散射辐射对杨树人工林生态系统总初级生产力的影响*
2023-08-09万家鸣张志强
万家鸣 律 江 石 云 许 行,2 张志强,2
(1. 北京林业大学水土保持学院 北京 100083;2. 山西吉县森林生态系统国家野外科学观测研究站 吉县 042299;3. 北京市共青林场 北京 101300)
从20 世纪90 年代起,由于到达地面的辐射总量不断增加,全球大部分地区都经历了从变暗(到达地面的总辐射减少)到变亮的过程(Wanget al., 2016; Heet al., 2018; Liet al., 2020)。太阳辐射是植物光合作用的必要条件,到达地面的太阳辐射可以分为直接辐射和散射辐射两部分(Kanniahet al., 2012)。研究表明,全球变亮引起辐射总量及其组分比例的变化对陆地生态系统总初级生产力(gross primary productivity,GPP)具有重要影响(Zhouet al., 2020; Guiet al., 2021;Xueet al., 2021; Zhouet al., 2021a)。具体而言,散射辐射的减少使阴叶接收到的光照减少,从而降低整个冠层的生产力和光能利用效率(Matsuiet al., 2008)。与此同时,到达冠层的总辐射和直接辐射的增加,在一定范围内会显著提高阳叶的光合速率。例如,全球变亮引起总辐射的增加使得美国玉米产量增加了27%(Tollenaaret al., 2017)。因此,总辐射的增加和散射辐射的减少之间存在着一定的权衡效应(Mercadoet al., 2009; Hanet al., 2019),评估并量化散射辐射和直接辐射对生态系统光合作用的影响对于探究这一权衡关系至关重要。然而,目前这方面的研究相对较少。
散射辐射可以促进生态系统总初级生产力并增强陆地碳汇(Kanniahet al., 2012; Ezhovaet al., 2018;Zhanget al., 2020a; Zhouet al., 2021a; Guiet al., 2021)。Gu 等(2002)研究对比了散射辐射和直接辐射对苏格兰松树林、杨树林、落叶混交林、高草草原和冬小麦作物的影响,发现散射辐射增加会提高植物冠层的光能利用效率。Zhou 等(2021a)对全球通量网中200 多个站点研究发现,每增加1 W·m-2的散射辐射,GPP 增加0.44%~1.01%,散射辐射使GPP 提高的程度大约是相同数量直接辐射的2.5 倍。另一项模拟研究也发现,1981—2015 年散射辐射增强使全球GPP 增加了1.49 g·m-2d-1,占总GPP 的64.3%(Zhouet al., 2021b)。散射辐射比例从0.3 增至0.5,加拿大森林和湿地生态系统的GPP 分别增加了18%和7%(Leeet al., 2022),散射辐射增加也会使生态系统生产力显著提高(Xieet al., 2020)。因此,在全球辐射总量及其比例显著变化的背景下,认识和理解森林生态系统光合作用对散射辐射变化的响应机制,对于准确评估其碳汇能力、预测其生态服务功能和评价其适应性和稳定性均具有十分重要的意义。
生态系统表观量子效率(α)和最大光合速率(Amax)可描述生态系统光合作用的潜力,是确定光响应曲线和评估陆地生态系统功能的重要参数(Ruimyet al.,1995)。通常,α 和Amax被认为是植物或生态系统的固定光合属性,但事实上α 和Amax具有一定的季节动态并且受到多种生物物理因素的影响,如叶面积指数(LAI)、辐射、温度(Ta)和土壤含水量(SWC)等(Ehleringeret al., 1983; Zhonget al., 2016)。不同类型植被和生态系统的α 和Amax对温度的响应存在差异:在温带混交林和芦苇湿地生态系统,α 和Amax随温度升高呈指数增长(Zhanget al., 2006; Zhouet al., 2009);草地生态系统的α 和Amax随温度升高呈线性增长(Fuet al., 2009);而在亚热带常绿阔叶林,α 与温度呈较弱的负相关关系,Amax则与温度无显著关系(Zhanget al., 2006)。此外,LAI 的增加则对α 和Amax有促进作用(Gaoet al.,2017)。因此,定量研究特定生态系统α 和Amax随环境条件的变化特征,对于明确未来辐射变化背景下陆地生态系统的功能至关重要。
本研究针对位于我国华北平原的杨树人工林生态系统,定量研究散射辐射和直接辐射对α 和Amax的影响,分析环境要素对Amax的调控机制,利用模型拆分并量化生态系统散射辐射和直接辐射所对应的α和Amax,并探究其季节动态;明确生物物理因素对散射辐射所对应的Amax的影响;探究散射辐射对生态系统总初级生产力的影响机制。
1 研究区概况
北京市顺义区共青林场(116°42′41″E, 40°06′27″N)属典型的暖温带半湿润大陆性季风气候,四季分明。根据顺义气象站(116°37′E, 40°08′N)的长期观测,该地区年均气温11.5 ℃,年均降水量576 mm,95%的降水发生在生长季(4—10 月)。年均相对湿度50%,夏季(6—8 月)总日照时数690 h。土壤为砂土,透水性强,保水能力较低。研究期间,地下水位稳定在2 m 左右。
试验区主要树种为欧美杨(Populus euramericanacv. '74/76')人工林,于1996 年按4 m × 3 m 的株行距栽植,面积178 hm2。2015—2018 年杨树平均树高为(18.65 ± 1.12)m; 平均胸径为(25.90 ± 1.74) cm。2015—2018 年生长季的最大叶面积指数(LAI)分别为(3.08 ± 0.06)、(3.36 ± 0.08)、(3.26 ± 0.09)和(2.87 ±0.07) m2·m-2。林下植被以红瑞木(Swida alba)、珍珠梅(Sorbaria sorbifolia)、连翘(Forsythia suspense)和沙地柏(Sabina vulgaris)为主,林下植被地上生物量约为0.06 kg·m-2。
2 研究方法
2.1 通量和气象观测
本研究采用涡度相关技术,利用开路式红外气体分析仪(LI-7500,LI-COR,USA)和三维风速仪(R3,Gill Instruments,UK),对CO2浓度的脉动和三维风速开展长期定位观测,以获取净生态系统生产力(net ecosystem productivity, NEP)。其中红外气体分析仪根据CO2在可见光中的特定吸收波峰来测定其浓度,该方法是目前对陆地生态系统和大气间碳通量进行原位和连续观测的标准方法(Baldocchi, 2020; Papale,2020; 陈世苹等, 2020)。在杨树人工林中心建造了一座30 m 高的塔,涡度相关系统安装在通量塔28.5 m处。在距地表0.5、1.5、5、15 和30 m 处,安装5 个温度和湿度传感器(155A,Campbell Scientific Inc.,USA),用于测量Ta和相对湿度(RH)。塔顶安装有总辐射表(LI-200x,LI-COR,USA)、光量子传感器(LI-190SB,LICOR,USA)和净辐射仪(CNR1,Kipp & Zonen.,NL),用于测量总辐射(G)、光合有效辐射(PAR)和净辐射(Rn)。用太阳光照辐射计(BF5,Delta-T,UN)测量散射辐射。通量和微气象数据分别记录在CR3000 和CR1000(Campbell Scientific Inc.,USA)中。用植物冠层分析仪(LAI-2200C,LI-COR,USA)测定叶面积指数。
2.2 通量数据处理和质量控制
本研究采用共青林场杨树人工林2015—2018 年生长季的通量数据和微气象数据。通量数据处理利用Eddypro(v7.0.6)完成包括去除异常值、平面坐标转换(Fokenet al., 2012)、WPL 校正(Webb, 1982)、通量数据质量分析以及缺失数据的插补等处理。对处理后的数据进行稳态测试(Foken, 2005),将所有数据分成9 级。舍弃风向在> 350°和< 45°(北偏角)之间湍流强度低的第9 级和第8 级数据。摩擦风速低于阈值的数据被舍弃(Foken, 2005)。同时使用KM(Kormann and Meixner)模型去除研究区域以外的数据(Kormannet al., 2001)。由传感器障碍或随机电信号异常造成的异常值、超出临界值的数据、超出原始数据偏锋和峭斜度标准范围的数据被舍弃。最后利用线性内插法(小于2 h 的缺失数据)和边际平均采样(MDS)法对缺失数据进行插补(Foken, 2005),使数据尽可能形成完整的30 min 通量序列。2014—2017 年半小时数据计算的生长季能量平衡比为0.79,表明该站点的涡度相关法观测数据可靠(Xuet al., 2020b)。
2.3 数据计算
根据观测的净初级生产力(NEP)和采用短期温度敏感性方法模拟的生态系统呼吸(ER)之和计算生态系统总初级生产力(GPP)(Reichsteinet al., 2005; Xuet al., 2020b),公式如下:
式中,Rref是参考温度下的ER,E0,short表示决定短期温度敏感性的活化能参数(本研究中为4 天),Tref是参考温度(10 ℃),T0为常数(-46.02 ℃),Tsoil是5 cm 处的土壤温度。
Prestley-Taylor 系数(η)是实际潜热(LE)与湿表面潜热理论值(LEeq)的比值,能够解释土壤水分对LE的限制程度。计算方法为:
式中,H代表显热(W ·m-2), γ为干湿球常数(67 Pa·K-1),Δ 为饱和水汽压曲线的斜率(Pa·K-1),Tmean表示日平均空气温度(℃)。一般来说,η ≥ 1 出现在水分供应充足的潮湿地表,表明LE 主要受能量的限制。而η <1 一般出现在干燥的生态系统中,说明此时LE 受限于水的供应(Krishnanet al., 2012)。
2.4 散射辐射测定、计算与验证
散射辐射(PARdif)由晴空指数(CI)和太阳高度角(β)计算得到(Reindlet al., 1990; Xuet al., 2018),计算方法为:
当0 ≤ CI ≤ 0.3 且Gf/G0≤ CI 时,
当0.3 < CI ≤ 0.78 且0.1 CI ≤Gf/G0≤ 0.97CI 时,
当0.78 < CI 且Gf/G0≥ 0.1CI 时,
式中,fDPAR 代表散射光合有效辐射比例,Gf是全球太阳辐射的散射辐射分量(W·m-2)。
晴空指数(CI)是在树冠上方观察到的太阳辐射(G)与地外全球水平太阳辐射度(G0)的比率(Guet al.,1999; 张弥等,2009),其计算方法如下:
式中,Gsc为太阳常数(1 370 W·m-2),d为日序数。
研究分析了2015—2018 年数据,由于站点2018年才安装散射辐射观测装置,考虑到散射辐射模拟的准确性和验证的充分性,本研究使用2018、2019 年散射辐射实测数据对模拟的散射辐射进行验证。
2.5 光响应方程参数计算
Emmel 等(2020)比较18 个光响应方程后发现,拆分直接辐射(PARdir)和散射辐射(PARdif)影响的直角双曲线模型(Michaelis-Menten)(公式(15))具有较好的表现。直角双曲线模型具有坚实的生态学理论基础和广泛且可靠的应用(Chenet al., 1999; Xuet al.,2017)。依据Emmel 等的计算方法步骤,利用散射辐射、直接辐射和总初级生产力代入拆分直接辐射和散射辐射影响的直角双曲线光响应模型来评估生态系统尺度的表观量子效率和最大光合速率:
式中,α 为表观量子效率,Amax为最大光合速率,PAR为光合有效辐射,PARdir是直接光合有效辐射,PARdif是散射光合有效辐射。散射辐射比例DF = PARdif/(PARdir+ PARdif)。αdir和αdif分别表示直接辐射和散射辐射影响下的表观量子效率。Amax,dir和Amax,dif分别表示直接辐射和散射辐射影响下的最大光合速率。为确保在一定辐射水平下能够进行光合作用,保证分析的有效性,本研究选择白天(PAR > 4 µmol·m-2s-1)且无降水(precipitation = 0)时间的通量数据(30 min 平均值)用于拟合光响应方程。
由于直角双曲线模型在模拟GPP 时会出现不饱和现象,本研究采用PAR 为2 000 µmol·m-2s-1时的GPP 作为最大光合速率(Emmelet al., 2020),PAR 为2 000 µmol·m-2s-1时直接辐射和散射辐射的光合同化率可分别计算得出:
式中:A2000,dif和A2000,dir分别表示PAR 为2 000 µmol·m-2s-1时散射辐射和直接辐射对应的光合同化率。αdir和αdif分别表示直接辐射和散射辐射影响下的表观量子效率。Amax,dir和Amax,dif分别表示直接辐射和散射辐射影响下的最大光合速率。
2.6 统计分析
αdir、αdif、A2000,dir和A2000,dif的拟合使用R 4.0.3(R Core Team 2019)的nls 函数进行,要求拟合的光响应参数的P值小于0.05。回归分析与回归曲线绘制采用SigmaPlot (version 12.5,Systat Software Inc.,USA),要求所有选定的回归曲线都具有统计学意义(P<0.05)。方差分析和偏相关分析使用SPSS 25(SPSS Inc.,Chicago, USA),同样要求显著性P< 0.05。将均方根误差(RMSE)作为验证散射辐射计算值有效性的衡量指标之一:
式中:oi为实际观测值;pi为计算值;n为测量次数。
3 结果与分析
3.1 散射辐射计算值与实测值比较
利用2018、2019 年实际观测的散射辐射对计算得到的散射辐射结果进行验证,所得回归直线斜率为0.82,R2为0.87,RMSE 为51.67 W·m-2(P< 0.001)(图1),说明本研究散射辐射的模拟结果可靠,可利用其进行研究。
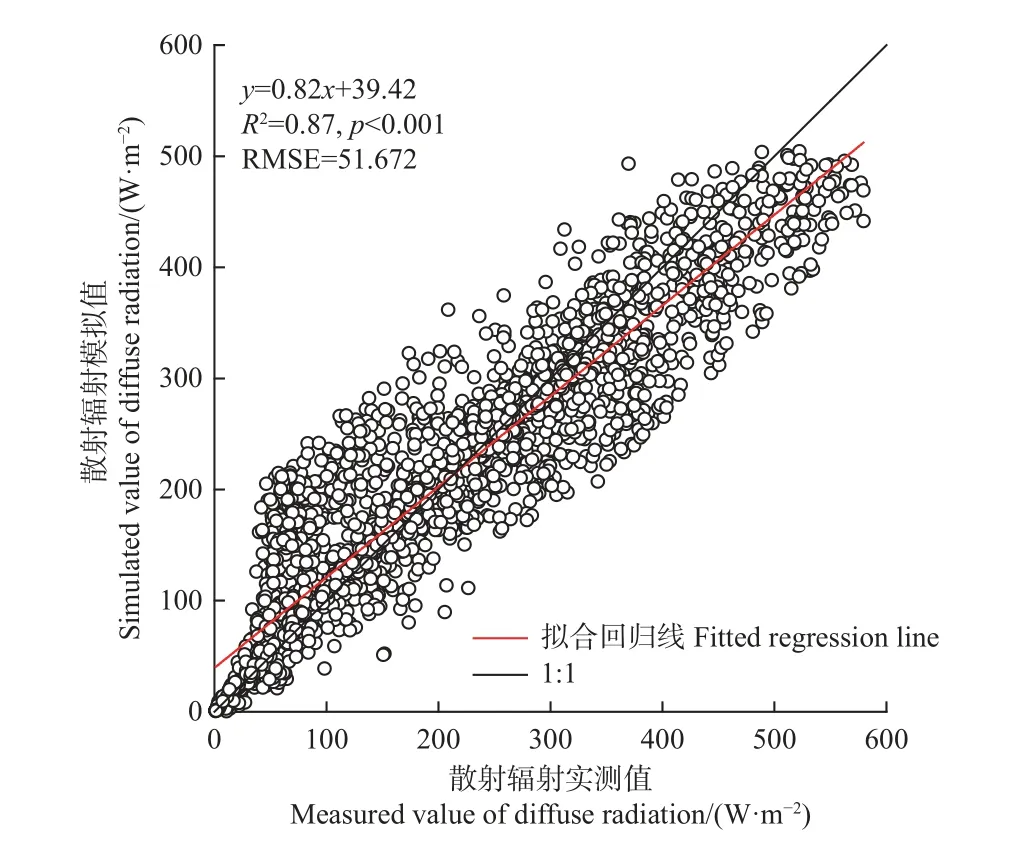
图1 2018—2019 年散射辐射实测值与模拟值比较Fig. 1 Comparison of measured and simulated values of diffuse radiation in 2018—2019
3.2 生长季环境变量和叶面积指数的季节和年际动态
该生态系统2015—2018 年生长季每年月均Ta、VPD、PAR、PARdir、PARdif和LAI 呈现相似的年际变化特征(图2)。月均气温呈单峰变化模式,且在7 或8 月份达到最大值,4 年最大月均值分别为(26.5 ±0.09)、(26.7 ± 0.08)、(27.1 ± 0.09)和(26.9 ± 0.08)℃(图2a)。期间月均VPD 最大值出现在5 月份,分别为1.13、1.59、1.32 和0.97 kPa,之后呈下降趋势(图2a)。PAR、PARdir和PARdif也呈现出明显的季节变化趋势,PAR 和PARdir趋势相似。PARdif在一年内的大多时候比PARdir大,除2015 年外,PARdif在每年的7 月达到最大值(图2b)。与温度类似,各年的LAI也呈现单峰变化趋势,在8 月份达到峰值(图2c)。2015—2018 年生长季的平均LAI 分别为(2.43 ± 0.19)、(2.50 ± 0.21)、(2.45 ± 0.20)和(2.35 ± 0.16) m2·m-2。

图2 2015—2018 年生长季月均气温(a)、饱和水汽压差(a)、光合有效辐射(b)和叶面积指数(c)的变化Fig. 2 Changes in monthly air temperature (a), vapor pressure deficit (a), photosynthetically active radiation (b) and leaf area index (c) during the growing season 2015—2018
3.3 αdir、αdif、A2000,dir、A2000,dif 和GPP 的季节动态
研究期间,该生态系统αdif、A2000,dir和A2000,dif月均值呈现出一致的季节动态变化特征,而αdir无明显季节变化特征(图3)。αdif、A2000,dir和A2000,dif呈先增加后减小的单峰变化趋势,同其气温和LAI 的季节动态相吻合。散射辐射影响的αdif和A2000,dif值显著高于直接辐射影响的αdir和A2000,dir值(pα= 0.02;pA2000= 0.03)。αdif在6 月达到最大值,A2000,dir和A2000,dif在7 月达到最大值。该杨树人工林的αdir值变化在0.027~0.044µmol·µmol-1PAR 之间, αdif值变化在0.034~0.083µmol·µmol-1PAR 之间;A2000,dir变化在 7.51~26.43µmol·m-2s-1之间,A2000,dif变化在13.84~41.40 µmol·m-2s-1范围。αdif与αdir的差值在夏季较大(0.026~0.056µmol·µmol-1PAR), 春 秋 较 小 (0.002 7~0.013µmol·µmol-1PAR);A2000,dif与A2000,dir的差值在春夏较大(10.04~17.06 µmol·m-2s-1), 秋季较小(5.51~8.14µmol·m-2s-1)。在生长季,散射辐射作用的表观量子效率是直接辐射作用的表观量子效率的5.6 倍(αdif/αdir=5.6),散射辐射作用的最大光合速率是直接辐射作用的最大光合速率的6.1 倍(A2000,dif/A2000,dir= 6.1),这2 个比例在不同月份有很大差异,夏季尤为明显。四年生长季GPP 月均值的季节动态也呈现先增加后减少的单峰变化模式(图4),与αdif、A2000,dir和A2000,dif一致。
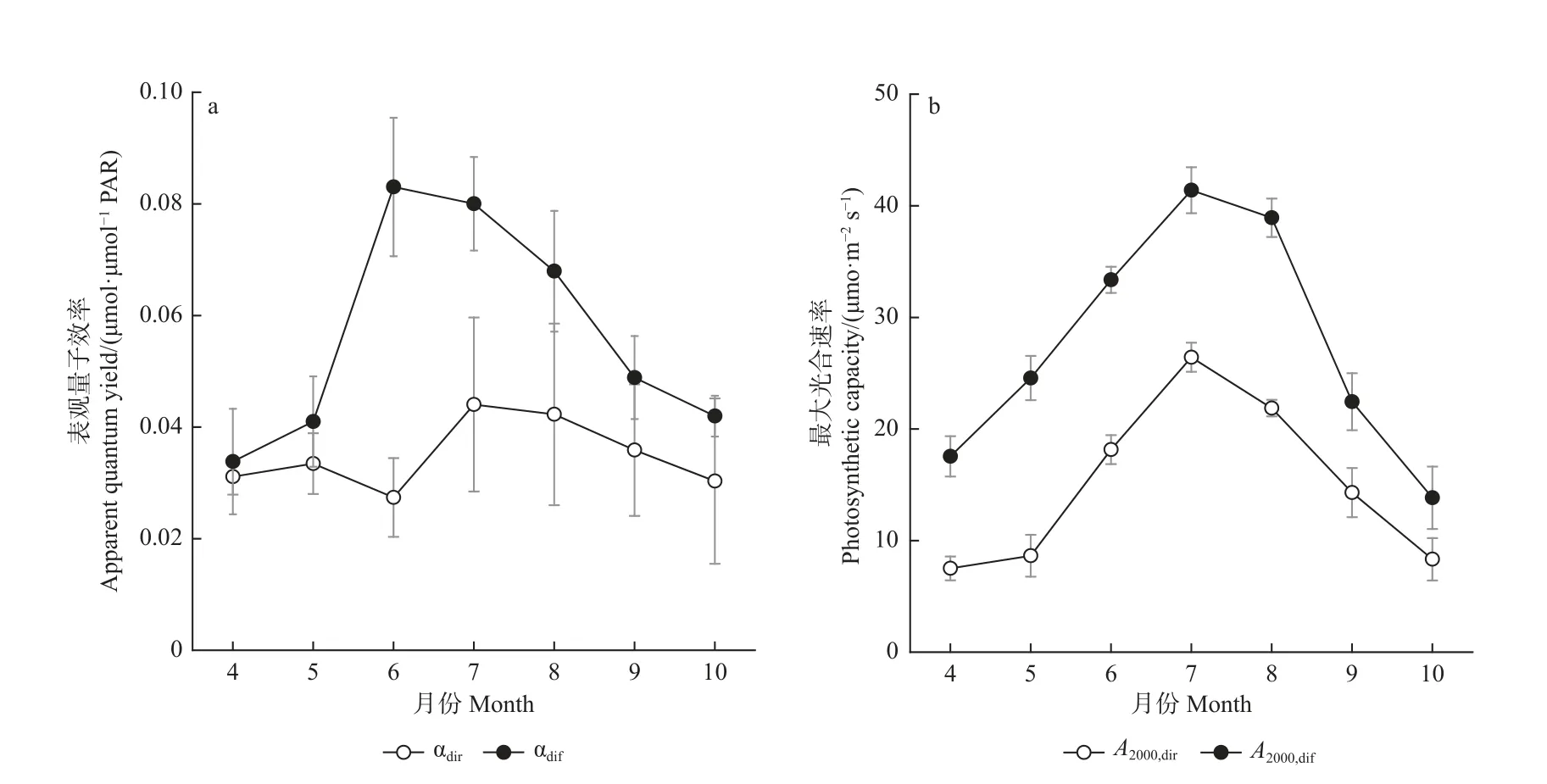
图3 2015—2018 年生长季表观量子效率(a)和最大光合速率(b)平均月变化Fig. 3 Average monthly variation of αdir and αdif (a)、A2000,dir and A2000,dif (b) during the growing season of 2015—2018
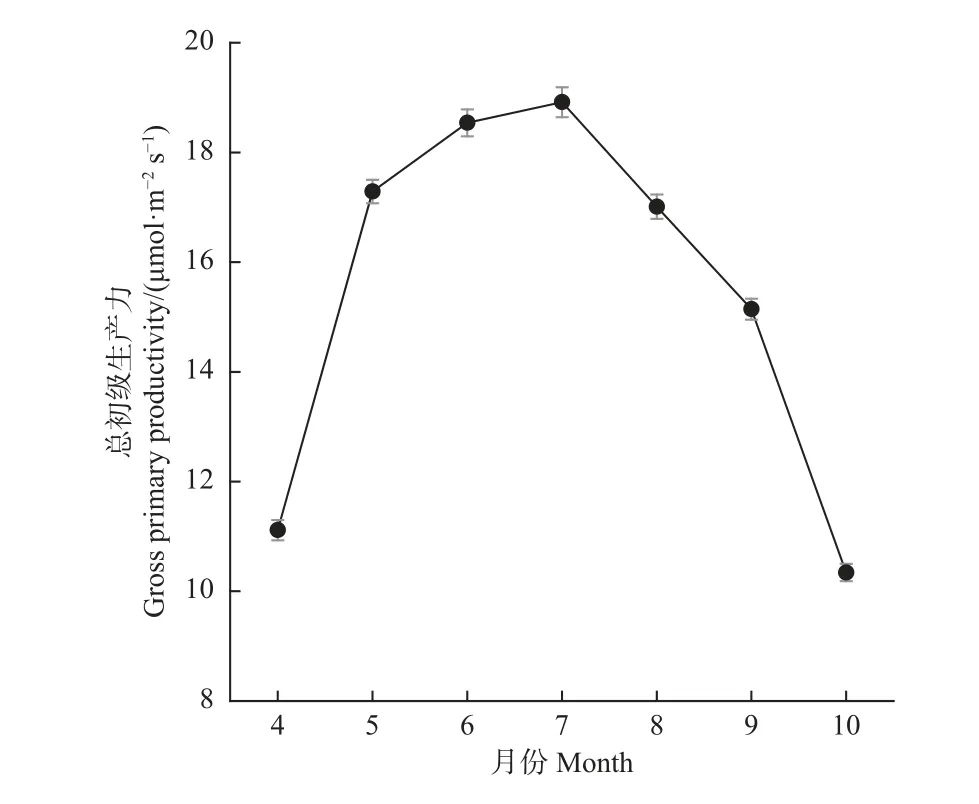
图4 2015—2018 年生长季生态系统总初级生产力月均值的季节动态Fig. 4 Seasonal dynamics of monthly mean ecosystem gross primary productivity during the growing season of 2015—2018
3.4 αdir、αdif、A2000,dir 和A2000,dif 对生态系统生产力的影响
偏相关分析表明,光响应参数中仅A2000,dif与GPP呈显著相关(P< 0.01)(表1),说明A2000,dif是主导该生态系统GPP 的光响应参数。由图5 可知,2015—2018年生长季GPP 与散射辐射和直接辐射比例的关系,GPP 随PARdif/PARdir增加呈显著增加趋势(P< 0.001)。
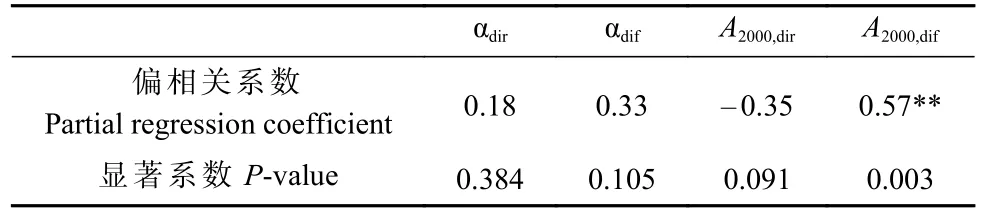
表1 2015—2018 年生长季总初级生产力与αdir、αdif、A2000,dir 和A2000,dif 的偏相关系数和P 值①Tab. 1 Partial regression coefficients and P values between gross primary productivity and αdir、αdif、A2000,dir and A2000,dif during the growing seasons of 2015—2018

图5 2015—2018 年生长季生态系统总初级生产力(GPP)与散射光合有效辐射和直接光合有效辐射比例的对数关系Fig. 5 Ecosystem gross primary productivity (GPP) in relation to the logarithm of the ratio of diffuse photosynthetically active radiation to direct photosynthetically active radiation
3.5 生物物理因素对A2000,dif 的影响
由于GPP 主要受到A2000,dif的控制(表1),故本研究主要探究生物物理因素如何影响A2000,dif。选取Ta、η、CI 和LAI 与A2000,dif进行偏相关分析,结果见表2。A2000,dif在夏季主要受Ta的影响,A2000,dif随着Ta的增加而增加(P< 0.05),偏相关系数为0.74。而A2000,dif在春秋主要受到LAI 的影响,LAI 与A2000,dif呈显著正相关关系(P< 0.01),偏相关系数为0.69。而η 和CI 无论是在春秋还是夏季,对A2000,dif都没有显著的影响。进一步通过回归分析发现,春秋季LAI 的动态可以解释53%的A2000,dif变化(图6a)。在夏季,温度可以解释A2000,dif49%的变化(图6b)。

表2 2015—2018 年春秋和夏季气象因子与A2000,dif 的偏相关系数①Tab. 2 Partial regression coefficients between meteorological factors and A2000,dif for spring and autumn, and summer of 2015—2018
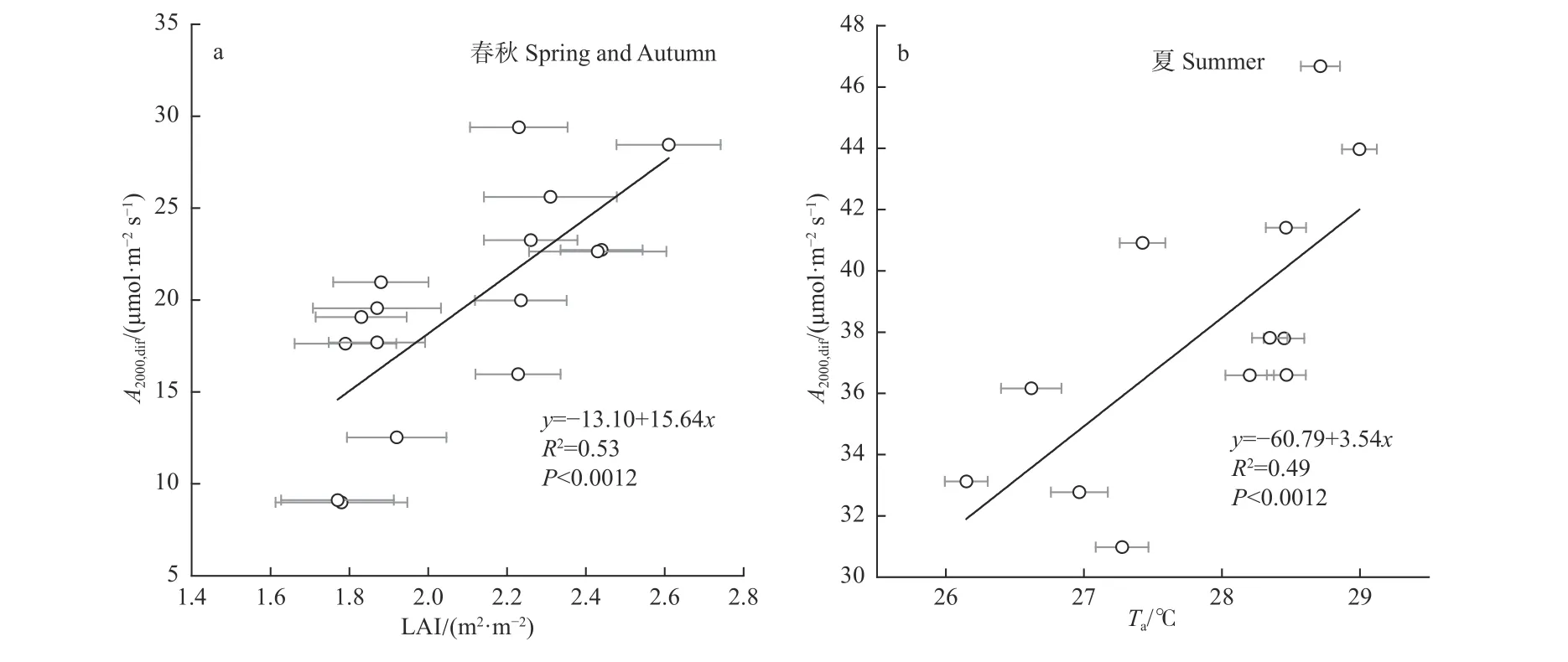
图6 2015—2018 年生长季叶面积指数(LAI)(a)、空气温度(Ta)(b)对散射辐射影响的最大光合速率(A2000,dif)的影响Fig. 6 Effect of leaf area index (LAI)(a) and air temperature (Ta)(b) on maximum photosynthetic rates affected by diffuse radiation (A2000,dif)during the growing season of 2015—2018
4 讨论
4.1 直接辐射和散射辐射对应的α 和A2000 对GPP 的影响
通常来说,α 和Amax是冠层的光合特性,模型中根据所研究的特定植物将它们取做固定值(Yanet al.,2009)。本研究显示,散射辐射和直接辐射对应的α和A2000值有显著差异,并且这些参数存在季节变化,与Emmel 等(2020)对6 种农作物的研究结果一致。忽略光合速率的季节性差异,可能导致落叶阔叶混交林NEP 高估50%(Wilsonet al., 2001)。本研究中,杨树人工林在生长季的αdif/αdir和A2000,dif/A2000,dir分别为5.6 和6.1,表观量子效率的比值大于Emmel 等(2020)研究的6 种农作物的表观量子效率比值(αdif/αdir= 4)。
散射辐射对应的A2000和α 更大,这表明在直接辐射和散射辐射同步变化的情况下,散射辐射变化对该生态系统GPP 的影响更重要。此外,在光响应方程的4 个参数中,A2000,dif是决定GPP 变化的最关键因素(表1)。随着PARdif/PARdir的增加,在同等光照水平下,散射辐射所占比例增加,最大光合速率受散射辐射的影响大于受直接辐射的影响,因此,生态系统GPP 也会增大(图5)。在区分了阴阳冠层的“双叶”光能利用效率模型(TL-LUE)中,阴冠层的光能利用效率是阳冠层的2.5~3.8 倍(Heet al., 2013)。这说明,相比于直接辐射,冠层利用散射辐射的效率更高。
4.2 散射辐射对GPP 的影响
散射辐射增加能提高生态系统的GPP(Guet al.,1999; Mercadoet al., 2009; Zhouet al., 2021)。一方面,散射辐射增多促进光对树冠的渗透,从而使光在叶片之间的分配更均匀(Ryuet al., 2019);另一方面,散射辐射增加为光合作用创造更有利的微气象条件(Zhanget al., 2020b; Xieet al., 2020)。此外,散射辐射可增加蓝/红光比例,从而刺激光化学反应和气孔开放(Urbanet al., 2007)。Xu 等(2017)的研究表明,散射辐射是中国北方杨树人工林GPP 最重要的驱动因素。模拟结果显示,散射辐射使中国陆地生态系统的GPP每年增加3.20 g· m-2(Xueet al., 2021)。还有研究表明,散射辐射施肥效应随LAI 的增加而增加(Fuet al.,2009),在冠层较复杂的生态系统中,PARdif对GPP 的相对贡献更大(Guiet al., 2021)。本研究区林分未完全郁闭,但是最大LAI 大于3 m2· m-2,当阳冠层达到光饱和时,散射辐射对GPP 的影响明显(Wohlfahrtet al., 2008),整个冠层的GPP 由散射辐射主导。
但也有一些研究发现,散射辐射对生态系统光合生产力的影响也会受到其他因素的调控。Alton 等(2008)对阔叶林、针叶林、混交林和草地的研究表明,即使散射辐射增加,但由于总辐射急剧下降,使GPP降低了10-40%。散射辐射的促进作用也可能被直接辐射减少所抵消,对苏格兰松林的研究表明,散射辐射比例超过阈值0.75 时,冠层的LUE 基本不会变化(Altonet al., 2005)。但是,这些结果也表明,其他生物物理因素也影响着散射辐射在权衡效应中的主导地位(Luet al., 2017; Zhanget al., 2020b)。陆地植物的生产力随散射辐射增加而增长,前提要考虑水、温度和营养物质等是否受到限制(Kanniahet al., 2012)。
4.3 生物物理因素对A2000,dif 的影响
本研究表明,A2000,dif的变化不是直接由散射辐射和直接辐射比例的不同导致的,而可能是受到了一些生物物理因子的影响。生态系统冠层特征,如叶面积指数、冠层高度等,是影响生态系统光合作用的重要因素(Fuet al., 2009; Chenet al., 2019; McKendryet al.,2019; Emmelet al., 2020)。在本研究区,春秋季节,A2000,dif主要受LAI 的控制,而在夏季,Ta是控制A2000,dif的主要因素(表2)。A2000,dif在叶片扩张期(4—6 月)急剧增加,在夏季中期变化相对较小,然后在秋季叶片衰老期(9、10 月)下降明显,说明LAI 的季节性变化与A2000,dif在生长季的变化趋势有一定的联系。类似地,对中国黄土高原玉米耕地的研究表明,每增加单位LAI,Amax增加1.12 mg·m-2s-1(Gaoet al., 2017)。Emmel等(2020)对瑞士多种农作物的研究发现,A2000,dif与LAI 呈正相关。夏季生长季中期A2000,dif主要受Ta影响,与在中国温带混交林(Zhanget al., 2006)和草地(Fuet al., 2009)的研究结果一致。夏季的较高温度能够提高光合作用酶的活性以及光合电子传输效率(Berryet al., 1980),从而使冠层光合速率增强。同时,直接辐射和散射辐射均在夏季达到峰值(图2),植物冠层能接收更多的辐射,因此GPP 也更大。本研究发现,土壤水分对杨树人工林的A2000,dif没有显著影响,这可能是由于研究地点靠近河岸,地下水位高,生态系统未缺水(Xuet al., 2018; Xuet al., 2020a)。CI 与A2000,dif无显著相关关系,表明天空状况的变化没有引起散射辐射对应的光合速率发生明显改变。
5 结论
本研究基于对我国华北地区广泛种植的杨树人工林生态系统开展的长期定位观测,通过拆分、量化直接辐射和散射辐射对应的表观量子效率和最大光合速率,从生态系统表冠量子效率和冠层光合能力的角度,发现辐射组分变化主要通过影响散射辐射对应的最大光合速率对生态系统总初级生产力产生重要影响。散射辐射和直接辐射对应的α 和A2000都存在季节动态。散射辐射影响的αdif和A2000,dif要显著高于直接辐射影响的αdir和A2000,dir,这种差异在生长季中期更明显。A2000,dif在春季和秋季主要受LAI 的控制,而在夏季,Ta是控制A2000,dif的主要因素。A2000,dif是影响杨树人工林生态系统GPP 动态的主要因素,因此,在散射—直接辐射的协同变化过程中,散射辐射主导着该杨树人工林生态系统的GPP。
