三华李ANS 基因克隆及其在果实采后转色过程中的表达分析
2023-08-11许薇陈振林刘英健劳琪珍宋慕波
许薇,陈振林,刘英健,劳琪珍,宋慕波
(1.贺州学院食品科学与工程技术研究院,广西 贺州 542899;2.大连工业大学食品学院,辽宁 大连 116034)
三华李(Prunus salicinaLindl.cv.Sanhua)属于蔷薇科李亚科,是华南地区著名特色水果,原产自广东翁源县,后逐渐扩展至南方各省份[1,2]。三华李果实为圆形或近圆形,果肉紫红色,肉质爽脆,酸甜可口,营养价值高[3]。 三华李成熟果实中富含花青素,这是其重要特点[4];有研究表明三华李中总花青素含量仅次于黑布李,花青素主要成分为矢车菊素[5]。 目前针对三华李果实花青素合成的相关研究较少。
花青素是植物中广泛存在的一种水溶性天然色素,属于类黄酮化合物,在新鲜的蔬菜和水果中大量存在,使其呈现出明亮的颜色,从而吸引消费者;同时,花青素类物质也具有抗氧化、清除自由基等多种生理活性功能[6-9]。 花青素的合成主要分3 个阶段:第一阶段是苯丙氨酸经过多步反应生成4-香豆酰CoA;第二阶段是4-香豆酰CoA在系列酶的催化作用下生成黄酮类物质(二氢槲皮素、二氢杨梅素)和黄酮醇;第三阶段是二氢槲皮素、二氢杨梅素和黄酮醇这三类物质在二氢黄酮醇-4-还原酶(dihydroflavonol-4-reductase,DFR)的作用下生成无色的花青素(无色矢车菊素、无色天竺葵素和无色翠雀素),然后花青素合成酶(ANS)将无色的花青素转化为相应的花青素类物质(矢车菊素、天竺葵素和翠雀素),再经过糖基化、酰基化、甲基化等过程的修饰作用,最终形成稳定可见的花青素[10]。 可见,ANS 是花青素生物合成途径后期的关键酶,能催化无色花青素脱水氧化形成有色花青素。 编码ANS 的基因首先在紫苏中被克隆[11],在多数植物中,ANS由一个小基因家族所编码。 已有研究发现,在参薯[12]、紫色不结球白菜[13]、苹果梨[14]和凤丹牡丹[15]中ANS基因的表达与组织中花青素含量呈正相关。 然而,草莓果实从转红到完全成熟的过程中,虽然花青素含量呈上升趋势,但ANS基因表达量却有所下降[16]。 葡萄果实经乙醇处理后花青素含量增加,但基因表达水平没有变化[17]。 由此可见,花青素的生物合成存在复杂的调控机制。
三华李属呼吸跃变型果实,后熟过程中花青素快速积累,但目前尚未有针对三华李花青素合成基因的系统研究。 虽然三月李的基因组测序工作已由国内课题组完成,为李属果树的分子生物学研究奠定了基础[18],但仍未有针对三华李ANS基因的相关研究。 本研究从三华李果实中克隆获得ANS基因,对其序列进行生物信息学分析,并研究其在三华李不同组织和果实后熟过程中的表达模式,为进一步探究三华李花青素合成途径及其调控机理奠定基础。
1 材料与方法
1.1 试验材料
供试三华李植株种植于广西贺州市贺街镇三华李驿站果园。 2021 年6 月1 日(花后约130天)采集七八成熟的果实及茎、叶。 果实采后立即运回实验室,挑选大小均一、无病害及机械损伤的果实,分为两组,分别作为对照组和乙烯处理组,分别用自来水和5 mL/L 乙烯利溶液浸泡1 min,捞出晾干后自封袋密封24 h,之后打开自封口通风透气,于25℃下贮藏;每两天取样一次,放入-80℃超低温冰箱保存。 茎、叶及果皮和果肉各组织用液氮速冻,-80℃保存待用。
1.2 试验方法
1.2.1 RNA 的提取与cDNA 合成 利用华越洋植物RNA 提取试剂盒提取三华李不同组织和不同后熟阶段的总RNA,具体操作方法依照说明书进行。 得到总RNA 后通过超微量分光光度计测定其浓度。 以提取的总RNA 为模板,利用反转录试剂盒进行反转录合成cDNA,合成方法参照试剂盒说明书,保存于-20℃冰箱中,用于后续实验。
1.2.2PsANS基因的克隆 从三华李转录组数据库中获得ANS 序列信息,利用Primer Premier 5.0软件设计特异性引物,引物序列见表1。 以三华李不同后熟阶段的cDNA 为模板,克隆PsANS基因的CDS 序列,进行PCR 扩增。 PCR 反应程序为:94℃预变性3 min;95℃变性15 s,55℃退火30 s,72℃延伸2 min,35 个循环;72℃延伸10 min,于4℃保存。 PCR 产物经过回收、连接并转化至DH5α 感受态细胞,鉴定阳性克隆后测序。 引物合成和基因测序全部交由上海生物科技有限公司完成。

表1 三华李ANS 基因克隆和表达所用引物
1.2.3PsANS基因生物信息学分析 利用NCBI中的BLASTN、BLASTP 进行基因和蛋白序列比对;通过Expasy-ProtParam Tool 在线软件对蛋白的基本理化性质进行预测,初步分析目的基因的功能;亲疏水性利用ProtScale 以Hphob./Kyte &Doolittle 算法进行预测;采用TMHMM 2.0 进行跨膜结构预测;采用SignalP-5.0 预测蛋白质有无信号肽;采用ProtComp Version 9.0 进行蛋白亚细胞定位预测;采用NCBI 的Conserved Domains 分析功能结构域;使用SMART 在线数据库分析蛋白结构域;蛋白质的二级结构采用ExPaSy-SOPMA在线软件进行分析;蛋白质三级结构采用SWISSMODEL 进行在线预测。 利用在线软件STRING预测ANS 与其他蛋白之间的互作情况(以拟南芥蛋白数据库为参考);蛋白序列和同源比对分别使用DNAMAN 和ClustaX 软件。 采用MEGA 5 软件的Neighbor-Joining 法构建系统进化树。
1.2.4PsANS表达分析 根据获得的PsANS基因cDNA 序列设计荧光定量PCR 特异引物(表1)。 以三华李茎、叶、果皮、果肉不同组织以及三华李后熟过程中的样品cDNA 为模板,参考TaKa-Ra 公司的SYBR Green qPCR Master Mix 说明书进行荧光定量PCR 扩增。 试验设置3 个生物学重复,用2-ΔΔCt方法计算PsANS的相对表达量,采用SPSS 26 软件进行LSD 方差分析。
2 结果与分析
2.1 三华李ANS 基因的克隆
三华李果实二代有参转录组参考物种为三月李(Prunus salicina),参考基因组版本为GCA_014863905.1_SCAU_Psal_1.0。 在三华李果实二代有参转录组数据库中筛选初步注释为ANS基因的转录本,其在转录组中的编号为evm.TU.UTG5995.3;在NCBI 中的比对结果表明,该片段与其他物种ANS基因的蛋白序列同源性在80%以上,其中与日本裸樱(Prunus yedoensisvar.nudiflora, PQQ12998) 和 欧 洲 李(Prunus domestica,AHZ30597)ANS的蛋白序列同源性最高。 参考转录组中该基因序列设计引物,以三华李的cDNA为模板进行特异性PCR 扩增,获得ANS基因的CDS 序列(图1);将获得的目的片段与pMD18-T载体连接并转化DH5α 大肠杆菌,挑选阳性克隆测序。 经测序后发现该片段长度为1 074 bp,可编码357 个氨基酸,将其命名为PsANS,GenBank登录号为OP131916。 将该cDNA 片段与三月李基因组序列进行比对,发现该转录本对应的DNA序列在510~726 bp 间有1 个长度为217 bp 的内含子,其基因结构如图2 所示。

图1 三华李ANS 基因cDNA 片段琼脂糖凝胶电泳图谱

图2 三华李ANS 基因DNA 结构图
2.2 PsANS 蛋白理化性质分析
通过在线软件Expasy-ProtParam Tool 分析,该蛋白质的分子式为C1901H3048N508O566S19,分子量为40 401.37,总原子数为5 717。 组成PsANS 蛋白的氨基酸中占比最多的是谷氨酸(Glu),达10.9%,其次为亮氨酸(Leu),占比10.1%。 PsANS蛋白的理论等电点为5.46,不稳定系数为47.96,属于不稳定蛋白。
2.3 PsANS 蛋白亲疏水性分析
预测发现(图3),PsANS 的氨基端和羧基端表现出疏水性,亲水性大部分集中在中心区域;疏水出现在第191 位的氨基酸残基,亲水出现在第65 位氨基酸残基。 PsANS 蛋白的亲水性总平均值(GRAVY)为-0.371,预测该蛋白属于亲水性蛋白。 PsANS 蛋白不存在跨膜区,无信号肽,为非分泌蛋白。 通过ProtComp Version 9.0 在线软件预测PsANS 蛋白在植物细胞中可能定位在细胞质。

图3 PsANS 蛋白疏水/亲水性预测
2.4 PsANS 蛋白二级和三级结构预测
在线软件ExPaSy-SOPMA 预测PsANS 蛋白的二级结构,结果(图4)表明该蛋白二级结构中α-螺旋(h)占35.29%,β-折叠(t)占5.32%,无规则卷曲(c)占41.18%,延伸链(e)占18.21%。 利用SWISS-MODEL 以拟南芥ANS 蛋白为模板构建PsANS 蛋白的三维结构模型(图5)。 以拟南芥蛋白数据库为参考,通过STRING 预测PsANS 潜在的互作关系,结果(图6)表明,ANS 蛋白主要与TT8(LDOX,无色花青素双加氧酶)、TT4(CHS,查尔酮合成酶)、DFR(二氢黄酮醇-4-还原酶)、UF3GT(类黄酮3-O-葡萄糖基转移酶)等存在互作关系。

图4 PsANS 蛋白二级结构预测

图5 PsANS 编码蛋白三级结构模型
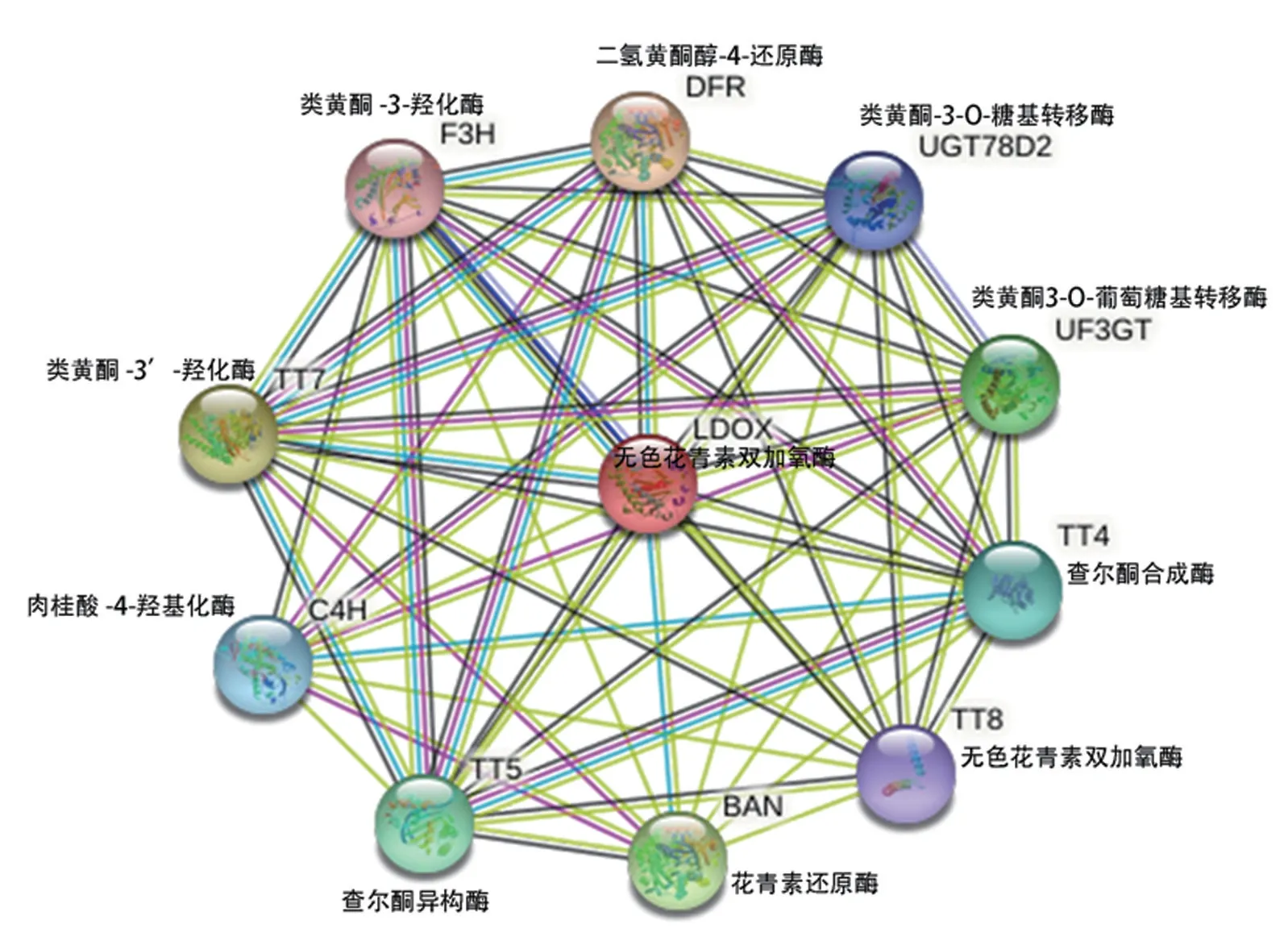
图6 ANS 蛋白互作网络模型预测
2.5 蛋白序列保守结构域、同源比对和系统进化分析
利用NCBI-CDD 对PsANS 蛋白的结构域进行分析,显示PsANS 为PLN03178(花青素合成酶)多域蛋白,有1 个保守结构域,属于2-酮戊二酸-Fe2+-双加氧酶家族[2OG-Fe(Ⅱ)-Oxy,第216 ~311位];该保守结构域含有与2-酮戊二酸特异性结合的精氨酸Arg(R293,302)和丝氨酸Ser(S240,270,283,289,304)位点7 个,与Fe2+结合具有双加氧功能的组氨酸His(H236,238,247,274,292)和天冬氨酸Asp(D277)位点6个,这些位点在不同物种的ANS 序列中高度保守。通过与其他12 种植物的ANS 蛋白进行同源性多重比对发现,ANS 氨基酸序列较为保守(图7),PsANS 与同为蔷薇科的欧洲李更近源(图8)

图7 PsANS 与其他已知ANS 蛋白的同源性比较

图8 三华李与其他植物ANS 蛋白系统进化树
2.6 PsANS 在三华李不同组织中的表达分析
植物次生代谢合成相关基因往往具有特异性的时空表达模式,即在代谢旺盛的组织中,相关基因的表达水平较高[10]。 由图9 可见,PsANS在三华李的4 个不同组织中均有表达,其表达量在三华李果肉中最高,其余依次是果皮、叶、茎,各组织间存在显著的表达差异。
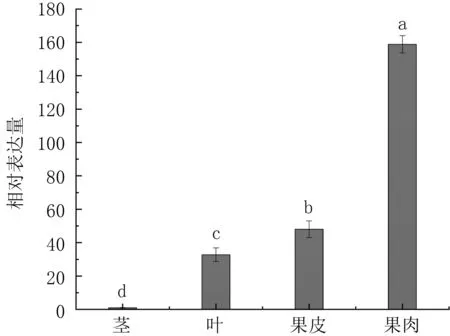
图9 PsANS 在三华李不同组织中的表达
2.7 PsANS 在三华李果实后熟过程中的表达分析
利用荧光定量PCR 技术分析PsANS在三华李后熟过程中的表达变化,结果(图10)显示,对照组和乙烯处理组PsANS表达量在后熟过程中都增加,贮藏6 d 的表达量分别是0 d 的3.6 倍和6.2 倍。 与对照组相比,乙烯处理组PsANS表达显著上调(P<0.05)。 贮藏4 d 时,乙烯处理组的PsANS达到表达高峰,是0 d 的8.9 倍。
3 讨论与结论
三华李因富含花青素等抗氧化活性物质深受广大消费者喜爱和科研工作者关注[7]。 研究三华李果实花青素合成关键基因可为培育具有优良外观品质的李子品种提供理论依据。 花青素合成酶(ANS)是花青素合成通路末端的酶,参与花青素的合成和累积,在果实着色及花色形成中具有重要作用[12]。
本研究首次从三华李果实中克隆获得ANS基因,并将其命名为PsANS,该序列的编码区长度为1 074 bp,可编码一个由357 个氨基酸组成的蛋白质;与同属蔷薇科的苹果梨[14]ANS基因长度一致。 本研究发现ANS 的氨基酸序列在不同植物中具有较高的保守性,都具有典型的2OG-Fe(Ⅱ)-Oxy 家族保守结构域和花青素合成酶催化位点Arg、Ser、His、Asp。 PsANS 与其他植物ANS蛋白性质相似,为无跨膜区、无信号肽的不稳定亲水性蛋白,是一种非分泌性蛋白[19]。 花青素主要贮藏于植物细胞的液泡中,但其合成主要在细胞质的内质网表面进行,需经各种修饰后才被运至液泡等部位储存[20],因此,包括ANS 在内的一系列酶集中形成多酶复合体在细胞质中催化花青素的合成[21]。 本研究预测发现PsANS 也定位于细胞质中。
以往研究发现ANS基因表达具有明显的组织特异性,在植物体的各个组织中均有表达,但表达量存在差异[12,22,23]。 本研究发现,PsANS基因在三华李不同组织中均有表达,果肉中的表达量最高,茎中最低,暗示PsANS在三华李果肉花青素合成过程中扮演重要角色。 花青素含量是判断三华李果实成熟度的显著标志,在成熟过程中快速上升。 三华李是呼吸跃变型果实,后熟过程中果肉的花青素快速合成,而外源乙烯处理能促进果实快速软化和转红。 以往研究表明外源乙烯对果实花青素合成有显著影响,例如乙烯处理可上调桑椹ANS的表达并促进果实花青素合成[24];但红梨和桃果实的花青素合成受乙烯处理显著抑制[25,26]。 可见,乙烯对不同植物花青素合成的影响较为复杂。 本研究发现,三华李果实后熟过程中PsANS表达呈上升趋势,外源乙烯处理进一步促进其表达,表明PsANS响应乙烯信号,在李果实后熟转色过程中起重要作用。 但乙烯信号途径和花青素合成途径中转录因子间的作用机制还需进一步研究。
本研究首次克隆得到三华李果实花青素合成酶基因PsANS全长cDNA,该序列长度为1 074 bp,编码357 个氨基酸。 PsANS 蛋白具有花青素合成酶的典型结构特征,氨基酸序列与日本裸樱和欧洲李的ANS 氨基酸序列相似度高;三华李花青素合成酶基因的表达存在组织器官差异性,果肉中表达量最高,茎中最低;三华李后熟转色过程中PsANS表达呈上升趋势,乙烯处理进一步上调了PsANS的表达,表明其对三华李果实发育和后熟过程中花青素的合成起着重要作用。
