鼠疫疫情发生的概率显著上升大约10%。该影响是样本中均值的5 倍左右(=0.1/0.021),稍高于霍乱。虽然在25—50 千米及更远区段的影响不再显著,但仍然可以看出点估计值是逐渐减小的。因此,鼠疫细菌战显著影响疫情发生概率的范围在25 千米内,但在50 千米内仍表现出一定程度的扩散效应。
图2 (c)是伤寒细菌战对周边地区伤寒疫情流行扩散的影响。很明显,图2 (c)左分图中战前的各区段系数都集中在0 值附近,说明战前伤寒疫情并不经常出现。在50 千米范围内,抗战前后的差异较大。图2 (c)右分图则显示,伤寒细菌战中心50 千米范围内有显著的伤寒瘟疫扩散效应。在25 千米区段范围内,伤寒疫情也显著上升了大约10%,其影响是均值的17 倍左右,比霍乱和鼠疫细菌战在同样范围内的影响更大。25—50 千米区段的伤寒疫情发生概率显著上升大约3%,其影响是均值的5 倍左右。因此,日军实施的伤寒细菌战显著增加了周边50 千米范围内伤寒疫情发生概率。
(二)战区与非战区
日军实施的大多数细菌战位于战区,那么非战区的疫情是否与日军细菌战有关? 我们分别估计了战区和非战区样本的细菌战扩散效应。图3 报告了战区与非战区分样本的回归结果。在图3 (a)战区样本中,回归结果与基准回归中的结论基本一致。但在(b)非战区样本中,除霍乱细菌战仍然在250 千米范围内有显著的扩散效应外,鼠疫和伤寒细菌战均不再显著。战区与非战区呈现出不同的扩散效应,说明细菌战的影响范围还取决于所处的环境。因此,日军细菌战导致的疫情扩散主要集中于战区,而霍乱细菌战在非战区的传播范围依然可以达到细菌战中心周围250 千米范围内。①为了排除战争强度可能造成的影响,我们还将战争持续年数作为控制变量加入回归,发现并不影响本文结论,结果请见《经济科学》官网“附录与扩展” 中图A6 和表A1。
(三)稳健性分析
1.平均效应
我们首先考虑直接使用与日军细菌战距离作为解释变量来检验稳健性。在基准回归中,采用与细菌战中心距离各区段虚拟变量的方法可以观察到细菌战对不同区段疫情的影响。而直接使用距离的方法进行回归分析,得到的估计值应该被解释为细菌战对周围地区疫情发生率影响的平均效应。我们利用双重差分法来估计其影响,识别方程如下:
与式(1)不同的是,d2epidCenterc表示c县与最近细菌战中心距离,I(1937≤year≤1945)表示观测值在1937—1945 年间为1,否则为0。δ即为所关注的系数,表示与细菌中心距离每增加1 千米,疫情发生概率的变化。uc和vt分别是县固定效应和年份固定效应。其他变量与式(1)一致。为了排除样本选择的偏误,我们分别对距离最近细菌战中心500 千米和300 千米的样本进行估计。
图4 报告了使用与细菌战中心距离作为解释变量的平均效应。可以看出,与霍乱、鼠疫和伤寒细菌战的距离缩短,使得发生相应疫情的概率显著上升。具体来说,距离霍乱、鼠疫和伤寒细菌战中心每近100 千米,相对应的疫情发生概率就分别至少上升1.0、0.5 和0.5 个百分点。因此,关于平均效应的稳健性分析也说明距离细菌战中心越近,疫情发生的概率就越大。
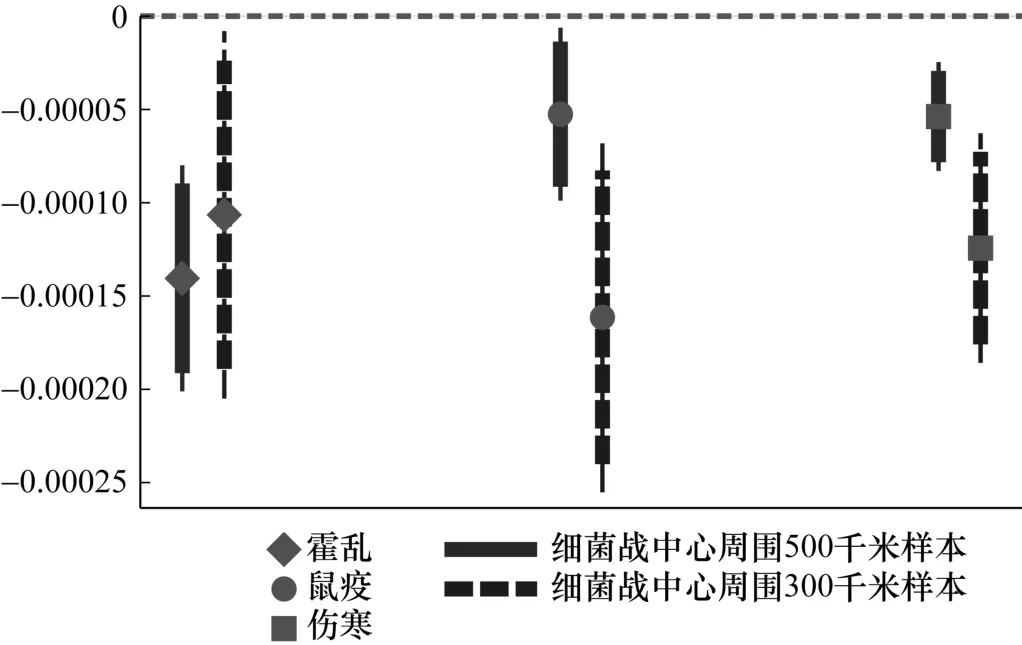
图4 平均效应
2.安慰剂检验
最后,我们检验了细菌战中心周围是否有其他种类的疫情扩散。细菌战通常只导致同类疫情的扩散,因此霍乱、鼠疫和伤寒三类细菌战对周围其他类型疫情的影响应该不显著。根据已有资料,我们检验了所有霍乱、鼠疫和伤寒细菌战中心周围是否存在天花、白喉、疟疾、痢疾和回归热疫情扩散效应。图5 的结果显示,细菌战中心周围并未发现其他类型疫情的显著扩散效应。需要注意的是,这并不能说明日军细菌战没有实施过这些细菌战,仅能说明霍乱、鼠疫和伤寒细菌战周围对其他类型的瘟疫流行无显著影响。因此,关于其他类型瘟疫的安慰剂检验表明基准回归的结果不是随机产生的。

图5 安慰剂检验
五、细菌战对生存率的影响
我们进一步分析细菌战对地区人力资本水平造成的影响。Becker (2007)曾强调,任何能够影响生存率投资的研究都应被视为关于健康人力资本的研究。生存率的变化在一定程度上可以反映人口的变化情况,因此常被用于分析各种因素与人口之间的关系(Meng 等,2015;Chen 等,2020)。我们也选用这一指标进行分析。考虑到生物武器的致命性,离细菌战中心较近的地区易出现较大的伤亡。因此,我们将与所有霍乱、鼠疫和伤寒细菌战中心的距离作为解释变量,来观察在较近的地区是否存在较大的人口伤亡。估计模型如下:
其中,cohortGrowthp是指地级市p在全面抗战期间(1937—1945 年)出生队列规模与战前(1928—1936 年)相比的增长率,以反映战争期间的生存率。d2epidCenterp是各市与最近的细菌战中心距离。受关注的系数γ表明与细菌战中心每近1 单位距离生存率的变化。Xp是一系列控制变量,包括土地适宜度、地形崎岖度、海拔和人口密度,εp是误差项。
为了让细菌战与生存率之间的关系更加直观,我们在图6 中呈现了与细菌战中心距离和生存率之间的无条件相关性。图6 显示,离细菌战中心较近的区域,生存率较低。例如,距细菌战攻击较近的衢州和丽水地区,生存率呈现负值,说明这些地区在战时的存活人数相较于战前有所下降。而离细菌战中心较远的地区,生存率较高。因此,细菌战对抗战期间的生存率有负面影响。

图6 与细菌战中心距离和生存率的无条件相关性
表2 报告了细菌战对战时生存率影响的回归结果。第(1)列全样本系数结果表明,距离细菌战中心每近100 千米,生存率会显著减少3.3 个百分点。1931 年日军迅速占领了东北三省(辽宁、吉林和黑龙江),因此该地区的出生人数可能在1937 年之前就已经开始下降。为了排除这些地区的影响,第(2)列剔除了东北地区样本,结果显示细菌战对生存率仍然有显著负面影响。第(3)列的估计值升高到8.2 个百分点,说明离细菌战中心越近影响越大。第(4)列为距离细菌战中心500 千米内样本的回归结果,显示离细菌战中心每近100 千米,生存率显著下降8.4 个百分点。第(5)列和第(6)列逐步加入了地理因素和人口密度,系数并无太大变化且仍然显著。由于细菌战主要发生于全面抗战期间,对更早的生存率应无显著影响。为此,我们在第(7)列中将因变量替换为1928—1936 年与更早的1919—1927 年相比的生存率,作为安慰剂检验。第(7)列的结果证实,细菌战对更早的生存率无显著影响。因此,细菌战显著降低了战时生存率。①我们报告了与细菌战中心距离区段效应、替换人口普查数据样本和关于人口迁移的检验结果,发现依然稳健,请见《经济科学》官网“附录与扩展”。
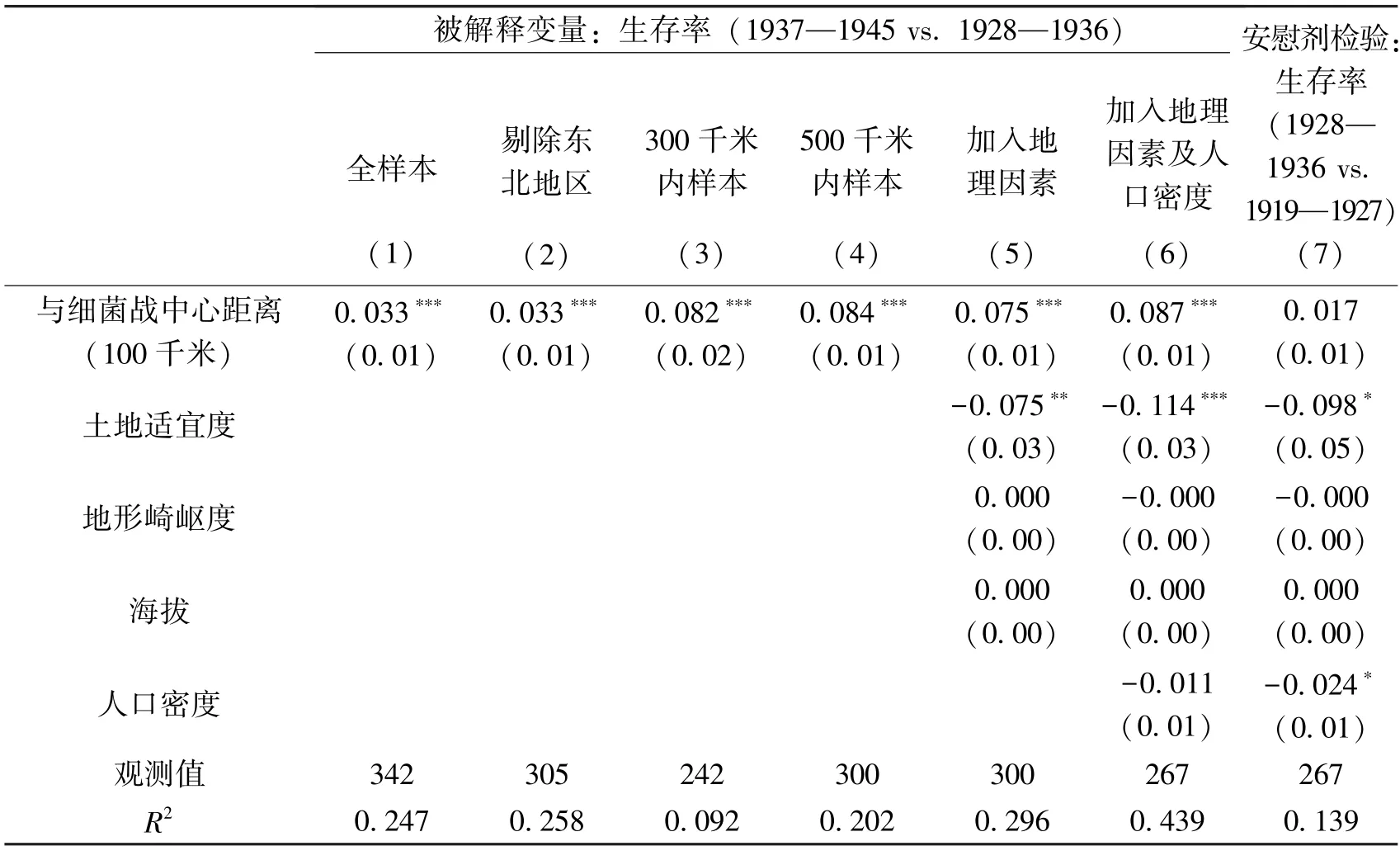
表2 细菌战对生存率的影响
表3 报告了细菌战对不同群体生存率的异质性影响。第(1)列和第(2)列分别显示,离细菌战中心每近100 千米,农村和城市地区的生存率会分别显著下降8.9 和8.6个百分点。农村和城市地区无显著差异的发现与Besouw 和Curtis (2021)一致,说明细菌战并不会造成农村和城市地区的人口结构差异。由此也说明,农村人口向城市流动会很难实现,城市“安全港” 效应仍值得进一步讨论(Glaeser 和Shapiro,2002)。第(3)列和第(4)列的结果显示,每近细菌战中心100 千米,女性和男性生存率分别会显著下降约8.8 和8.6 个百分点。因此,细菌战对农村与城市地区、女性与男性群体的生存率均有显著的负面影响,但差异不大。
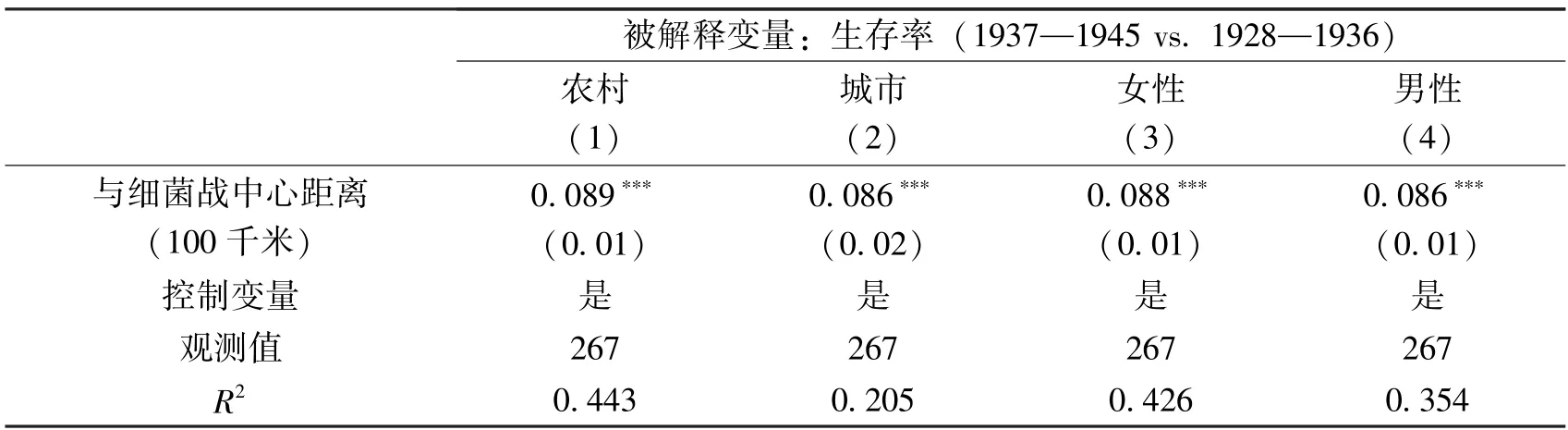
表3 细菌战对不同群体生存率的影响
最后,表4 报告了细菌战在中长期对人力资本损失的影响。我们首先通过出生队列规模计算出能够反映人力资本质量损失的生存率,然后再通过1936 年和1953 年两次人口普查数据计算出县级总人口增长率。第(1)列说明离细菌战中心越近,在长期中的就业人口生存率越低。由于教师的知识密集度较高,第(2)列报告了细菌战对这部分人群的影响,结果说明对教师就业人口生存率也存在显著的负面影响。第(3)列说明拥有小学学历的人口生存率同样受到细菌战的负面影响。最后,第(4)列表明,离细菌战中心越近,1936—1953 年的中期总人口增长率越低。因此,在中长期,细菌战对就业、教育以及总人口增长都有显著负面影响。

表4 细菌战对健康人力资本的中长期影响
六、结论
第二次世界大战中,日军在中国战场上进行了惨无人道的细菌战攻击。根据现有披露的日军实施的大规模细菌战资料,本文量化分析了细菌战与战时瘟疫流行之间的关系。结果表明,细菌战攻击会显著增加周围地区霍乱、伤寒和鼠疫疫情的发生概率,分别可扩散至距细菌战中心250 千米、50 千米和25 千米范围内。尽管其传播范围有所不同,但三类细菌战对周围地区疫情有较明显的扩散效应。本文还发现,离细菌战中心每近100千米,生存率平均下降约8 个百分点,说明细菌战对全面抗战期间的总体健康人力资本水平造成了负面影响。
本文结论有如下启示: 第一,我们关于细菌战对健康人力资本影响的量化分析说明,生物武器所造成的人力资本损失不容忽视,并可为相关流行病的疫苗研发和公共卫生政策制定提供参考。第二,我们关于细菌战所造成疫情范围不同的发现说明,战争的特殊性会影响经济发展的方向。第三,我们回应了已有文献关于细菌战是否导致同时期疫情的争议,尽管战前存在不同程度的瘟疫流行,但细菌战仍是战时大量人口伤亡的重要原因之一。
