基于SSR分子标记的面包果资源遗传多样性分析
2023-08-14伍宝朵高效梅胡丽松谭乐和闫林吴刚
伍宝朵 高效梅 胡丽松 谭乐和 闫林 吴刚

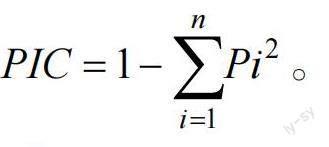


摘 要:面包果被认为是最有潜力解决热带饥荒的粮食作物,具有较高的经济价值。我国面包果资源引种于多个国家,来源较丰富,目前未对其进行系统的分类和遗传多样性分析。为明确面包果资源间遗传距离,揭示其亲缘关系,本研究从75 对SSR 标记中筛选出33 对扩增条带清晰、多态性高、稳定性好的标记,用于面包果资源的遗传多样性分析。
对面包果资源进行PCR 扩增,使用8%变性聚丙烯酰胺凝胶检测PCR 扩增产物,利用NTSYS 软件进行遗传多样性相关分析。结果表明:33 个SSR 标记的扩增条带多态性比率为94.57%,共检测出199 个多态性位点,平均每个标记检测出6.030 个;SSR 标记的PIC 值在0.623~0.940 之间,平均为0.865;面包果资源间遗传相似系数在0.517~0.951 之间,平均遗传相似系数为0.685,且硬面包果Ⅰ型ZZP 资源内个体间差异最大,平均遗传相似系数为0.626,无核型面包果XYS1-3 资源内个体间差异最小,平均遗传相似系数为0.723;遗传相似系数为0.713 时,将30 份面包果资源聚成5 类,XBL(Artocarpus odoratissimus)和园林栽培硬面包果Ⅰ型种质(ZZP、LX、STH、HJ5)聚为一类,无核型面包果(SLLK、XYS2、XYS3、XYS5、XYS6、YN)聚为一类,硬面包果Ⅱ型(SLM2、YDNXY1、G1、G2)聚为一类,硬面包果Ⅱ型(W1、W3、SMY、8-4、8-3、5-5、5-6)聚为一类;三维图能较好地将XBL 和园林栽培硬面包果Ⅰ型与食用价值较大的无核型面包果和硬面包果Ⅱ型区分开。依据聚类分组情况,可推测XYS2、XYS3、XYS5 和XYS6 可能引种于越南,XYS1-3、YBG 和XYS1 可能引种于印度尼西亚,FJ 可能是无核型面包果。该研究阐明了30 份面包果资源间的亲缘关系,为面包果品种选育、鉴别提供理论依据。
关键词:面包果;种质资源;SSR 分子标记;亲缘关系;遗传多样性
中图分类号:S667.9 文献标识码:A
面包果属于桑科(Moraceae ) 菠萝蜜属(Artocarpus),起源于南太平洋岛国,被认为是最有潜力解决热带地区饥荒的粮食作物,具有较高的经济价值[1-2]。我国海南、广东、云南、广西、福建等地均有零星种植。果实营养价值丰富,富含淀粉、蛋白质、维生素和矿物质等,树姿美,可作行道树、庭院树等[3-5]。JONES 等[6]通过形态鉴定将其分为A. camansi Blanco、A. altilis (Parkinson)Fosberg 和A. mariannensis Trécul,早期A. altilis×A.mariannensis 杂交种和驯化A. altilis×A. mariannensis雜交种。中国热带农业科学院香料饮料研究所自20 世纪90 年代开始面包果试种和种质资源保存相关研究,目前已收集保存面包果种质资源50 余份,其中无核型面包果30 余份。根据有无种子将面包果分为无核型面包果和有核型面包果,有核型面包果通称硬面包果,硬面包果包括硬面包果Ⅰ型和硬面包果Ⅱ型[7]。
简单重复序列(simple sequence repeats,SSR)具有共显性、多态性高、重复性好、种属间有良好的通用性、技术难度低等特点。已广泛应用于基因定位、遗传图谱构建、指纹图分析、品种鉴定和遗传多样性评价等[8-11]。国外学者有一些关于面包果分子标记开发和遗传多样性相关研究报道。ZEREGA 等[12]利用扩增片段长度多态性(amplified fragment length polymorphism, AFLP)对200 多个面包果品种之间的关系进行研究,绘制了DNA 指纹图谱,发现太平洋岛屿的面包果资源遗传多样性最丰富。WITHERUP 等[13]从面包果基因组文库中分离出25 个微卫星位点,并在A. altilis、A. camansi、A. mariannensis 和A. altilis×A. mariannensis 共354 份样本中检测出等位基因的平均数为2~9。ZEREGA 等[14]利用19 个微卫星位点对225 份面包果种质资源进行了评估,等位位点数为2~24 个,平均11.63 个。GARDNER 等[15]从面包果A. camansi 和A. altilis 的转录组中开发出15 个叶绿体SSR 标记,在A. odoratissimus、A. altilis、A. camansi 和A. altilis×A. mariannensis中均能扩增,所有标记在A. camansi 中均具有多态性。BELLIS 等[16]从面包果基因组DNA 文库中共获得47 607 个简单序列重复位点,并开发了50个SSR 标记,所有的位点在A. altilis 均具有多态性,每个标记的等位位点数为2~19。
遗传多样性是生物多样性的重要组成部分,可以揭示物种或居群的进化历史,对种质资源的收集、保存、评价和利用具有十分重要的意义[17-19]。
目前,暂未见我国面包果资源遗传多样性的研究报道。本研究基于前人已开发的SSR 分子标记对我国收集的不同生态区域面包果种质资源开展系统的遗传多样性分析,明确资源间遗传相似系数,揭示其亲缘关系,为品种选育和利用奠定良好基础。
1 材料与方法
1.1 材料
供试材料为保存于木本粮食种质资源分库的30 份面包果资源,采集健康嫩叶,用密封袋密封,带回实验室置于–80 ℃冰箱保存备用。
1.2 方法
1.2.1 DNA 提取 按照OMEGA DNA 试剂盒(HP Plant DNA Kit)说明书提取基因组DNA,并用微量分光光度计(NanoDrop 2000c)检测其浓度。30 份面包果种质资源信息见表1。
1.2.2 引物的合成与筛选 查阅面包果引物开发、遗传多样性研究相关英文文献,参考BELLIS等[16]开发的50 对SSR 引物(mAaCIR ) 和WITHERUP 等[13]开发的25 对SSR 引物(MAA),共75 对。引物由上海立菲生物技术有限公司合成。
1.2.3 PCR 扩增及聚丙烯酰胺凝胶电泳 PCR 扩增采用20 μL 体系:DNA 模板2 μL,10×Buffer2 μL,正反向引物各0.4 μL,dNTPs 0.4 μL,Taq酶0.3 μL,ddH2O 补足。扩增程序为:94 ℃ 5 min;94 ℃ 30 s,50~62 ℃ 45 s,72 ℃ 45 s,35 个循环;72 ℃ 10 min;12 ℃ 30 min。PCR 扩增产物电泳检测方法参照吴刚等[20]的方法,用8%变性聚丙烯酰胺凝胶检测,在150 U、2500 mA、500 W条件下电泳40~60 min。电泳结束后立即进行银染显影,将胶板放入硝酸银染液(1 g 硝酸银+2 mL甲醛+1 L ddH2O)中银染25 min,胶板银染后再放入氢氧化钠显影液(15 g 氢氧化钠+5 mL 甲醛+1L ddH2O)中显影至出现清晰条带。
1.3 数据处理
根据扩增条带的位置,有条带的读为“1”,无条带的读为“0”。构建(0,1)矩阵表。采用NTSYSpc2.1 软件对30 份面包果资源进行聚类、主成分、遗传相似系数分析,以非加权组平均法(unweightedpair-group method with arithmeticmeans, UPGMA)进行聚类分析,绘制系统聚类图,并绘制三维散点图。同时计算多态信息含量(polymorphic informationcontent, PIC)值,PIC 值等于1 减去所有等位基因频率的平方的总和,即
Pi为等位基因频率,n 为等位基因数。
2 结果与分析
2.1 SSR 标记筛选
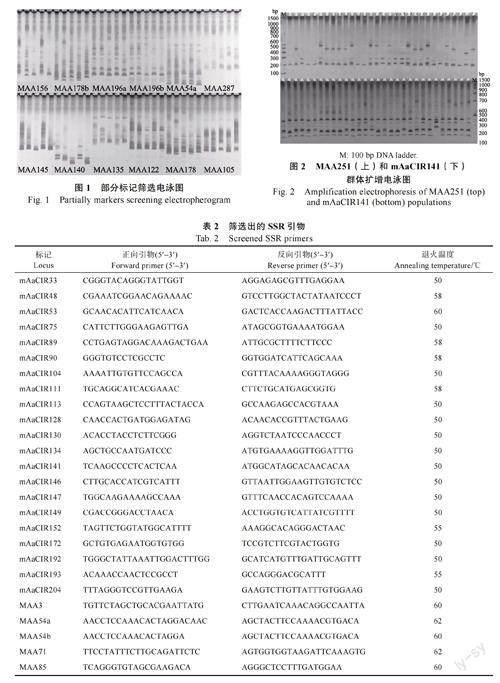
选取5 份代表性面包果资源(XYS1、XYS6、KML、LX、STH)用于引物筛选(部分引物筛选见图1)。从75 对引物中筛选出33 对(表2)扩增条带清晰、具有多态性的引物用于供试面包果资源群体扩增。图2 为选取的代表性SSR 引物MAA251 和mAaCIR141 的扩增结果。
2.2 SSR 标记多态性分析
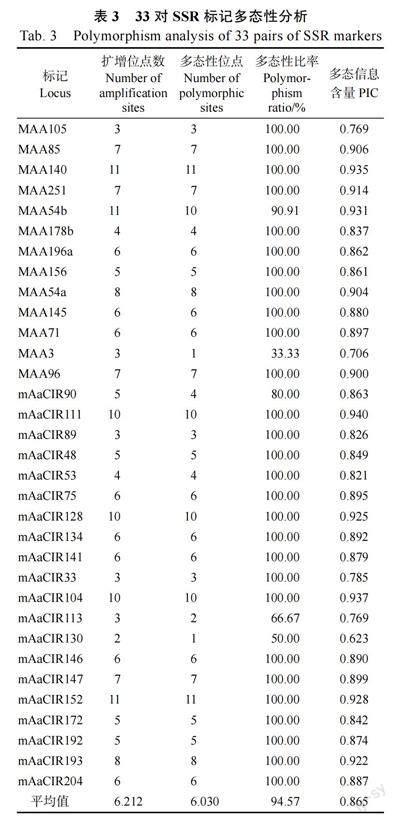
筛选出的33 对SSR 引物在30 份面包果资源中共检测出199 个多态性位点,平均为6.030 个。
其中引物MAA140 和mAaCIR152 的多态性位点最多,为11 个;引物mAaCIR130 的多态性位点最少,为1 个。除MAA54b、MAA3、mAaCIR90、mAaCIR113、mAaCIR130 外,其他引物扩增条带的多态性比率均为100%。SSR 引物的PIC 值在0.623~0.940 之间,平均为0.865。PIC 值在0.9 及以上的引物有11 对,占比为33.33%(表3)。SSR标记MAA54b、MAA178b、mAaCIR75、mAaCIR134、mAaCIR147、mAaCIR152、mAaCIR89 和mAaCIR90 在不同类型面包果资源中具有特异性扩增条带,扩增效果较好,具有较高的多态性。
2.3 遗传相似系数分析
由表4 可知,30 份面包果资源的遗传相似系数在0.517~0.951 之间,平均遗传相似系数为0.685。根据遗传相似系数可以看出不同种质资源之间亲缘关系的远近,本研究所有供试种质资源之间均存在不同程度的遗传差异。STH 与YN的遗传相似系数最小(0.517),表明遗传距离最远,遗传差异最大;XYS3 与XYS2 的遗传相似系数最大(0.951),表明遗传距离最近,遗传差异最小。同一种质资源内个体间也存在不同程度的遗传差异,其中ZZP 个体间差异最大,个体间平均遗传相似系数为0.626,最小遗传相似系数为0.532, 最大遗传相似系数为0.873; 而XYS1-3 个体间差异最小,平均遗传相似系数为0.723,最小遗传系数为0.585,最大遗传相似系数为0.922。
2.4 聚类分析
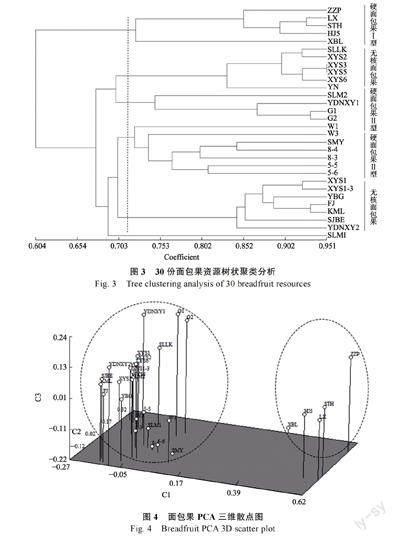
基于33 对多态性标记,用NTSYS 软件对30份面包果资源进行聚类,同时进行主成分分析(principal component analysis, PCA),绘制三维散点图。遗传相似系数聚类分析显示(图3),在遗传相似系数为0.713 时,硬面包果Ⅰ型(ZZP、LX、STH、HJ5)和XBL 聚为一类;无核型面包果SLLK、XYS2、XYS3、XYS5、XYS6 和YN聚为一类;硬面包果Ⅱ型(SLM2、YDNXY1、G1、G2)聚为一类;来源于南太平洋岛国的硬面包果Ⅱ型(W1、W3、SMY、8-4、8-3、5-5、5-6)聚为一类;无核型面包果(XYS1、XYS1-3、YBG、KML、SJBE、YDNXY2)和FJ 为1 个小组。面包果PCA 三维散点图(图4)将面包果资源分为5 个组,与聚类图一致。三维图能较好地将园林栽培硬面包果Ⅰ型(ZZP、LX、STH、HJ5)和XBL 与食用价值较大的无核型面包果和硬面包果Ⅱ型区分开。
3 讨论
面包果起源于南太平洋岛国,它的传播历程与南太平洋地区人类的迁徙历史密切相关。种质资源遗传多样性研究是保护、研究和创新利用面包果的重要步骤。WITHERUP 等[13]利用从面包果基因组文库中开发的25 对MAA 引物评估了354份样本, 不同种中检测到平均等位位点数为1.90~7.86,平均观察杂合度(Ho)和期望杂合度(He)分别为0.15~0.55 和0.20~0.62。ZEREGA等[14]利用19 对MAA 引物评估225 份面包果种质资源,平均等位位点数11.632 个,PIC 值为0.627,He 为0.441。GARDNER 等[15]从面包果叶绿体中开发出15 个SSR 标记,不同种的平均等位位点数为2.200~2.533,无偏单倍体多样性为0.147~0.602。BELLIS 等[16]从面包果基因组DNA 文库中开发了50 个SSR 标记,PIC 值为0.680。本研究采用33 对SSR 引物对30 份面包果资源进行扩增,共检测出205 个等位位点,平均为6.212 个,PIC 变化范围为0.623~0.940,平均为0.680,说明选用的33 对引物可较好地鉴定面包果种质资源的遗传差异。遗传多样性可以揭示物种或居群的进化历史,是物种适应新环境的基础,遗传多样性水平越高,物种对新环境的适应能力就越强[19, 21]。已有研究发现,菠萝蜜属包含70 多个种,起源于南太平洋岛国,香菠萝(A. odoratisimus)与A. altilis,A. camansi 和A. mariannensis 属于同一个亚属,具有较近的亲缘关系[22],本研究中XBL 和硬面包果Ⅰ型(ZZP、LX、STH、HJ5)聚为一类,说明与硬面包果Ⅱ型和无核型面包相比,A. odoratissimus与硬面包Ⅰ型具有较近的亲缘关系。ZEREGA等[12]利用AFLP 标记绘制了256 份面包果资源的指纹图谱,认为面包果至少有2 个起源中心,大多数美拉尼面包果类型和原产地西亚和波利尼西亚面包果品种是由A. camansi 经过多代无性繁殖和选育而来,而大多数密克罗尼西亚面包果品种由来源于A. camansi 的面包果和A. mariannensis杂交而来。面包果整个驯化过程中代表性的资源分别是:2 个野生祖先种A. camansi 和A. mariannensis,有核二倍体A. altilis,无核三倍體A.altilis,早期A. altilis×A. mariannensis 杂交种,高度驯化A. altilis×A. mariannensis 杂交种[23-25]。A.camansi、A. mariannensis、A. altilis(2n)和早期A.altilis×A.mariannensis 杂交种属于有核型面包果,而A. altilis(3n)和驯化A.altilis×A. mariannensis杂交种(3n)是无核型面包果[6],因此,面包果资源的聚类与地理分布和引种来源地有关。本研究中面包果样本引种于多个国家,样本来源较丰富,能代表我国面包果现有的种植情况和群体结构,聚类结果可将大多数资源根据区分开。其中XYS1-3、YBG、XYS1、XYS2、XYS3、XYS5 和XYS6 等7 份无核型面包果是20 世纪50年代由越南、泰国、马来西亚和印度尼西亚等东南亚归国华侨携带引进。聚类结果显示XYS2、XYS3、XYS5 和XYS6 与SLLK 和YN 聚为一类,因此推测这4 份资源可能引种于越南;XYS1-3、YBG 和XYS1 与KML、SJBE 和YDNXY2 聚为一类,结合我国面包果引种历史,推测XYS1-3、YBG 和XYS1 可能引种于印度尼西亚。FJ 与XYS1-3、YBG、XYS1、KML、SJBE 和YDNXY2等无核型面包果聚为一类,且与KML 的遗传相似系数为0.932,结合本研究聚类结果与类型有关,因此,推测FJ 可能是无核型面包果。
JONES 等[6]使用57 个形态描述符描述了221份面包果资源,通过数量性状描述可以将资源分为A. camansi、A. mariannensis、A. altilis(2n),A. altilis(3n),A. altilis×A. mariannensis(2n)和A. altilis×A. mariannensis(3n)6 类,也可以将资源分为来自美拉尼西亚、西波利尼西亚、东波利尼西亚和密克罗尼西亚4 个地方,置信水平均在0.95。ZEREGA 等[12]利用SSR 标记也可将资源依据类型和区域分类。我国目前还没有面包果资源形态性状描述的系统研究,因此,为更系统全面地对我国面包果资源分类及亲缘关系鉴定,后续需要多种表型性状结合分子标记进一步探讨。
参考文献
[1] 吴刚, 白亭玉, 苏兰茜, 胡丽松, 谭乐和. 面包果芽接繁殖技术[J]. 林业科技通讯, 2020(11): 74-76.
[2] 吴刚, 胡丽松, 朱科学, 秦晓威, 郝朝运, 谭乐和. 面包果在海南兴隆的引种调查初报[J]. 中国南方果树, 2017,46(4): 99-101.
[3] 符紅梅, 谭乐和. 面包果的应用价值及开发利用前景[J].中国南方果树, 2008(4): 43-44.
[4] 吴刚, 朱科学, 王颖倩, 白亭玉, 张彦军, 胡丽松, 郝朝运,谭乐和. 面包果主要营养组分研究初报[J]. 中国热带农业,2018(2): 39-44.
[5] 谭乐和. 菠萝蜜 面包果 尖蜜拉栽培与加工[M]. 北京:中国农业出版社, 2017: 121-122.
[6] JONES A M P, MURCH S J, WISEMAN J, RAGONE D.Morphological diversity in breadfruit (Artocarpus,Moraceae): insights into domestication, conservation, and cultivar identification[J]. Genetic Resources and Crop Evolution,2013, 60(1): 175-192.
[7] 吴刚, 谭乐和. 面包果品种资源与栽培利用[M]. 北京: 中国农业出版社, 2021: 17-34.
[8] 井敏敏, 黄炳钰, 戴小红, 李栋梁, 陈晶晶. 基于SSR 标记的澳洲坚果种质资源遗传多样性分析[J]. 热带作物学报, 2022, 43(2): 262-270.
[9] 周强. 老芒麦EST-SSR 分子标记开发与遗传多样性分析[D]. 兰州: 兰州大学, 2016.
[10] 陈明丽, 王兰芬, 武晶, 张晓艳, 杨广东, 王述民. 普通菜豆基因组SSR 标记开发及在豇豆和小豆中的通用性分析[J]. 作物学报, 2014, 40(5): 924-933.
[11] 仇静静. 野生花生全基因组SSR 标记的开发与应用[D].济南: 山东师范大学, 2018.
[12] ZEREGA N J C, RAGONE D, MOTLEY T J. Complex origins of breadfruit (Artocarpus altilis, Moraceae): implications for human migrations in Oceania[J]. American Journal of Botany, 2004, 91(5): 760-766.
[13] WITHERUP C, RAGONE D, WIESNER-HANKS T, IRISH B, SCHEFFLER B, SIMPSON S, ZEE F, ZUBERI M I,ZEREGA N J C. Development of microsatellite loci in Artocarpus altilis (Moraceae) and cross-amplification in congeneric species[J].Applications in Plant Sciences, 2013, 1(7):1200423.
[14] ZEREGA N, WIESNER-HANKS T, RAGONE D, IRISH B,SCHEFFIER B, SIMPSON S, ZEE F. Diversity in the breadfruit complex (Artocarpus, Moraceae): genetic characterization of critical germplasm[J]. Tree Genetics & Genomes,2015, 11(1): 1-26.
[15] GARDNER E M, LARICCHIA K M, MURPHY M,RAGONE D, SCHEFFLER B E, SIMPSON S, WILLIAMS E W, ZEREGA N J C. Chloroplast microsatellite markers for Artocarpus (Moraceae) developed from transcriptome sequences[J]. Applications in Plant Sciences, 2015, 3(9):1500049.
[16] BELLIS D F, MALAPA R, KAGY V, LEBEGIN S,BILLOT C, LABOUISSE J P. New development and validation of 50 SSR markers in breadfruit (Artocarpus altilis,Moraceae) by next-generation sequencing[J]. Applications in Plant Sciences, 2016, 4(8): 1600021.
[17] 彭丽娟, 牟柯澴, 张健伟, 赵兆, 山诗瑶, 薛新如, 朱丽,许琳玉, 李焕秀. 基于SSR 及ISSR 标记的菜豆遗传多样性分析[J]. 分子植物育种, 2022, 20(21): 7161-7173.
[18] 冷嘉文, 乔中全, 唐丽, 王晓明, 陈前欣, 邵雯雯. 基于SSR 分子标记的紫薇遗传多样性分析[J]. 湖南生态科学学报, 2021, 8(4): 1-7.
[19] 杨梦婷, 黄洲, 干建平, 徐君驰, 庞基良. SSR 分子標记的研究进展[J]. 杭州师范大学学报(自然科学版), 2019,18(4): 429-436.
[20] 吴刚, 胡丽松, 黄丽芳, 郝朝运, 谭乐和. 菠萝蜜种质资源遗传多样性的SRAP 分析[J]. 热带作物学报, 2015,36(8): 1385-1391.
[21] 敬丹, 骆翔, 陈利娜, 李好先, 唐丽颖, 曹尚根. 基于SSR分子标记的78 份核桃种质资源遗传多样性分析[J]. 江西农业学报, 2020, 32(6): 11-16.
[22] GARDNER E M, JOHNSON M G, PEREIRA J T, PUAD A S A, ARIFIANI D, WICKETT N J, ZEREGA N J C. Paralogs and off-target sequences improve phylogenetic resolution in a densely sampled study of the breadfruit genus (Artocarpus, Moraceae)[J]. Systematic Biology, 2021, 70(3):558-575.
[23] RAGONE D. Breadfruit Artocarpus altilis (Parkinson) Fosberg[M]. Rome: International Plant Genetic Resources Institute,1997: 14-19.
[24] RAGONE D. Chromosome numbers and pollen stainability of three species of Pacific Island breadfruit (Artocarpus,Moraceae)[J]. American Journal of Botany, 2001, 88(4):693-696.
[25] RAGONE D. Breadfruit: diversity, conservation and potential[ C]//I international symposium on breadfruit research and development, 2007, 757: 19-30.
