海南不同成熟期诺丽果的蛋白鉴定及功能分析
2023-08-14邱静宋媛王妮王佳人陈静陈光英赵振东何文英
邱静 宋媛 王妮 王佳人 陈静 陈光英 赵振东 何文英







摘 要:诺丽果(noni)是海南的一种特色水果,具有多种药效价值,但关于其植物蛋白的研究却很少。本研究采用基于BPP+酚提取法的双向电泳技术,通过优化实验条件,对花蕾期、半成熟及成熟期等3 个时期产自海南的诺丽果实进行蛋白质的提取分离,获得其植物蛋白指纹图谱;利用Image Master 5.0 软件分析凝胶图像筛选出高表达蛋白;再利用MALDI-TOF-MS 技术对这些蛋白酶解产物进行质谱鉴定;通过生物信息学手段对3 种诺丽果实的高表达蛋白初步进行蛋白功能分析。结果表明:花蕾期、半成熟及成熟诺丽果实分别有44、49 和40 个高表达蛋白,通过质谱鉴定,确定了其中分别有33、22 和39 个蛋白,从属不同植物功能的蛋白种类;不同成熟期的高表达蛋白差异较大,涉及叶绿体连接酶、半胱氨酸蛋白酶、蛋白激酶、泛素连接酶、生长素反应因子等多种蛋白质。该研究为进一步开发利用诺丽果的植物蛋白提供科学合理的理论指导。
关键词:诺丽果;不同成熟期;蛋白;功能分析
中图分类号:S759.83 文献标识码:A
诺丽(Noni)即海巴戟(Morinda citrifolia L.),又名热带海巴戟、海巴戟天、四季果等,属茜草科巴戟天属植物,是一种生长于热带及亚热带的多年生常绿阔叶灌木或小乔木,波利尼西亚土著将诺丽作为民间药物使用已有2000 多年历史,民间多用于治疗发烧、腹泻、便秘、哮喘、恶心呕吐、蚊虫叮咬及动物咬伤引起的感染等疾病[1-2]。
诺丽植株的多部位含有多种活性成分,近年来已从诺丽的根、茎、叶、花、果实、树皮、芯材等部位分离鉴定了300 余种化合物,主要包括黄酮、多糖、香豆素、蒽醌、环烯醚萜、生物碱、木脂素、香豆素、三萜、甾醇等化学成分[3]。现代药理学研究表明,诺丽具有抗炎、抗癌、抗氧化、抗焦虑、抗抑郁、降血脂、降血糖、肝保护、改善记忆、提高耐力、提高免疫力、减轻骨质疏松症、改善关节疼痛及活动度、对缺血性神经元损伤的保护作用、多巴胺双向调节等作用[4]。作为一种传统药食同源的植物,诺丽果实也是一种特色的热带水果,据报道,其中含有275 种以上的营养成分,除含抗氧化物、东莨菪碱、多醣体、及罕见的对人体健康非常重要的17 种虹甙类(环烯醚萜类)等化学成分以外,还包括人体所需的20 种必需氨基酸、9 种人体内无法自行生成的必需氨基酸;另含有丰富维生素B1、维生素B2、维生素B6、维生素B3、叶酸等14 种维生素,含有钾、钠、锌、钙、铁、镁、磷、铜和硒等16 种矿物质,硒含量为其他水果的10 倍以上[5-7]。
由于诺丽的高营养价值,早在2010 年诺丽果浆被中华人民共和国卫生部列入了新资源食品名单,诺丽种植在我国海南省和云南省已具备一定规模[3]。通过追踪不同产地诺丽果关键发育期主要活性物质的动态变化,不仅为诺丽果原材料的实时采收、质量控制以及缩短诺丽产品生产成本与周期提供科学依据[8],也可对不同成熟度诺丽果进行主要活性成分分析。国内最新报道了云南产诺丽果不同成熟度诺丽果中多糖、总黄酮、总酚、总蒽醌、莨菪亭、总皂苷、桃叶珊瑚苷、蛋白质含量测定的研究,结果表明成熟度与活性成分间具有一定的相关性[8]。
国外也有较多关于诺丽果实及种子的种植、营养价值、药理活性及机制的研究[9-11]。通过查阅国内外文献,发现虽然有较多关于诺丽化学成分提取及其生物或药理活性的研究,但还未见有诺丽不同成熟度的蛋白提取及其指纹图谱的建立或其中功能蛋白发现分析的报道。作为一种有较高营养价值及药理活性的药食同源植物,研究其所含植物蛋白名称、发现与其药物活性相关的关键蛋白,对深度了解诺丽果实、指导育种、进一步开发诺丽果实均具有非常重要的理论和实际意义。本研究利用蛋白质组学的双向电泳技术[12],获得海南不同成熟期的诺丽果蛋白图谱,并利用生物质谱及生物信息学技术,鉴定分析諾丽果关键发育期主要蛋白质的动态变化,为进一步深入研究和开发诺丽果提供科学合理的指导。
1 材料与方法
1.1 材料
花蕾期、半成熟及成熟诺丽果果实样品,均采集于海南海口东山镇诺丽果种植园(20°02′45.97″N,110°11′38.39″E),经海南师范大学生命科学院的陈玉凯副教授鉴定确为花蕾期、半成熟及成熟诺丽果果实(图1)。将采集回来的花蕾期、半成熟及成熟诺丽果样品用超纯水清洗干净,用锡箔纸包裹住放入保鲜袋中,于–80 ℃的冰箱中保存备用。
蛋白提取仪器及试剂:台式恒温振荡器(精宏);QL-866 涡旋混合器;SIGMA3-18K 低温超速离心机(德国SIGMA);超纯水系统(上海和泰);EYELA 摇床(东京理化);蛋白质等电聚焦仪(Ettan IPGphor3), 固相pH 梯胶条(pH3~10);Image scanner III 扫描仪(美国通用公司GE Healthcare);Image Master 5.0 凝胶图像分析软件;MultiTemp IV 恒温循环器;UV-2700 紫外分光光度计(日本岛津);KQ2200E 型超声波清洗仪(曙峰企业);GM-0.33A 型隔膜真空泵(津腾);Voyager-DE PRO ABI4700 时间飞行质谱仪(美国ABI 公司)。所有试剂均为国产分析纯试剂,所有实验用水为超纯水。
1.2 方法
1.2.1 BPP+酚抽提法提取蛋白 (1)取出样品,切成小块, 置于预冷的干净研钵中, 加入少许 PVPP 粉末和少许二氧化硅粉末,在液氮环境中研磨到粉末状;称取3 g 研磨好的样品粉末,加入至10 mL 离心管中,再加入10 mL BPP 提取缓冲液,充分涡旋震荡10 min;再加入10 mLTris饱和酚,涡旋10 min;在SIGMA3-18K 低温超速离心机中离心15 min(4 ℃,转速:16000×g);移取上层清液转移至干净离心管中,加入5 mLBPP 提取缓冲液,涡旋震荡10 min 后离心15 min;再移取3 mL 上层清液转移至干净离心管中,加入15 mL 预冷过饱和硫酸铵甲醇溶液(AM 沉淀剂),置于–20 ℃冰箱中沉淀12 h 以上;从冰箱中取出离心管,用 AM 沉淀剂配平后离心15 min弃上清液;每管再加入2 mL 预冷甲醇,去上清液,涡旋均匀,重复2 次;再分别加入 2 mL 预冷丙酮,重复2 次转移及离心步骤;去除上清液,分别加入适量DTT,混合均匀,按10 mg/mL 加入适量裂解液,于20 ℃恒温溶解2 h 以上;待蛋白完全溶解到裂解液后,离心,取上清液,测定蛋白浓度。
(2)标准曲线法测定蛋白质含量。称取1 mg牛血清蛋白充分溶解于1 mL 超纯水中,配制成1 mg/mL 的标准牛血清蛋白储备液。分别准确量取1 mg/mL 的标准牛血清蛋白储备液若干体积,用超纯水配制成0、2、4、6、8、10 μg/mL 的牛血清蛋白标准溶液。利用紫外分光光度计,测定在595 nm 处的吸光度值,根据朗伯-比尔定律,以牛血清蛋白标准溶液的浓度为横坐标,吸光度值为纵坐标,制作标准曲线。
1.2.2 蛋白质含量测定 采用Bradford 法测定样品的蛋白浓度[13], 使用紫外分光光度计, 在595 nm 的波长下测定花蕾期、半成熟及成熟诺丽果蛋白提取液的紫外吸光度值,根据标准曲线方程,计算样品中蛋白质的含量。
1.2.3 单向电泳实验 根据吴秀丽等[14]的方法进行单向电泳实验,确定双向电泳的最佳上样浓度。
1.2.4 双向电泳实验 移取含有1.3 mg 蛋白质的提取液,用蛋白裂解液稀释至样品体积为455 μL,加5 μL IPG Buffer 于离心管中,轻轻震荡使其混合均匀,转移至IPG 胶条(pH 3~10)上,20 ℃恒温水化18 h 以上;保持室温20 ℃左右,设置聚焦极限电流为每根胶条50 μA,电压参数如表1所示,进行一向等电聚焦;再将胶条放置在摇床上平衡15 min;在制备好的聚丙烯酰胺凝胶上进行二向垂直电泳操作,设置参数为:每张胶片5 W进行预电泳1 h,每张胶片7 W 至电泳结束;再进行凝胶染色和脱色、凝胶扫描和用ImageMaster 软件分析凝胶图谱[14],筛选出高表达的蛋白并进行编号。
1.2.5 生物质谱鉴定及分析 挖取高表达蛋白质,分别用150 μL 超纯水、150 μL 脱色液和100μL 乙腈处理,放置在摇床上摇荡30 min,转速设置为150 r/min,摇荡后吸出离心管中的水,以上步骤重复3 次,直至蛋白粒变为无色,置于室温下风干;将蛋白样品离心后加入适量胰蛋白酶,放置在4 ℃冰箱中1 h 左右;再将其放置在PCR 仪上酶解蛋白(37 ℃,13 h);设置转速7000×g,离心5 min(20 ℃),备用;利用MALDI-TOF-MS进行蛋白鉴定;将所得的质谱数据通过MascotDistiller 软件进行分析,再利用Matrix Science(http://www.matrixscience.com)网站进行搜库比对,确定蛋白的名称、种类及其他信息。
2 结果与分析
2.1 不同成熟期诺丽果蛋白提取
通过Bradford 法测定3 种诺丽果的蛋白浓度,在波长为595 nm 处通过测定系列梯度浓度的标准牛血清蛋白溶液的吸光度值,建立标准曲线方程(未显示),计算得出花蕾期、半成熟及成熟诺丽果蛋白溶液的浓度分别为6.808、11.30、5.89 mg/mL。
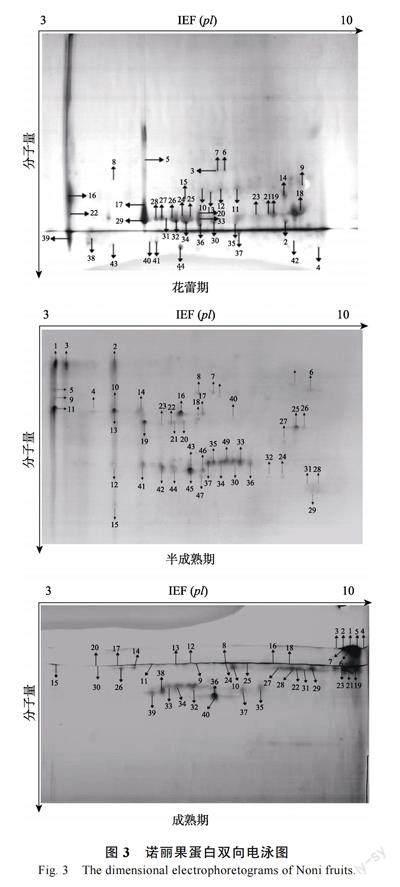
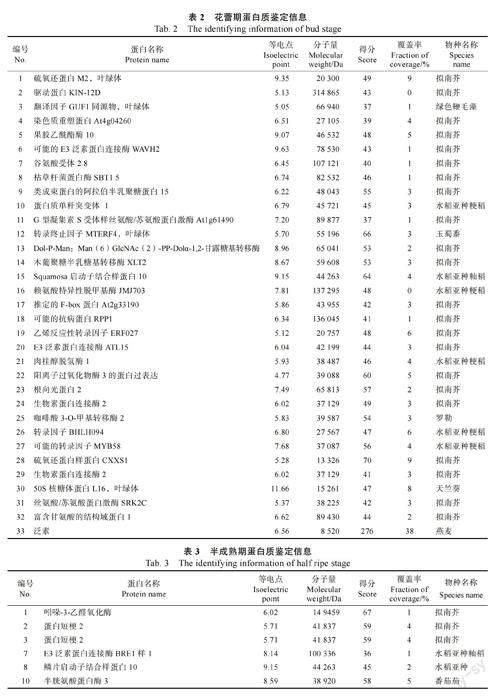
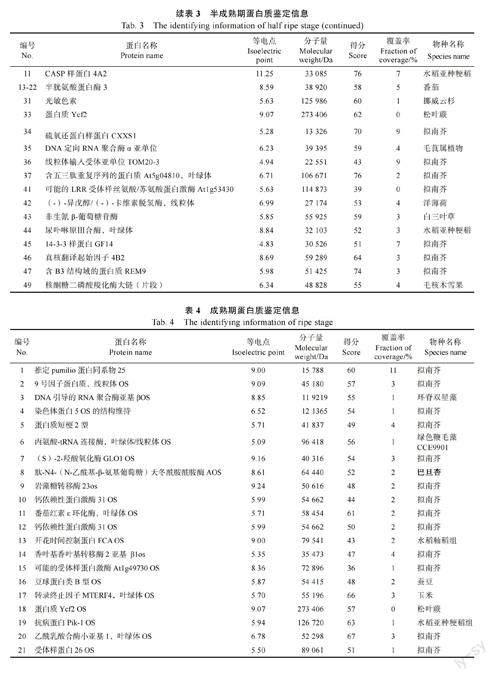
为获得双向电泳合适的上样浓度,进行单向电泳实验,结果如图2 所示。不同成熟期诺丽果的蛋白分子量介于15~130 kDa 之间,花蕾期的蛋白主要在17、20、25 kDa 左右,半成熟期的蛋白主要在17、20、33 kDa 左右,而成熟期的蛋白则主要集中在15、20、25、33 kDa 左右。另外,从图2 可以清晰看出样品的蛋白条带,说明不同成熟期诺丽果实蛋白质未降解,可以进行后续双向电泳实验操作。
在优化实验条件下,通过双向电泳实验可以得到清晰明顯、分离效果很好的蛋白凝胶图谱,再经ImageMaster 软件分析在蛋白凝胶图谱上标记高表达的蛋白点。花蕾期、半成熟及成熟诺丽果实分别有44、49 和40 个高表达蛋白(图3)。
2.2 不同成熟期诺丽果蛋白质谱鉴定
将不同成熟期诺丽果中的若干高丰度蛋白质点,经过挖点、酶解和提取处理后,利用基质辅助激光解吸附电离飞行时间质谱仪(MALDITOF-MS)对其进行分析和鉴定,并设置参数以基质峰、酶自切峰进行校正,测定各蛋白肽段的质谱数据,再通过植物蛋白质数据库进行对比来确定这些蛋白的名称、种类等信息,最终成功鉴定出花蕾期有33 个蛋白点、半成熟期有23 个蛋白点、成熟期有39 个蛋白点,结果分别如表2、表3 及表4 所示。
2.3 不同成熟期诺丽果蛋白功能分析
以上通过利用双向电泳实验和MALDI-TOFMS生物质谱技术,对海南不同成熟期的诺丽果实中植物蛋白的提取和鉴定实验,成功匹配了花蕾期、半成熟及成熟诺丽果实分别有44、49 和40 个高表达蛋白,通过质谱鉴定,相应确定了分别有33、49 及39 个蛋白,从属不同植物功能的蛋白种类,对这些差异表达的蛋白进行了简单的生物学功能分析,详见表5~表7。花蕾期和半成熟期相同的蛋白有:硫氧还蛋白、驱动蛋白、叶绿体、丝氨酸/苏氨酸蛋白激酶、蛋白连接酶、Squamosa 启动子结合样蛋白、转录因子;花蕾期和成熟期相同的蛋白有:转移酶、可能的抗病蛋白、F-box 蛋白;半成熟期和成熟期相同的蛋白有:蛋白短梗、半胱氨酸蛋白酶、转移酶、叶绿体、蛋白质Ycf2。
3 讨论
诺丽果不同成熟期的蛋白相差较大,表现在众多的生长、发育及遗传功能方面。花蕾期的23号根向光蛋白与半成熟期的31 号光敏色素均与植物的光感应相关,由于开花是植物从营养生长到生殖生长的一个重要转折点,直接影响着种子植物生育期的早晚、授粉及种子发育的最佳时期,这暗示不仅可通过人为调节光的某些特性来调控花蕾期诺丽果的根向光蛋白,从而促使花蕾期诺丽果尽快开花,也可从不同成熟的诺丽果提取根向光蛋白并用于科学培植诺丽果[91]。此外,发现花蕾期的8 号蛋白枯草杆菌蛋白酶[92]、18 号蛋白可能的抗病蛋白RPP1[93]与成熟期的19 号抗病蛋白Pik-1[94],均与植物抵抗病菌相关,也表明这3种抗病蛋白可有望开发为高效、低毒的生物农药。
本研究发现部分蛋白与动物体内的蛋白相同,具有不同的生物学或药学功能,如花蕾期的1、28 号蛋白和半成熟期的34 为硫氧还原蛋白,作为一种小的热稳定酸性蛋白,是细胞中重要的氧化还原蛋白,具有调节细胞抗氧化、抗胁迫、细胞凋亡、DNA 结合转录因子活性等功能,说明在诺丽果花蕾期到半成熟期的发育过程中,可能在抗氧化应激、阻止线粒体介导的细胞死亡、调节某些基因的表达等方面起关键作用[15];作为人体重要的内源性抗氧化系统之一,硫氧还原蛋白被研究证明具有出色的抗氧化活性或调节众多细胞生存相关信号通路,从而抵抗活性氧损伤[95]。
花蕾期的2 号、成熟期的24 号蛋白为驱动蛋白,作为一类重要的微管调节蛋白,可能在诺丽果的神经元的发育、纺锤体的组装和染色体的分离过程中起着重要的作用[16],人体驱动蛋白是分子马达的一种,其通过结合和水解ATP,导致颈部发生构象变化,促使驱动蛋白沿着微管“行走”,完成细胞中货物运输的工作,能为细胞生命活动提供动力[96]。花蕾期中编号6、20、24 和29 均为蛋白连接酶,其中编号6 和20 为E3 泛素蛋白连接酶,其广泛参与细胞内的代谢过程,比如细胞增殖、细胞周期调控、细胞的凋亡,人体这些酶的调节异常与多种疾病有关,如神经系统疾病与恶性肿瘤等[23]。编号24 和29 为生物素蛋白连接酶,通过调节微生物的乙酰辅酶A 羧化酶和丙酮酸羧化酶,控制重要代谢途径的关键反应,从而有效控制微生物生长,也是极具开发前景的抗菌新靶点[24];花蕾期的18、成熟期的19 抗病蛋白能识别病原体及微生物,并触发免疫,对抗病原体以及微生物的侵扰[35]。花蕾期17、成熟期的34 号为F-box 蛋白,作为泛素连接酶复合物SCF的重要组成物,可能参与调控诺丽果的整个细胞周期,对人体诊治肿瘤以及生殖方面的疾病也有重要的作用[34]。成熟期的8 号天冬酰胺酰β-羟化酶是一种能催化L-天冬酰胺水解生成L-天冬氨酸和氨的酰胺基水解酶,广泛存在于植物、动物和微生物中;人类天冬氨酰参与并调节恶性肿瘤细胞的侵袭及转移,在多种恶性肿瘤中过表达,参与并调节恶性肿瘤细胞的侵袭及转移,并可介导抗肿瘤免疫反应,在肿瘤靶向治疗、免疫治疗中的作用逐渐受到重视[67]。成熟期的10 级12 号蛋白为钙依赖性蛋白激酶,在Ca2+介导的信号转导中起重要作用,可能参与调控成熟期诺丽果的植物激素信号通路、生物胁迫反应及其根、茎、叶或种子发育等过程[69];人体钙依赖性蛋白激酶对胆管癌细胞HuCCT1 血管的生成有一定的影響和调控[97]。半成熟期的10、13、14、16、18、19、20、21、22 和成熟期的29 号为半胱氨酸蛋白酶,是植物中重要的蛋白酶家族之一,可能参与半成熟期和成熟期诺丽果种子萌发、幼苗发育、胁迫响应和组织分化衰老等过程[46];人体半胱氨酸蛋白酶抑制剂S 对于胃癌的鉴别具有重要作用,对极有可能作为新的肿瘤标志物以及它对于胃肠癌诊断具有极其重要的参考价值[98]。
诺丽果是一种药食同源的植物,虽然大部分现有的研究是关于诺丽果不同部位化学成分的提取及在营养食品等方面的应用,但从蛋白质组学的方法挖掘诺丽果的科学及实用价值的国内外研究却很少有。本研究用蛋白质组学揭示了不同成熟期果实的蛋白种类及功能,以上结果表明在不同成熟期的诺丽果中大多数蛋白、激素以及酶均存在着药学作用,在一定程度上可补充说明诺丽果的药理活性,对后续深入了解诺丽果实、指导育种以及进一步开发诺丽果实提供理论依据。
参考文献
[1] DE LA CRUZ-SÁNCHEZ N G, GÓMEZ-RIVERA A,ALVAREZ-FITZ P, VENTURA-ZAPATA E, PÉREZGARCÍA M D, AVILÉS-FLORES M, GUTIÉRREZROMÁN A S, GONZÁLEZ-CORTAZAR M. Antibacterial activity of Morinda citrifolia Linneo seeds against Methicillin-Resistant Staphylococcus spp[J]. Microbial Pathogenesis,2019, 128: 347-353.
[2] 晏永球, 童应鹏, 陆雨, 江石平, 江瑜, 左丽敏, 杨君, 邵云东, 王平. 诺丽的化学成分及药理活性研究进展[J]. 中草药, 2017, 48(9): 1888-1905.
[3] 李戈, 张丽霞, 牛迎凤, 元超, 李小花, 赵俊凌. 不同来源地诺丽果质量分析[J]. 中华中医药杂志, 2016, 31(9):3766-3769.
[4] 刘金莲, 张睿, 刘晔斌, 侯燕, 王林元, 王淳, 张建军, 高学敏. 诺丽的文献研究及中药药性理论探讨[J]. 中国中药杂志, 2020, 45(5): 984-990.
[5] 李法营, 蓝增全, 刘昌芬, 李青红. 诺丽研究进展(一)—国内外研究进展[J]. 安徽农业科学, 2009, 37(32):15819-15821.
[6] 王丽. 诺丽果的营养与保健功能[J]. 中国妇幼健康研究,2017, 28(4): 139-140.
[7] 周城, 王根女. 诺丽功能活性和应用研究进展[J]. 农业研究与应用, 2019, 32(Z1): 38-43.
[8] 景瑞雪, 孔凡, 白沙沙, 田亮, 崔亚娟, 刘洋. 西沙诺丽果关键发育期主要活性物质及其含量变化分析[J]. 中国食物与营养, 2020, 26(12): 51-55.
[9] 徐荣, 许木果, 姜士, 杨焱, 付鎵榕, 岩利. 不同成熟度诺丽果主要活性成分分析[J]. 热带农业科技, 2022, 45(2):30-35, 40.
[10] JAHURUL M H A, PATRICIA M, SHIHABUL A,NORAZLINA M R, GEORGE M R, NOORAKMAR A W,LEE J S, JUMARDI R, JINAP S, ZAIDUL I S M. A review on functional and nutritional properties of noni fruit seed(Morinda citrifolia L.) and its oil[J]. Food Bioscience, 2021,41: 101000.
[11] HAN H S, SONG K B. Noni (Morinda citrifolia) fruit polysaccharide films containing blueberry (Vaccinium corymbosum)leaf extract as an antioxidant packaging material[J].Food Hydrocolloids, 2021, 112: 106372.
[12] 邱智敏, 郑碧娟, 陈辉, 何艺凡, 陈世品. 油茶雌蕊蛋白双向电泳分离体系的建立[J]. 热带作物学报, 2016, 37(3):548-554.
[13] BRADFORD M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry,1976, 72(1/2): 248-254.
[14] 吴秀丽, 王旭初, 段红叶, 李建玲, 何文英. 海南不同产地草豆蔻的差异蛋白分析[J]. 热带农业科学, 2018, 38(1):53-61.
[15] 谢婧. 豌豆根瘤菌硫氧还蛋白基因trxs 的功能研究[D].武汉: 中南民族大学, 2019.
[16] 蒋蕾蕾, 吴作基, 王春光. 微管解聚型驱动蛋白的结构与功能(英文)[J]. 细胞生物学杂志, 2008(2): 137-141.
[17] 吴新辉, 郭钰滢, 夏楠, 唐杨, 郭泓伯, 王洋, 杜恩在. 中国东部城市森林阔叶树种叶绿素含量空间特征及其影响因素[J]. 北京师范大学学报(自然科学版), 2022, 58: 1-8.
[18] 闫凯歌. 核糖体翻译因子EF-G 和EF4 的结构和功能研究[D]. 北京: 清华大学, 2016.
[19] 何晓芳. 拟南芥叶绿体定位的mTERFs 家族蛋白的功能分析[D]. 上海: 上海师范大学, 2016.
[20] 王静文, 王兴军, 马长乐, 李膨呈. 植物核糖体应激响应机制研究进展[J]. 植物学报, 2022, 57(1): 80-89.
[21] 刘萌. 青霉木质纤维素酶转录调控互作蛋白初探[D]. 济南: 山东大学, 2017.
[22] 刘艳丽, 马林龙, 曹丹, 金孝芳, 冯琳, 龚自明. 茶树果胶乙酰酯酶家族成员的鉴定和生物信息学分析[J]. 茶叶科学, 2019, 39(5): 521-529.
[23] 谢少利, 幸天勇, 邓世山. 泛素蛋白连接酶E3 介导的EMT 在肿瘤浸润与转移中的作用[J]. 川北医学院学报,2015, 30(5): 732-737, 742.
[24] 杨朝福, 张奶玲, 王菊, 贺艳斌. 新型抗菌靶点BPL 抑制剂的研究进展[J]. 中国药物化学杂志, 2022, 32(7): 538-547.
[25] 张启平. NbGLRs 基因家族鉴定及其调控烟草系统性抗性防御TMV 侵染的机制[D]. 扬州: 扬州大学, 2022.
[26] 彭帅. 地衣芽孢杆菌W10 枯草杆菌蛋白酶Sp1 抗菌和诱导植物抗病性机理研究[D]. 扬州: 扬州大学, 2021.
[27] 李家宝. 棉花类成束蛋白阿拉伯半乳聚糖蛋白基因(GhFLA)的克隆与功能分析[D]. 重庆: 西南大学, 2007.
[28] 孙玉芬. 水稻分蘖基因MONOCULM1 的多态性研究[D].北京: 中国科学院遗传与发育生物学研究所, 2008.
[29] 赵永, 温李, 谢志明, 高金秋, 柴泽斌, 汪雨琪, 李涵冰,李洋, 丛建民. 蓖麻丝氨酸/苏氨酸蛋白激酶RcPK2 基因克隆及蛋白纯化[J]. 分子植物育种: 1-10[2023-05-15].
[30] 刘叩晗. 禾谷镰刀菌中甘露糖基轉移酶复合体的功能研究[D]. 合肥: 安徽农业大学, 2021.
[31] 解敏敏, 晁江涛, 孔英珍. 参与木葡聚糖合成的糖基转移酶基因研究进展[J]. 植物学报, 2015, 50(5): 644-651.
[32] 汪先菊, 张绍宇, 徐泽, 石佩, 康同洋, 李琴, 赵彩平. 桃SPL 基因家族成员鉴定及其基因表达与果实发育的相关性分析[J]. 农业生物技术学报, 2020, 28(9): 1521-1534.
[33] MARTINEZ-GAMERO C, MALLA S, AGUILO F. LSD1:Expanding functions in stem cells and differentiation[J].Cells, 2021, 10(11): 3252.[34] LIU Q, GUO X H, CHEN G P, ZHU Z G, YIN W C. Silencing SlGID2, a putative F-box protein gene, generates a dwarf plant and dark-green leaves in tomato[J]. Plant Physiology and Biochemistry, 2016, 109: 491-501.
[35] 閆佳, 刘雅琼, 侯岁稳. 植物抗病蛋白研究进展[J]. 植物学报, 2018, 53(2): 250-263.
[36] 黄荣峰, 张执金, 王娟, 张丽霞. 乙烯反应因子(ERF)调控植物耐盐性的分子基础[C]//全国植物生物学大会论文集,2013: 240-241.
[37] LACHAGARI V R, BODANAPU R, CHAKRAVARTTY N, LEKKALA S P, LALAM K, KURIAKOSE B,VEMIREDDY L R, VELAYUTHAM D, THOMAS G,GUPTA S. Uncovering genome wide novel allelic variants for eating and cooking quality in a popular Indian rice cultivar,Samba Mahsuri[J]. Current Plant Biology, 2019, 18:100111.
[38] 任思齐, 蔚丽慧, 魏美玲, 郑佳敏, 何茂硕, 郑恺, 范桂枝.罗布麻MYB 转录因子家族生物信息学分析[J]. 分子植物育种, 2021, 20(19): 6371-6381.
[39] 蓝雨纯. 灰霉菌胁迫下小立碗藓肉桂醇脱氢酶1 基因功能初探[D]. 贵阳: 贵州师范大学, 2020.
[40] 何之常, 朱英国. 籼、粳两系杂交水稻幼根中吲哚乙酸氧化酶和阳离子过氧化物酶的初步研究[J]. 江苏农业学报,1996(1): 52-55.
[41] 常立, 文国琴. 植物蓝光受体研究进展[J]. 生物技术通讯,2004(2): 169-171.
[42] SHOWALTER A M. Structure and function of plant wall protein[J]. Plant Cell, 1993, 5: 9-23.
[43] GAO C Y, TANG D Z, WANG W. The role of ubiquitination in plant immunity: fine-tuning immune signaling and beyond[J]. Plant and Cell Physiology, 2022, 63(10): 1405.
[44] 王金祥. 绿豆下胚轴不定根形成不同时期的激素调控[D].广州: 华南师范大学, 2002.
[45] MOUCHEL C F, BRIGGS G C, HARDTKE C S. Natural genetic variation in Arabidopsis identifies BREVIS RADIX, a novel regulator of cell proliferation and elongation in the root[J]. Genes & Development, 2004, 18(6): 700-714.
[46] 闫龙凤, 杨青川, 韩建国, 刘志鹏. 植物半胱氨酸蛋白酶研究进展[J]. 草业学报, 2005(5): 11-19.
[47] REYT G, CHAO Z, FLIS P, SALAS-GONZÁLEZ I, CASTRILLO G, CHAO D Y, SALT D E. Uclacyanin proteins are required for lignified nanodomain formation within casparian strips[J]. Current Biology, 2020, 30(20): 4103-4111.
[48] 岳玲琦, 邢巧娟, 张晓兰, 梁雪, 王乾, 齐红岩. 光敏色素互作因子在植物抵御逆境胁迫中的作用进展[J]. 园艺学报, 2021, 48(4): 632-646.
[49] 张媛媛. 光敏色素的结构及其信号调控机制[J]. 湖北农业科学, 2020, 59(4): 5-10.
[50] PETER E, WALLNER T, WILDE A, GRIMM B. Comparative functional analysis of two hypothetical chloroplast open reading frames (ycf) involved in chlorophyll biosynthesis from Synechocystis sp. PCC6803 and plants[J]. Journal of Plant Physiology, 2011, 168(12): 1380-1386.
[51] HO T H, HONG M K, NGO H P T, KANG L W. Expression, crystallization and preliminary X-ray crystallographic analysis of DNA-directed RNA polymerase subunit L from Thermococcus onnurineus NA1[J]. Acta Crystallographica Section F: Structural Biology Communications, 2014, 70(5):639-642.
[52] FAN A C, KOZLOV G, HOEGL A, MARCELLUS R C,WONG M J, GEHRING K, YOUNG J C. Interaction between the human mitochondrial import receptors Tom20 and Tom70 in vitro suggests a chaperone displacement mechanism[J]. Journal of Biological Chemistry, 286(37):32208-32219.
[53] TIAN T, IKEDA J I, WANG Y, MAMAT S, LUO W,AOZASA K, MORII E. Role of leucine-rich pentatricopeptide repeat motif-containing protein (LRPPRC) for anti-apoptosis and tumourigenesis in cancers[J]. European Journal of Cancer,2012, 48(15): 2462-2473.
[54] ZHAN J R, SHOU C, ZHENG Y C, CHEN Q, PAN J, LI C X, XU J H. Discovery and engineering of bacterial(−)- isopiperitenol dehydrogenases to enhance (?)-menthol precursor biosynthesis[J]. Advanced Synthesis & Catalysis, 2021, 363(16): 3973-3982.
[55] 李晨辉. 产β-葡萄糖苷酶菌株的筛选鉴定及亚麻籽饼粕脱毒工艺的研究[D]. 兰州: 兰州大学, 2017.
[56] MORANT A V, JØRGENSEN K, JØRGENSEN C,PAQUETTE S M, SÁNCHEZ-PÉREZ R, MØLLER B L, BAK S. β-Glucosidases as detonators of plant chemical defense[J]. Phytochemistry, 2008, 69(9): 1795-1813.
[57] 曹凤. 小麦尿卟啉原Ⅲ合酶基因克隆、序列分析及其在大肠杆菌中的表达[D]. 杨凌: 西北农林科技大学, 2007.
[58] ORMANCEY M, THULEAU P, MAZARS C, COTELLE V. CDPKs and 14-3-3 proteins: emerging duo in signaling[J].Trends in Plant Science, 2017, 22(3): 263-272. (in Chinese)
[59] 周红. 水稻Pip2-1 基因在RSV侵染过程中的功能研究[D].合肥: 安徽农业大学, 2013.
[60] 王莲哲, 向丽君, 李建康, 王渊, 周帅, 李冰冰. 小麦B3类转录因子REM-1 基因的克隆与表达分析[J]. 分子植物育种, 2019, 17(15): 4853-4858.
[61] 董尚胜, 童启庆. 茶树叶片内核酮糖二磷酸羧化酶酶蛋白水平的研究[J]. 浙江农业大学学报, 1992, 18(S): 40-44.
[62] RYDER S P. Pumilio RNA recognition: the consequence of promiscuity[J]. Structure, 2011, 19(3): 277-279.
[63] 王硕. 植物细胞器基因组变异与进化机制研究[D]. 昆明:昆明理工大学, 2016.
[64] 王斌, 刘连瑞. RNA 聚合酶的模板特异性[J]. 遗传学报,1986(4): 254-258.
[65] 计文. 水稻染色体结构维持蛋白SMC1 及SMC3 在DSB修复中的功能研究[D]. 扬州: 扬州大学, 2019.
[66] 苏晨, 余露山, 姚彤炜, 曾苏. 重组大鼠长链2-羟酸氧化酶的克隆与表达[J]. 中国药学杂志, 2013, 48(5): 331-336.
[67] 怀琪琳, 韩连奎, 许川, 梅宏. 天冬氨酰-(天冬酰胺酰)β-羟化酶在肿瘤中的作用研究进展[J]. 中国医刊, 2021,56(3): 253-257.
[68] 徐佳璐, 庞秀秀, 燕小梅. 岩藻糖基转移酶在肝癌中的作用[J]. 生命的化学, 2020, 40(9): 1564-1570.
[69] 何乐平, 张蕾. 植物钙依赖性蛋白激酶及其相关蛋白激酶(CDPKs/CRKs) 的研究进展[J]. 安徽农业科学, 2020,48(18): 26-30, 51.
[70] 王春菲, 贾倩云, 贾真真, 郭玉涛, 许锐, 孙露, 高书文,梁智慧, 郭敬功. 番茄红素ε-环化酶调节烟草抗旱性的研究[J]. 河南大学学报(自然科学版), 2019, 49(4): 444-449.
[71] 李浩杰. 芸薹属基因组中开花时间基因的鉴定和进化分析[D]. 成都: 四川农业大学, 2013.
[72] 刘谈, 吕朝耕, 王升, 杨婉珍, 郭兰萍. 基于新疆紫草转录组的对羟基苯甲酸香叶基转移酶(PGT)基因的挖掘及生物信息学分析[J]. 中国中药杂志, 2016, 41(8): 1422-1429.
[73] 才晓溪, 沈阳, 胡冰霜, 王研, 陈悦, 孙明哲, 贾博为, 孙晓丽. 野生大豆类受体蛋白激酶基因GsCBRLK 超量表达提高水稻耐盐碱性[J]. 植物生理学报, 2020, 56(12):2683-2694.
[74] 百度百科. 豆球蛋白[EB/OL]. [2023-05-09]. https://baike.baidu.com/item/%E8%B1%86%E7%90%83%E8%9B%8B %E7%99%BD/5600985?fr=aladdin.Baidu Baike. legumin[EB/OL]. [2023-05-09]. https://baike. baidu.com/item/%E8%B1%86%E7%90%83%E8%9B%8B% E7%99%BD/5600985?fr=aladdin. (in Chinese)
[75] 黄淑颜, 丘式浚, 陈家逸, 陈健璞, 李晓云, 李玲. 线粒体转录终止因子(mTERF)蛋白家族的研究进展[J]. 生命科学研究, 2016, 20(5): 455-459.
[76] 杨程惠子, 唐先宇, 李威, 夏石头. NLR 及其在植物抗病中的调控作用[J]. 植物学报, 2020, 55(4): 497-504.
[77] 孙笑非, 黄星, 陈博, 李顺鹏, 何健. 抗甲磺隆假单胞菌的分离及其乙酰乳酸合酶的大小亚基ilvIH 基因的克隆和表达[J]. 微生物学报, 2008(11): 1493-1498.
[78] 杨诗勤, 张灵芝, 王旭东, 蒋涛, 敖雁, 彭运鹏, 姚骅珊,任胜杰. 水稻类受体蛋白激酶基因OsBDG1 的克隆及功能初步分析[J/OL]. 分子植物育种, 2022:1-9[2023-05-15].
[79] 孙悦. NADH脱氢酶调控丝状真菌Podospora anserina 降解木质纤维素及子实体发育研究[D]. 深圳: 深圳大学, 2020.
[80] 杨从文, 杨瑜, 王艳霞, 费纪涛, 林英. 昆虫醛氧化酶研究进展[J]. 蚕业科学, 2010, 36(4): 656-666.
[81] 刘瑞娥, 胡长贵, 孙玉强. 植物生长素反应因子研究进展[J]. 植物生理学报, 2011, 47(7): 669-679.
[82] 胡小倩, 张颖翌, 李鑫, 闫海芳. 植物中Remorin 蛋白的研究进展[J]. 生物技术通报, 2020, 36(8): 136-143.
[83] 童彬. 青霉菌灭活菌丝体多肽提取物诱导植物抗病过程中脱落酸对胼胝质沉积的影响初探[D]. 昆明: 云南大学,2019.
[84] 孔荣荣. 水稻DEAD-box RNA 解旋酶蛋白基因TCD33 功能研究[D]. 上海: 上海师范大学, 2019.
[85] 孙莹莹, 马雪山, 苏迎春. 早期胚胎组蛋白H3K9me3 与H3K27me3 的研究进展[J]. 中华医学遗传学杂志, 2020,37(11): 1296-1300.
[86] 吴子豪. UDP-葡萄糖基转移酶基因EDR1 调控水陆稻胚乳发育和非生物胁迫抗性[D]. 合肥: 中国科学技术大学,2021. (in Chinese)
[87] 汪晗. 表观遗传与橡胶树自根幼态无性系和其供体之间的差异产生的分子机制初探[D]. 海口: 海南大学, 2014.
[88] 张琳, 彭连伟. 叶绿体ATP 合酶生物发生的研究进展[J].植物生理学报, 2019, 55(6): 703-710.
[89] 唐佳佳. 枇杷果胶酯酶基因克隆及表达特性分析[D]. 重庆: 西南大学, 2014.
[90] GERASYMENKO I M, SAKHNO L A, KYRPA T N,OSTAPCHUK A M, HADJIEV T A, Goldenkova-Pavlova I V, Sheludko Y V. Characterization of Nicotiana tabacum plants expressing hybrid genes of cyanobacterial Δ9 or Δ12 acyl-lipid desaturases and thermostable lichenase[J]. Russian Journal of Plant Physiology, 2015, 62(3): 283-291.
[91] ZHAO F, LYU X G, JI R H, LIU J, ZHAO T, LI H Y, LIU B, PEI YA X. CRISPR/Cas9-engineered mutation to identify the roles of phytochromes in regulating photomorphogenesis and flowering time in soybean[J]. The Crop Journal, 2022, 10(6):1654-1664.
[92] EKCHAWENG K, KHUNJAN U, CHURNGCHOW N.Molecular cloning and characterization of three novel subtilisin-like serine protease genes from Hevea brasiliensis[J].Physiological and Molecular Plant Pathology, 2017, 97:79-95.
[93] ZHANG X, DONG X N. Life-or-death decisions in plant immunity[J]. Current Opinion in Immunology, 2022, 75:102169.
[94] ZHOU Y, WAN TAO, YUAN B, LEI F, CHEN M J,WANG Q, HUANG P, KOU S Y, QIU W X, LIU L. Improving rice blast resistance by mining broad-spectrum resistance genes at Pik locus[J]. Rice Science, 2022, 29(2):133-142.
[95] 孙伊人, 杨子琪, 王欣然, 钱永军. 硫氧还蛋白-1: 预防心脏相关活性氧损伤的潜在靶点[J/OL]. 中国胸心血管外科临床杂志, 2022: 1-5. [2023-05-15]. http://kns.cnki.net/kcms/
[96] 张婷. 驱动蛋白侧向运动机制研究[D]. 呼和浩特市: 内蒙古大学, 2021.
[97] 陆路, 黎东明, 吴晓勇, 王学国, 王太成, 赵红岩, 宋文渊.胆管癌组织钙依赖性蛋白激酶表达及其对HuCCT1 血管生成的调控机制[J]. 中华实用诊断与治疗杂志, 2021,35(4): 335-339.
[98] 吴明兰, 林美花, 翟优, 吕朵, 张乔, 吴国兰, 冯立民, 申屠建中. 半胱氨酸蛋白酶抑制劑S 在胃肠癌中的诊断价值研究[J]. 中国临床药理学杂志, 2021, 37(5): 503-506.
