间伐对杉木人工林土壤微生物残体碳的影响*
2023-08-09崔朝伟彭丽鸿马东旭王佳琪江祥庆江先桂马祥庆林开敏
崔朝伟 彭丽鸿 马东旭 王佳琪 江祥庆 江先桂 马祥庆 林开敏
(1. 福建农林大学林学院 福州 350002;2. 国家林业和草原局杉木工程技术研究中心 福州 350002;3. 福建省沙县官庄国有林场 三明 350803)
土壤有机碳(soil organic carbon,SOC)是陆地生态系统中最大的碳库,其碳储量为植被碳库的3~4 倍、大气碳库的2~3 倍(Lehmannet al., 2015;于颖超等,2022),在全球碳循环中发挥着重要作用。植物、动物和微生物来源的有机碳通过土壤生物新陈代谢和有机碳淋溶输入或损失等方式影响SOC 固存(Nottinghamet al., 2015;Mouet al., 2021),长期以来,受研究手段和分析技术限制,有关报道多关注木质素、纤维素和半纤维素等植物碳源对稳定SOC 库的贡献(Zhuet al.,2003;Hallet al., 2020;Lianget al., 2017);然而,目前关于SOC 库形成与稳定机制的认知已逐渐从早期的腐殖质理论转化为关注微生物转化和调控SOC 库的新共识(Kallenbachet al., 2015;Schmidtet al., 2011;梁超等, 2021)。土壤微生物通过同化作用经“体内周转”将易分解有机物转化为微生物生物量和自身代谢产物(梁超等, 2021),微生物死亡残体及部分代谢产物与土壤团聚体结合或与有机质结合形成较难被微生物分解利用的有机-无机复合体(Köegel-Knabneret al.,2008; Lehmannet al., 2007), 进入非生物SOC 库(Cotrufoet al., 2019;Lavalleeet al., 2020),即微生物主要以贡献者角色调控SOC 库中微生物来源碳的动态。Hu 等(2022)研究发现,土壤pH、Ca2+含量可通过调节微生物群落影响土壤微生物残体碳含量。Chen 等(2017)分析得出,土壤酶介导的分解过程是控制全球养分循环的关键步骤。Liang 等(2019)基于META 分析估算温带农田、草地和森林生态系统表层土壤中微生物残体占SOC 的比例,分别为农田56%、草地62%和森林33%。Fan 等(2021)利用模型估算全球不同生态系统土壤中微生物残体碳占总SOC 的比例为10%~27%。可见,因SOC 库来源、周转时间和稳定机制不同,微生物残体在不同生境中对SOC 库的贡献存在不确定性。
生物标志物氨基糖是微生物细胞壁的重要组成部分,微生物死亡后会在土壤中存留很长时间,通过对氨基糖的计算能够得到微生物残体碳含量(Maet al., 2018;Shaoet al., 2017)。目前,已有4 种氨基糖可被标准化提取并量化分析(Zhanget al., 1996),包括氨基葡萄糖(glucosamine, GluN)、氨基半乳糖(galactosamine,GalN)、氨基甘露糖(mannosamine,ManN)和胞壁酸(muramic acid,MurA)。氨基葡萄糖是真菌几丁质的唯一成分和脱酰基几丁质的主要成分,也有一小部分来源于细菌肽聚糖;胞壁酸唯一来源于细菌,是细菌中脂多糖和细胞壁肽聚糖的主要成分(Lianget al.,2013);氨基半乳糖来源尚不明确,一般认为来源于细菌;氨基甘露糖来源不明确且含量极低,在文献报道中较少(Amelunget al., 2008)。基于土壤氨基糖的异源性可区分细菌或真菌对土壤有机碳积累的相对贡献(Joergensen, 2018),同时研究中常用氨基葡萄糖与胞壁酸的比值反映土壤细菌、真菌残体碳对土壤SOC 转化的相对贡献(Heet al., 2011;Iyyemperumalet al., 2008)。
杉木(Cunninghamia lanceolata)是我国南方特有的速生用材树种,具有生长迅速、材质优良、产量高、抗逆性强等特性(Wanget al., 2010)。第九次全国森林资源清查显示,我国现有杉木人工林面积达0.099亿hm2、蓄积量达7.55 亿m3,分占全国人工乔木林总面积、总蓄积量的1/4 和1/3。但杉木人工林因过度追求速生丰产、纯林化及多代连栽,导致地力衰退问题愈加突出,重要原因之一是SOC 库逐渐减少,直接制约和威胁当前杉木人工林的可持续发展,如何合理经营来维持和恢复杉木人工林SOC 库,成为缓解地力衰退的一个重要课题。间伐是杉木人工林经营采取的重要措施,其能否调控土壤微生物生长、代谢和微生物残体形成并向稳定有机质转化,从而驱动土壤微生物残体碳形成及SOC 积累的机理尚不清楚。鉴于此,本研究以福建省沙县官庄国有林场不同间伐强度杉木人工林为对象,以氨基糖为土壤微生物残体的组分标识物,系统探究杉木人工林土壤微生物残体对间伐的响应及对SOC 库的直接贡献,并深入探讨微生物残体碳与土壤理化性质、酶活性及SOC 库等指标的内在相互关系,寻找影响土壤微生物残体碳的关键驱动因子,揭示杉木人工林SOC 库形成和积累的调控机制,以期为杉木人工林可持续经营、缓解全球气候变化和实现我国碳中和目标提供科学依据。
1 研究区概况
研究区位于福建省沙县官庄国有林场所属的池村(117°43′15—117°43.18′E,26°32′61—26°32′67N),地处武夷山脉与戴云山脉之间,属中亚热带季风气候,平均海拔150~350 m,年均气温18 ℃,年均降水量1 700 mm,土壤类型以岩浆岩、沉积岩为主,土壤以黄红壤为主,腐殖质含量较高,立地指数18 左右。林下植被主要有杜茎山(Maesa japonica)、紫麻(Oreocnide frutescens)、粗叶榕(Ficus hirta)、紫珠 (Callicarpa bodinieri)、傅氏凤尾蕨(Pteris fauriei)、枸骨(Ilex cornuta)、华南毛蕨(Cyclosorus parasiticus)、中华薹草(Carex chinensis)等。
2 研究方法
2.1 样地设置
研究区试验林间伐前为8 年生杉木人工林,2009年造林时初植密度为3 250 株·hm-2,2017 年底基于大径材培育原则设计实施间伐(伐后移除间伐及其残留物,如枝条等,以消除因间伐残留物引起的试验结果差异)。设置弱度(weak thinning,WT;保留株数2 250 株·hm-2)、中度(moderate thinning,MT;保留株数1 800 株·hm-2)和强度(heavy thinning,HT;保留株数1 200 株·hm-2)3 种间伐强度,每处理3 次重复,共9 块样地(20 m×20 m),样地间距大于10 m,其概况见表1。
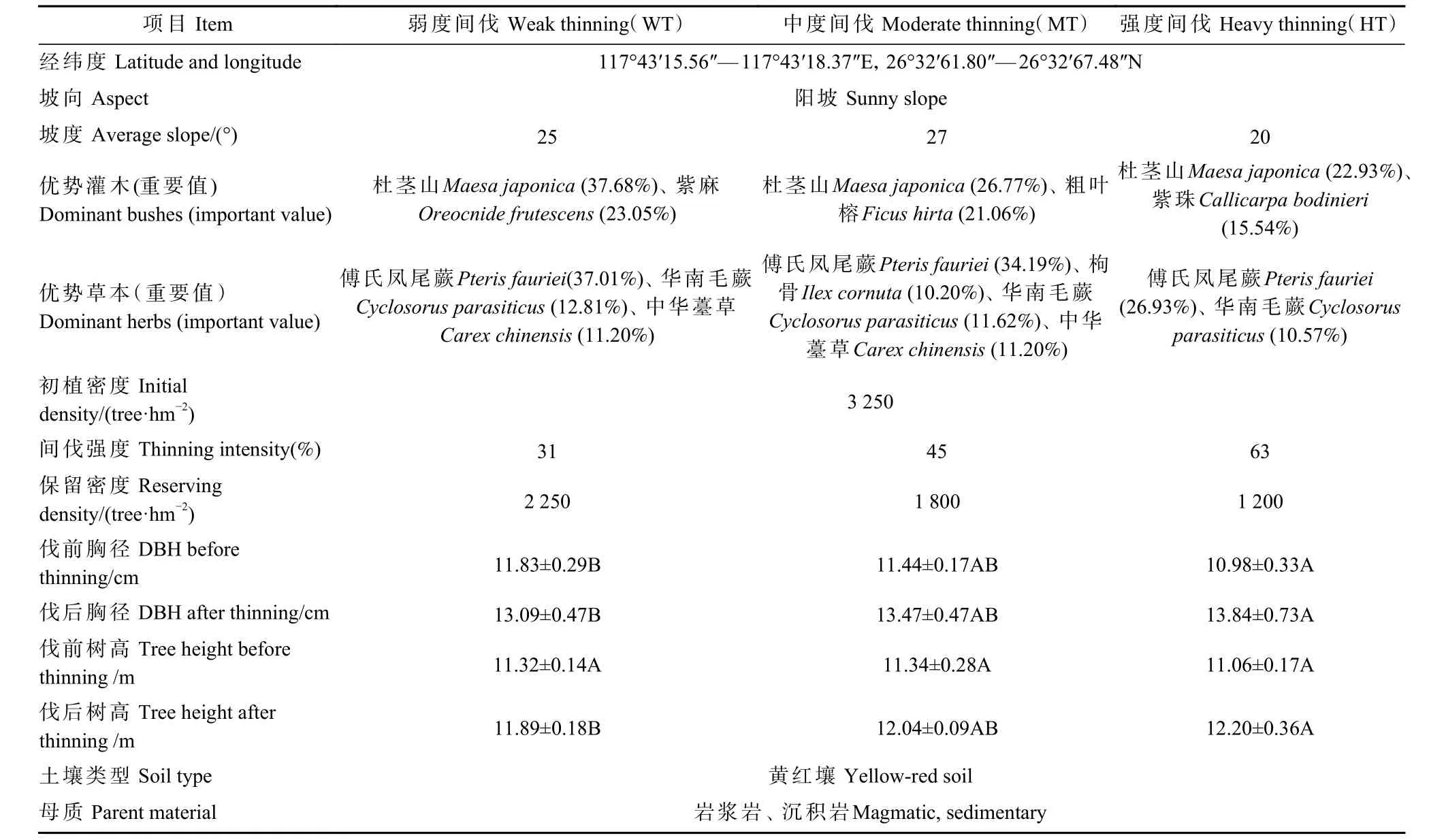
表1 林分样地概况①Tab. 1 Profile of stand sample plots
2.2 土壤样品采集
2021 年11 月,在每块样地内按“S”形设置5 个土壤取样点,分为0~10、10~20 cm 土层,每层分别取样后充分混合,采集1 kg 左右土样,储存在自封袋中带回实验室。土样一部分置于4 ℃冰箱内保存,用于微生物生物量碳氮测定;另一部分在实验室内自然风干后过2 mm 和0.149 mm 筛,用于土壤化学性质分析和氨基糖测定。
2.3 土壤样品分析
土壤密度(soil dendity,SD)和自然含水率(soil moisture content,SMC)采用环刀法测定;土壤pH(土水比1∶2.5)用pH 计测定;土壤有机碳(SOC)、全氮(total nitrogen,TN)含量用元素分析仪(Elementar,Vario Max CN, Germany)测定; 土壤全磷(total phosphorus,TP)含量用ICP 测定;有效磷(available phosphorus,AP)含量采用酸溶-钼锑抗比色法测定;土壤微生物生物量碳(microbial biomass carbon,MBC)含量通过氯仿熏蒸法、K2SO4浸提后用总有机碳分析仪(岛津TOC-V CPH)测定;可溶性碳(dissolved organic carbon,DOC)含量通过K2SO4浸提后用总有机碳分析仪测定;硝态氮(NO3--N)、铵态氮(NH4+-N)含量通过KCl 溶液浸提后用流动分析仪(Skala San++,荷兰)测定;酸性磷酸酶(soil acid phosphatase,SACP)活性采用磷酸苯二钠法测定;多酚氧化酶(soil polyphenol oxidase,SPPO)活性采用没食子素比色法测定;过氧化物酶(soil peroxidase,SPOD)活性采用邻苯三酚比色法测定;β-葡萄糖苷酶(β glucosidase,β)活性采用硝基苯-β-D-吡喃糖苷(nitrobenzene-β-D-pyranoside,PNPG)比色法测定。
氨基糖测定方法(Indorfet al., 2011):称取0.3 mg氮的土样置于水解瓶中,加入10 mL 6 mol·L-1盐酸,水解8 h。冷却后加入100 µL 内标1(肌醇),转移至旋转蒸发仪进行干燥,残留物用纯水溶解并用稀KOH 和HCl 将pH 调至6.6~6.8,转移至离心管以3 000 r·min-1离心10 min 去除沉淀。上清液用冷冻干燥仪冻干,残留固体物质用4 mL 无水甲醇溶解,再次以3 000 r·min-1离心10 min 达到除盐目的。将上清液转移至5 mL 衍生瓶中,45 ℃下用N2吹干,加入1 mL水,同时加入100 µg N-甲基氨基葡萄糖(内标2),摇匀后再次进行冷冻干燥。向干燥后的样品中加入300 µL 衍生试剂,加盖密封,75~80 ℃水浴加热30 min,其间振荡3~4 次使反应均匀。冷却至室温,加入1 mL乙酸酐,密封,水浴加热20 min。冷却后,加入1.5 mL二氯甲烷,涡旋使溶液混合均匀。去除过量衍生试剂的样品45 ℃下用N2吹干,溶于400 µL 乙酸乙酯-正己烷(容积比为1∶1)中,采用气相色谱质谱联用仪(Agilent 7890A-5975C,USA)对产物进行分离和检测。记录样品和标准品的保留时间,通过比较判断氨基糖衍生物的峰值,将纯化前向样品中加入的肌醇作为内标对氨基糖进行定量分析。
土壤真菌残体碳(fungal microbial residue carbon,MRCF)含量(g·kg-1)计算公式为:
土壤细菌残体碳(bacterial microbial residue carbon MRCB)含量(g·kg-1)计算公式为:
式中:氨基葡萄糖(GluN)、胞壁酸(MurA)的分子质量分别为179.2、251.2 g·mol-1;GluN、MurN 的单位为mmol·g-1。
微生物总残体碳(microbial residue carbon,MRC)含量(g·kg-1)为MRCF与MRCB含量加和(Danget al.,2018),微生物残体对有机碳的贡献为微生物残体碳含量与有机碳含量的比值。
2.4 数据统计分析
运用SPSS 22.0 软件进行单因素方差分析(oneway ANOVA)、多重比较(Duncan)和双因素方差分析(two-way ANOVA),Origin pro 软件绘图。结构方程模型(structural equation modeling,SEM)在AMOS 22.0软件中完成,利用SEM 分析土壤环境因子对MRC 的影响路径和程度,采用极大似然估计法对模型进行拟合。数据为平均值±标准差(n=3)。
3 结果与分析
3.1 不同间伐强度下土壤微生物残体碳含量差异
由图1 可知,在0~10 和10~20 cm 土层,土壤MRCB、MRCF、MRC 均随间伐强度增加而升高,且强度间伐显著高于弱度间伐(P<0.05);MRCF/MRCB在0~10 cm 土层表现为弱度间伐显著高于中度间伐和强度间伐(P<0.05),在10~20 cm 土层不同间伐处理间无显著差异。随土层深度增加,不同间伐强度下土壤MRCB、MRCF、MRC 均显著降低(P<0.05),变化范围分别为1.50~2.80、3.22~6.00、4.72~8.69 g·kg-1。经方差分析验证,间伐强度和土层深度对土壤MRCB、MRCF、MRC、MRCF/MRCB均有显著影响(P<0.05)。
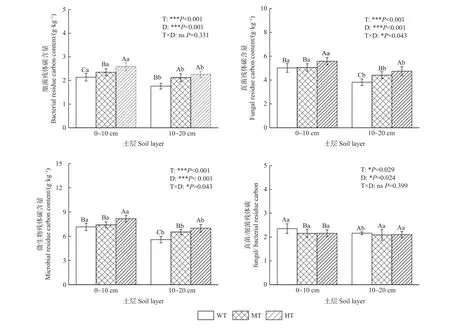
图1 不同间伐强度下微生物残体碳含量垂直分布Fig. 1 Vertical distribution of microbial carbon content under different thinning intensities
3.2 不同间伐强度下微生物残体碳对SOC 的贡献差异
由图2 可知,不同间伐强度下土壤MRCB、MRCF、MRC 对SOC 的贡献在0~10 cm 土层无显著差异,在10~20 cm 土层土壤MRC 先升后降,其中,中度间伐显著高于强度间伐(P<0.05)。随土层深度增加,土壤MRCB、MRCF、MRC 对SOC 的贡献随之增加,变化范围分别为13.20%~18.99%、28.42%~39.72%、41.62%~58.70%,中度间伐下土壤MRCB、MRCF、MRC 对SOC的贡献均为0~10 cm 土层显著高于10~20 cm 土层(P<0.05)。
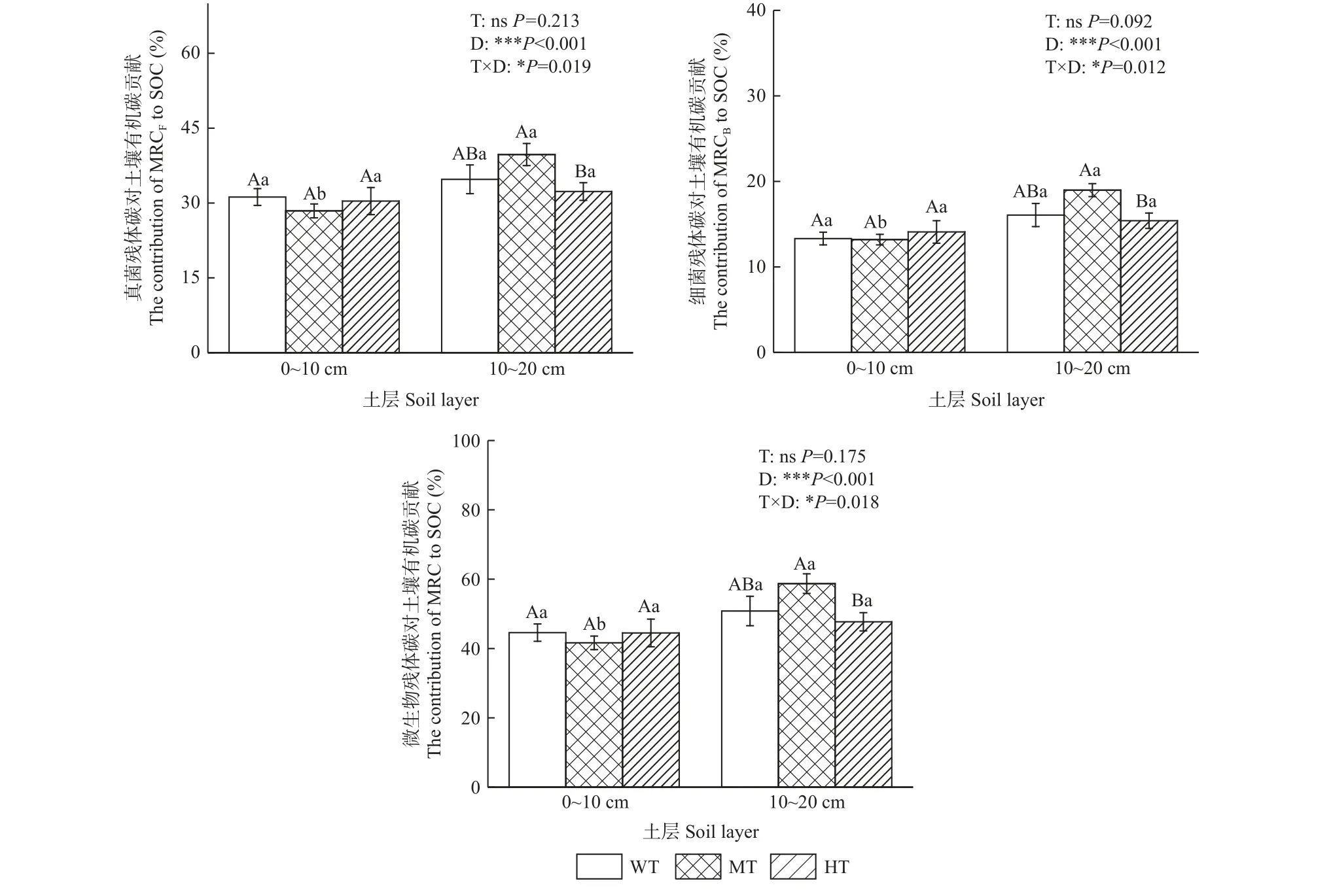
图2 不同间伐强度下微生物残体碳对土壤有机碳贡献的剖面分布Fig. 2 Profile distribution of microbial necromass carbon contribution to SOC under different thinning intensities
3.3 不同间伐强度下土壤理化性质特征差异
由表2 可知,在0~10 和10~20 cm 土层,不同间伐强度下土壤SOC、TP、AP、MBC、DOC、NO3--N、NH4+-N 含量均为强度间伐>中度间伐>弱度间伐(P<0.05);土壤SMC、TN 含量随间伐强度增加先降后升;SD、pH 随间伐强度增加而降低。随土层深度增加,土壤SMC、SOC、TN、TP、AP、MBC、DOC 含量均呈下降趋势,pH、SD、NO3--N、NH4+-N 含量与之相反。经方差分析验证,间伐强度(T)和土层深度(D)对SOC、TN、TP、AP、MBC、NO3--N、NH4+-N 含量均有显著或极显著影响(P<0.05)。
3.4 不同间伐强度下土壤酶活性差异
由图3 可知,在0~10 和10~20 cm 土层,土壤酸性磷酸酶、过氧化物酶和β-葡萄糖苷酶含量均随间伐强度增加而升高,且强度间伐显著高于弱度间伐(P<0.05);土壤多酚氧化酶活性在0~10 cm 土层表现为弱度间伐和强度间伐显著高于中度间伐(P<0.05),在10~20 cm 土层不同间伐处理间无显著差异。随土层深度增加,土壤酸性磷酸酶、多酚氧化酶、过氧化物酶和β-葡萄糖苷酶活性均呈不同程度下降趋势。经方差分析验证,间伐强度和土层深度对土壤酶活性有显著或极显著影响(P<0.05)。
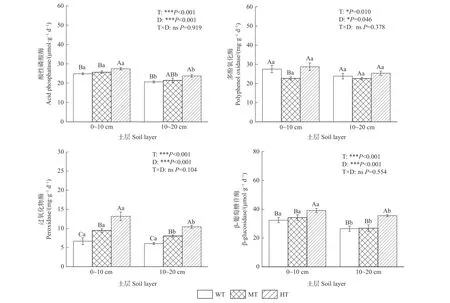
图3 不同间伐强度下土壤酶活性的垂直分布Fig. 3 Vertical distribution of soil enzyme activities under different thinning intensities
3.5 不同间伐强度下土壤微生物残体碳含量变化的驱动因子
SEM 模型a参数为χ2/df=0.992、P=0.506、CFI=1.000、IFI=1.002、RMSEA=0.000,模型拟合良好。由图4a 可知,在0~10 cm 土层,土壤物理性质与酶活性、微生物残体碳含量间均有正影响,路径系数分别为0.09、0.17;与化学性质间有负影响,路径系数为-0.60。土壤化学性质与酶活性、微生物残体碳含量和碳形态间均有正影响,路径系数分别为1.13、0.98 和1.49。酶活性与微生物残体碳含量、碳形态间均有负影响,路径系数分别为-0.02、-0.23。微生物残体碳含量与碳形态间有负影响,路径系数为-0.27。根据路径系数可知,化学性质是对酶活性、碳形态和微生物残体碳含量影响较大的潜变量。对0~10 cm 土层土壤理化性质、酶活性与微生物残体碳含量进行冗余分析(图5a)发现,土壤可溶性碳(P=0.002)和铵态氮(P=0.066)是对土壤微生物残体碳含量影响较大的土壤单因子。
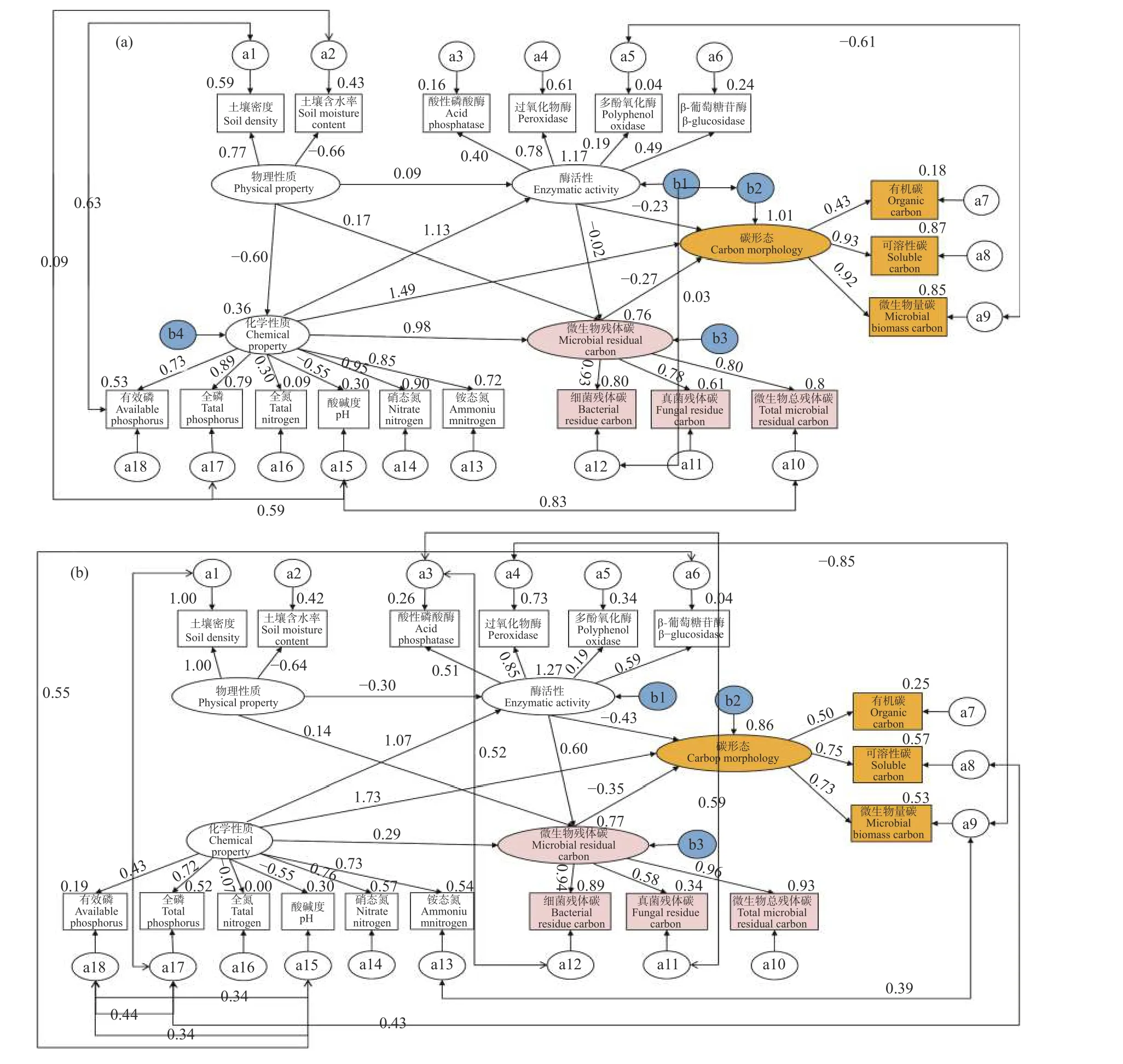
图4 微生物残体碳影响因素的结构方程模型分析Fig. 4 Structural equation model analyses of the factors affecting microbial residue carbon contents
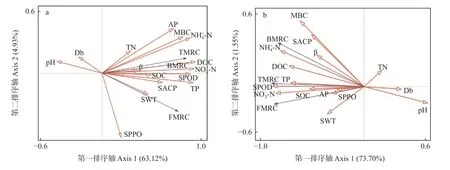
图5 土壤理化性质、酶活性与微生物残体碳的冗余分析Fig. 5 Redundancy analysis of soil physicochemical properties, enzyme activities and microbial carbon residues
SEM 模型b参数为χ2/df=1.167、P=0.105、CFI=0.938、IFI=0.945、RMSEA=0.080,模型拟合良好。在构建和修正模型b时发现,土壤物理性质与化学性质间关联度较低,且影响模型适配度,故删除二者间通路。由图4b 可知,在10~20 cm 土层,土壤物理性质与酶活性间有负影响,路径系数为-0.30;与微生物残体碳含量间有正影响,路径系数为0.14。化学性质与酶活性、微生物残体碳含量和碳形态间均有正影响,路径系数分别为1.07、0.29 和1.73。酶活性与微生物残体碳含量间有正影响,路径系数为0.60;与碳形态间有负影响,路径系数为-0.43。微生物残体碳含量与碳形态间有负影响,路径系数为-0.35。根据路径系数可知,化学性质是对土壤酶活性、碳形态影响较大的潜变量,酶活性是对微生物残体碳含量影响较大的潜变量。对10~20 cm 土层土壤理化性质、酶活性与微生物残体碳含量进行冗余分析(图5b)发现,土壤过氧化物酶活性(P=0.002)和硝态氮含量(P=0.034)是对土壤微生物残体碳含量影响较大的土壤单因子。
4 讨论
4.1 不同间伐强度下土壤微生物残体碳含量差异
抚育间伐刺激土壤MRC 积累,不同间伐强度下MRCB、MRCF、MRC 含量存在差异。与弱度间伐相比,强度间伐下0~10 和10~20 cm 土层土壤MRCB、MRCF、MRC 分别增长12.0%、18.9%、14.0%和27.84%、24.32%、25.44%,这是因为间伐强度增加导致林分郁闭度下降,林内光照条件改善,利于林下植被生长发育,其凋落物多样性及根部沉积新碳源刺激微生物生长的资源分配,从而促进容纳更多的微生物群落(Hogberget al.,2006;Dinget al., 2013)。同样MRCF/MRCB在不同间伐强度间也存在差异,如0~10 cm 土层弱度间伐的MRCF/MRCB显著高于中度间伐和强度间伐(P<0.05),10~20 cm 土层不同间伐强度间无显著差异(图1)。对于MRCB而言,由于弱度间伐下杉木人工林林木保留密度高、植被多样性低、凋落物数量多且难分解,而细菌主要分解易降解有机质(Groveret al., 2015),故细菌数量占总微生物数量降低,导致弱度间伐下0~10 cm 土层MRCF/MRCB相对较高;而在10~20 cm 土层植物碳源输入随土层深度增加而降低,且随间伐强度增加,土壤MRCB分别下降21.3%、10.9%和14.8%,MRCF分别下降30.6%、14.7%和17.6%,相较中度间伐和强度间伐,弱度间伐下MRCF较MRCB下降幅度更大,故在10~20 cm 土层不同间伐强度间无显著差异。不同土层深度下,土壤MRCB、MRCF、MRC、MRCF/MRCB表现为0~10 cm 土层高于10~20 cm 土层,与Fan 等(2020)、Sradnick 等(2014)研究结果一致。Ni等(2020)基于META 分析发现,随土层深度(地平线、0~20 cm、20~50 cm、50~100 cm)增加,土壤氨基葡萄糖(GluN)、氨基半乳糖(GalN)、胞壁酸(MurN)含量均显著降低(P<0.05),这是因为凋落物及根系残留物等植物碳源进入底层土壤前,土壤微生物可吸收大量碳源和有效养分,导致表层土壤含有较高的微生物生物量和较快的微生物周转效率,从而使土壤MRC 被持续固存在表层土壤中(Spohnet al., 2016)。
4.2 不同间伐强度下土壤微生物残体碳对SOC 的贡献差异
MRC 是土壤有机碳的重要来源。本研究MRC含量占土壤有机碳含量的41.62%~58.70%,其中MRCF的贡献(28.42%~39.72%)显著高于MRCB(13.20%~18.99%),该差异可能与细菌、真菌及自身稳定性和周转速率有关。细菌细胞壁中富含营养的化合物(肽聚糖)可快速分解,在碳源或养分缺乏情况下,其会作为土壤微生物主要碳源或氮源被分解利用(Huet al.,2019);真菌细胞壁中几丁质等顽固性结构化合物稳定性较高,且黑色素的保护使其分解性远低于其他类聚合物,因此真菌残体的降解速度比细菌慢(Fernandezet al., 2016)。
不同间伐强度下,土壤MRCB、MRCF、MRC 在0~10 cm 土层对SOC 的贡献无显著差异,在10~20 cm土层表现为中度间伐显著高于强度间伐(P<0.05),这归因于强度间伐下杉木人工林林下植被物种多样性和丰富度较高,导致植被凋落物及根系沉积物数量较多且大部分为易分解碳源,故强度间伐下植物残体来源碳占有机碳库比例升高,MRC 对SOC 的贡献低于弱度间伐和中度间伐。Prommer 等(2020)研究表明,更高的植物多样性会增加微生物生长、生物量和周转速率,从而推动土壤有机碳积累,但随植物多样性增加,MRC 对SOC 的贡献反而呈下降趋势。随土层深度增加,不同间伐强度下土壤MRCB、MRCF、MRC 对SOC 的贡献呈升高趋势,相较0~10 cm 土层,10~20 cm土层土壤MRCB、MRCF、MRC 分别增加9.28%~43.85%、6.29%~39.74%、7.24%~41.04%,与Ni 等(2020)、于颖超等(2022)研究结果一致。
4.3 不同间伐强度下土壤理化性质及酶活性差异
在0~10 和10~20 cm 土层,土壤SOC、TP、AP、MBC、DOC、NO3--N、NH4+-N 含量表现为强度间伐>中度间伐>弱度间伐,土壤pH、密度随间伐强度增加而降低,这是因为间伐强度较弱的杉木人工林再次郁闭后,林内光照不足,林下植被物种多样性、盖度和生物量受到限制,阻碍凋落物分解,从而影响土壤养分和物理结构状况(Testeet al., 2012)。Dang 等(2018)研究发现,与未间伐样地相比,强度间伐后松木人工林土壤SOC、TN、TP、AP 和NO3--N 含量分别增加28.02%、21.26%、26.92%、37.58%和4.88%(P<0.05),土壤pH 在不同处理间无显著差异。Zhou 等(2016)研究得出,杉木人工林土壤养分含量随间伐强度增加而升高,且间伐后因林下植被快速发育使土壤密度降低。徐雪蕾等(2019)研究指出,与未间伐和较弱间伐处理相比,强度间伐对杉木人工林表层土壤SOC、TN、TP含量有显著正效应(P<0.05)。上述研究结果与本研究较为一致,较强间伐处理改良土壤养分及孔隙状况的能力更明显。
同时,本研究不同间伐强度下土壤酶活性也存在差异,即随间伐强度增加,土壤酸性磷酸酶、过氧化物酶和β-葡萄糖苷酶活性呈升高趋势(P<0.05);土壤多酚氧化酶活性呈先下降后升高趋势,且在0~10 cm 土层弱度间伐和强度间伐显著高于中度间伐(P<0.05)。丁波等(2017)研究发现,强度间伐可显著提高杉木人工林土壤过氧化氢酶、碱性磷酸酶、脲酶和蔗糖酶活性(P<0.05)。也有研究表明,间伐强度增加会促进杉木人工林林下植被盖度和生物量增大,增加根系分泌物种类和凋落物组成多样性,为微生物分泌土壤酶提供底物,进而提高土壤酶活性(郝俊鹏等,2013),与本研究结果不尽相同。于立忠等(2017)研究发现,相比未间伐处理,较强间伐可显著提高肥力较差的日本落叶松(Larix kaempferi)人工林表层酚氧化酶和N-乙酰-β-氨基葡萄糖苷酶(N-acetyl-β-glucosaminidase,NAG)活性,显著降低林分亚表层β-葡萄糖苷酶、NAG 和酸性磷酸酶活性。需要指出的是,间伐如何导致土壤酶活性降低是一个十分复杂的过程,以往研究发现,间伐可改变林内光照、温度和水分条件,调整林分结构,促进林下植被根系活动(Testeet al., 2012;郝俊鹏等,2013),并对土壤微生物数量和活性产生一定影响,而土壤酶是十分敏感的生物活性物质,由间伐处理间接导致的林分条件变化均可能使土壤酶活性发生改变。
4.4 不同间伐强度下土壤微生物残体碳含量变化驱动因子分析
SEM(图4a、图5a)显示,在0~10 cm 土层,土壤化学性质是对微生物残体碳含量影响较大的潜变量。有研究表明,土壤化学性质如pH、Ca2+含量可通过调节微生物群落影响土壤微生物残体碳含量(Huet al.,2022);也有研究表明,在有机碳含量较高的土壤中,微生物残体碳含量也较高(Shaoet al., 2019)。本研究发现,可溶性碳(P=0.002)和铵态氮(P=0.066)是对土壤微生物残体碳含量影响较大的土壤单因子。可溶性碳作为微生物生长的速效底物,可为微生物提供能量从而影响微生物量(Kalbitzet al., 2003)。于颖超等(2022)构建SEM 时发现,土壤DOC 通过影响微生物生物量显著促进MRC 积累,真菌、细菌残体碳含量随土壤可溶性碳、氮、磷含量增加而增加(Wanget al.,2021;李庭宇等,2022)。同样,氮素作为微生物细胞壁几丁质和肽聚糖的重要组成元素,是影响土壤微生物残体碳积累的重要土壤因子(Schmidtet al., 2017)。丁雪丽等(2009)研究发现,在培养前期氨基糖总量随无机氮素提高而增加(P<0.05),且随培养时间延长,氨基糖总量有所下降,但对照组和低施氮组的下降幅度大于高施氮组。Zhang 等(2016)研究表明,当土壤中缺乏养分时,微生物残体碳会作为有效碳源或氮源被微生物活体优先利用;反之,则促进微生物残体碳在土壤中的积累。本研究中,随间伐强度增加,土壤DOC、NH4+-N 等速效底物含量显著提高,导致微生物残体原有分解和积累的平衡被打破,促使微生物残体在表层迅速积累,这与上述研究结果一致。而在10~20 cm 土层,SEM(图4b、图5b)显示,土壤酶活性是对微生物残体碳含量影响较大的潜变量。有研究表明,土壤中酶介导的分解过程是控制全球养分循环的关键步骤(Chenet al., 2017)。本研究中,随间伐强度增加,土壤酸性磷酸酶、过氧化物酶和β-葡萄糖苷酶活性均显著增加(P<0.05),其中酸性磷酸酶可将土壤中的有机磷分解成易利用的速效磷,缓解微生物代谢的磷限制(滕泽栋等,2017),过氧化物酶和β-葡萄糖苷酶分别是木质素等大分子的过氧化物酶和纤维素的水解酶,二者均可为微生物提供可利用的底物和能源(Zhouet al., 2021;邓先智等,2022),从而提高微生物利用碳源的能力。过氧化物酶(P=0.002)和硝态氮(P=0.034)是对土壤微生物残体碳含量影响较大的土壤单因子。综上可知,过氧化物酶可将土壤中大分子(木质素)等物质进行“剪切”,使其转化为可被微生物有效利用的小分子物质,进而提高土壤“微生物碳泵”体内周转速率(Lianget al., 2011),这种高速的细胞周转速率更有利于土壤微生物残体的积累(邓先智等,2022)。
5 结论
基于杉木人工林密度控制试验发现,随间伐密度增加,土壤养分和酶活性大多呈升高趋势;土壤微生物残体碳含量及其对SOC 库的贡献均有不同程度增加,且以真菌残体碳为主。在土壤垂直分布上,随间伐强度增加,微生物残体碳含量显著降低,而微生物残体碳对SOC 库的贡献呈升高趋势。SEM 分析表明,土壤化学性质和酶活性分别是对0~10 和10~20 cm 土层土壤微生物残体碳含量影响较大的潜变量,在杉木人工林经营中,可通过适度管理杉木人工林密度提高土壤养分和酶活性,促进微生物生长,并进一步提高微生物残体碳含量及对SOC 库的贡献。
