钙胁迫对黑骨藤种子萌发及幼苗生长的影响
2023-08-01姜晓琪杨荣艺李恒谦龙金兰吴之坤
姜晓琪, 杨荣艺, 李恒谦, 龙金兰, 吴之坤
(1.贵州中医药大学, 贵阳 550025; 2.毕节市中药研究所, 贵州 毕节 551700)
钙是植物体内必需的营养元素,是组成植物细胞壁的重要组分,对植物的发育起着极其重要的作用,同时还能够抵抗胁迫对植物造成的危害[1]。贵州属于典型的喀斯特地貌,在喀斯特环境下,钙离子过多会抑制植物的生长,同时还会影响农作物的产量等[2]。对于大多数的植物来说,种子萌发及幼苗生长期是其生长最脆弱的时期,在种子萌发以及幼苗期对胁迫环境适应能力的大小是决定其植物分布范围及种群生存能力的关键性因素,高钙的喀斯特特殊生境,可以促进植物的抗逆性特化发展[3]。因此,研究在贵州喀斯特地区生长的植物适应高钙胁迫的机制具有重要意义。
黑骨藤为贵州少数民族地区苗族和布依族较为常用的药物,其药材来源于夹竹桃科(Apocynaceae)杠柳属(Periploca)植物黑龙骨(PeriplocaforrestiiSchltr.)的全株,具有祛除风湿、活血化瘀、消炎、杀菌等功效,主要用来治疗跌打损伤、风湿性骨痛和风湿性关节炎等疾病[4]。基于以上优点,以黑骨藤为主药开发了黑骨藤追风活络胶囊、黑骨藤伸筋透骨液、苗药黑骨藤追风液等系列的产品,具有良好的发展前景。黑骨藤为多年生常绿藤本植物,其生长环境为海拔2 000 m以下阳光较为充裕的山坡杂林灌丛中,以及高山岩壁上,主要分布于贵州、广西、四川、重庆、云南等地的喀斯特地貌环境下[5]。目前,黑骨藤多为野生品种,很少有人工种植栽培,国内外有关黑骨藤的研究主要集中在野生资源的化学成分、生药学、质量控制、药理作用以及临床应用等方面[6-10],而在喀斯特环境下钙胁迫对黑骨藤种子萌发及幼苗生长影响的研究尚未见报道。鉴于此,针对喀斯特地区土壤钙离子浓度高的这一特点,以黑骨藤种子和幼苗期植株为研究对象,通过测定不同浓度CaCl2胁迫环境下黑骨藤种子和幼苗相关萌发指标、生长指标及生理指标,研究其对黑骨藤种子萌发及幼苗生长的影响,为今后黑骨藤的引种繁育研究提供一定的数据参考。
1 材料与方法
1.1 材 料
样品经贵州中医药大学魏升华教授和吴之坤副教授鉴定为黑骨藤,于2020年11—12月,在贵州中医药大学资源圃(原植株引种于修文县)采集黑骨藤种子进行相关研究实验。种子采集后晾于通风处,进行除杂后备用。
1.2 方 法
1.2.1种子处理
黑骨藤种子除杂后置于3层滤纸的培养皿(直径15 cm)作为发芽床,通过预实验,每皿分别用纯化水配制0,10,30,60,90,120 mmol/L的CaCl2溶液,以CaCl2浓度0 mmol/L为对照组。用5%次氯酸钠对黑骨藤种子进行消毒,处理8 min后,再用蒸馏水冲洗种子数遍,然后放入发芽床中,每个处理75粒种子,3次重复,每2 d更换1次发芽床。于人工气候箱内(25 ℃,光照16 h/黑暗8 h,相对湿度60%)连续培养,每天18:00时用注射针管定量加入CaCl2溶液。
发芽试验期间每天定时观测种子的萌芽状况,种子萌发以胚根露出种皮1 mm为标志,记录每个培养皿内种子萌发数和胚根胚轴的长度,萌发结束以萌发末期连续3 d萌发粒数不足供试种子总数的1%为标志。试验处理35 d后测定种子萌发后幼苗的苗高、叶片数(新叶片数)、叶长/叶宽、根长、根粗、须根数等生长指标。
1.2.2幼苗处理
种子处理后置于人工气候箱内(25 ℃,光照16 h/黑暗8 h,相对湿度60%)连续培养。种子露白后转移至装有已消毒土壤基质的育苗盘中进行培养备用,每天下午用纯化水进行喷洒,育苗盘保持湿润。经过实验观察,当育苗盘中的黑骨藤幼苗长出2~3对真叶后,将长势一致的黑骨藤幼苗移栽至一个装有混合土壤(V腐殖土∶V蛭石=2∶1)的花盆(直径10.3 cm、高8.5 cm)中,用纯化水一次性浇透,让其在花盆中适应一周后,黑骨藤幼苗无萎黄的现象,长势正常即可进行胁迫试验。试验前,分别配制浓度为10,30,60,90,120 mmol/L的CaCl2溶液,试验时以浇纯化水作为空白对照组,分别用不同比例的CaCl2溶液进行定期定时定量淋洒,每天18:00时用称重法补至原始重量。
试验处理45 d后测定幼苗株高、叶片数(新叶片数)、叶长/叶宽、根长、根粗、须根数等生长指标。采用紫外分光光度法测定过氧化物酶(POD)活性、可溶性糖含量,采用ELISA检测试剂盒测定超氧化物歧化酶(SOD)活性,采用微量法测定谷丙转氨酶和谷草转氨酶活性,采用分光光度法测定叶绿素含量。
1.3 数据分析
采用Microsoft Excel 2016软件进行数据处理和作图,使用SPSS 26.0软件进行单因素方差分析及显著性差异比较。
2 结果与分析
2.1 不同浓度CaCl2溶液对黑骨藤种子萌发指标的影响
由表1可知,随着钙胁迫浓度的增加,黑骨藤种子的发芽势、发芽指数均呈降低趋势;其萌发率在30 mmol/L以下时差异不显著,30 mmol/L以上浓度其萌发率随着浓度的增加显著下降。对照组的发芽势(44.44%)与萌发率(94.22%)均最高,对照组发芽势与钙浓度处理组相比,差异极显著。120 mmol/L处理组萌发率最低,除与90 mmol/L差异不显著外,与其他处理组均差异极显著。从发芽指数方面看,对照组黑骨藤种子活力最高,与高浓度钙处理组相比,差异极显著;各处理组间的萌发历期组间差异显著。结果表明,除了萌发率在30 mmol/L以下表现出一定的耐受性之外,经不同浓度钙处理,一定程度上会对黑骨藤种子萌发指标产生影响,抑制种子的萌发活力。
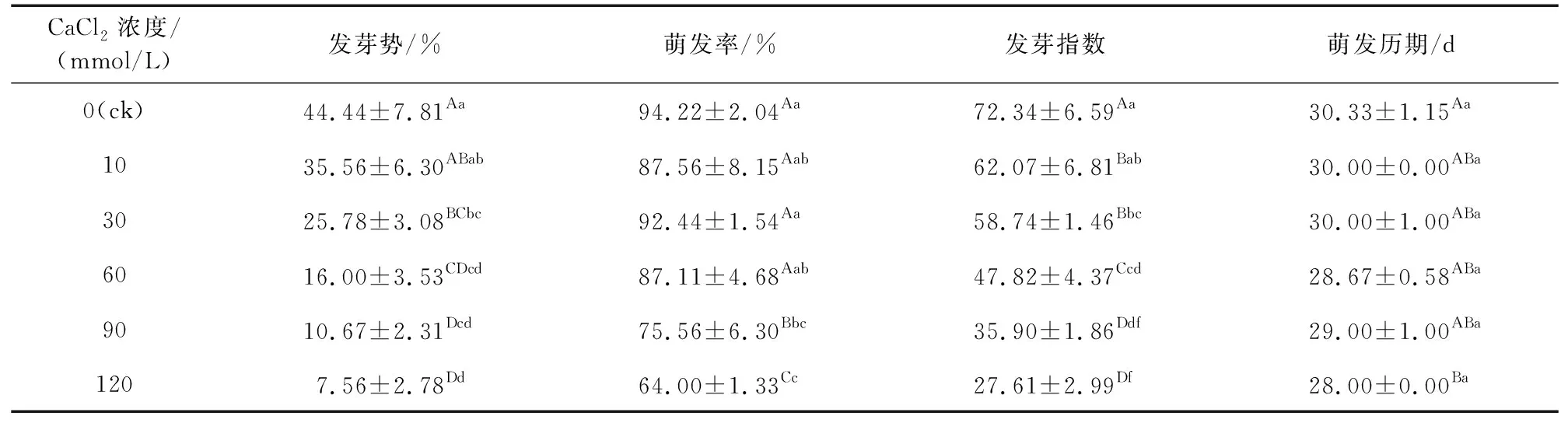
表1 不同浓度钙胁迫对黑骨藤种子萌发特性的影响Table 1 Effects of different concentrations of calcium stress on the seed germination characteristic of P. forrestii
2.2 不同浓度CaCl2溶液对黑骨藤种子逐日萌发率的影响
由图1可知,不同浓度钙处理的黑骨藤种子的萌发高峰期存在差异,低浓度处理组(0~30 mmol/L)的萌发高峰期在第7~15天,高浓度处理组(60~120 mmol/L)的萌发高峰期在第19~28天,相比于低浓度处理组往后延迟了4~6 d。逐日萌发率较高的是对照组和10 mmol/L处理组,对照组的萌发高峰期在第8天,萌发率为14.67%,出现7个发芽波峰;而10 mmol/L处理组的萌发高峰期在第11天,萌发率为15.11%,出现8个发芽波峰;30 mmol/L的萌发高峰期在第12天,萌发率为12.44%,共出现7个发芽波峰;60,90 mmol/L处理组的萌发高峰期均在第24天,萌发率均为12.44%,发芽波峰分别为4个、6个;120 mmol/L处理组的萌发高峰期在第23天,萌发率为12.00%,发芽波峰有6个。随着钙胁迫浓度的增加,黑骨藤种子的逐日萌发率呈下降趋势,且萌发高峰期持续时间也随之降低,说明在高钙胁迫环境下会抑制黑骨藤种子的萌发。
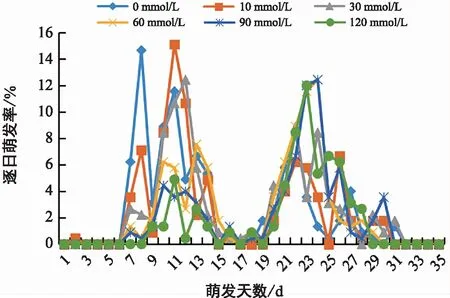
图1 钙胁迫对黑骨藤种子逐日萌发率的影响Fig.1 Effects of calcium stress on day-by-day germination rate of seed of P. forrestii
2.3 不同浓度CaCl2溶液对黑骨藤种子萌发后幼苗生长的影响
不同浓度钙处理组ANOVA分析结果(表2)表明,对照组与各钙浓度处理组的根长、根粗、须根数、茎长、叶长、叶宽、叶片数、苗高组间差异显著。由表2可知,120 mmol/L处理组无真叶长出,而根长和根粗最大,说明高浓度钙胁迫种子无法通过光合作用进行生长,只能借助胚根汲取发芽床水分进行生长,由于钙浓度过高,胚根不断伸长和膨大才能储存足够的水分。
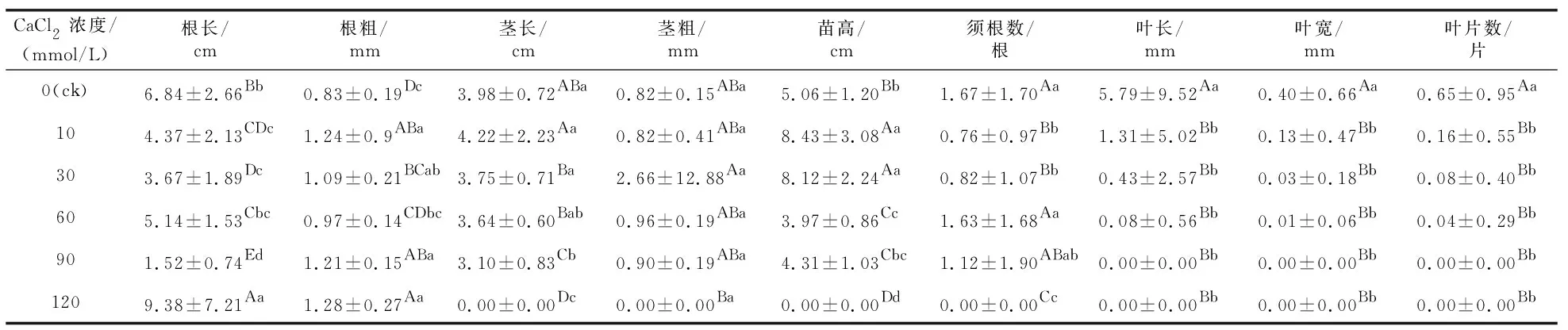
表2 钙胁迫处理35 d后黑骨藤幼苗的生长情况Table 2 Seeding growth of P. forrestii under calcium stress for 35 days
低浓度钙会促进黑骨藤种子的生长,浓度过高反而会抑制种子的生长。从直观形态上看,种子的真叶生长情况随着钙浓度的增大而呈梯度降低,说明钙胁迫会抑制黑骨藤种子叶的生长,进而影响种子幼苗的光合作用。
2.4 不同浓度CaCl2溶液对黑骨藤幼苗生长指标的影响
实验处理45 d后,在不同浓度钙胁迫处理环境下的黑骨藤幼苗均能够适应环境,但是各处理组之间的生长情况存在一定差异。在结束试验后,对各处理组中黑骨藤幼苗的根长、根粗、须根数、叶长、叶宽、新叶片数、株高和侧枝的数量进行测量与统计。由表3可知,30 mmol/L钙胁迫处理组黑骨藤幼苗的根长与10,60 mmol/L两个处理组间差异显著。经不同浓度钙胁迫环境处理下的黑骨藤幼苗根粗与对照组相比均差异极显著,120 mmol/L高浓度钙胁迫处理组的须根数相比于其他处理组具极显著差异。各处理组的叶长与叶宽差异不显著,新叶片数随着CaCl2浓度的增大呈逐渐减小的趋势,各处理组差异不显著。不同浓度CaCl2胁迫对黑骨藤幼苗的株高有明显的影响,如表3所示,随着CaCl2浓度的增大,黑骨藤幼苗株高呈逐渐递减的趋势,各浓度钙处理组与对照组相比差异极显著。10,30,60,90 mmol/L钙浓度处理下的黑骨藤幼苗株高趋于一致且无明显差异,90,120 mmol/L处理组株高无差异。

表3 钙胁迫处理45 d后黑骨藤幼苗的生长情况Table 3 Seeding growth of P. forrestii under calcium stress for 45 days
黑骨藤生长在独特的喀斯特地貌地区,具有耐钙的能力,但是黑骨藤生长在钙浓度过高的环境下也会对其植株的生长造成一定的影响。侧枝的生长也反映了植株的生长情况,从表3可看出,黑骨藤侧枝数量随着钙浓度的升高而降低,对照组明显高于处理组,说明在高钙环境下对黑骨藤幼苗的生长具有抑制作用。
2.5 不同浓度CaCl2溶液对黑骨藤幼苗叶绿素含量的影响
由图2可知,不同浓度钙胁迫对黑骨藤幼苗中的叶绿素含量产生一定的影响,10 mmol/L钙胁迫处理组黑骨藤幼苗叶片中的叶绿素含量累积最高,浓度达30 mmol/L后出现降低的趋势,而后又升高,120 mmol/L处理组的叶绿素含量最低。与对照组相比,各处理组的叶绿素含量差异极显著,说明叶绿素含量的高低与钙胁迫的浓度密切相关,黑骨藤幼苗的光合作用在低浓度胁迫下有促进作用,但是随着钙胁迫浓度的增大,黑骨藤幼苗叶片中叶绿素的含量逐渐降低。高浓度钙胁迫环境下黑骨藤植株光合作用低而导致该胁迫浓度下的幼苗出现萎黄的情况,这与随着钙胁迫浓度的增加,叶片数和侧枝数逐渐减少的结果基本上一致,说明在高钙环境下可以抑制黑骨藤的光合作用。
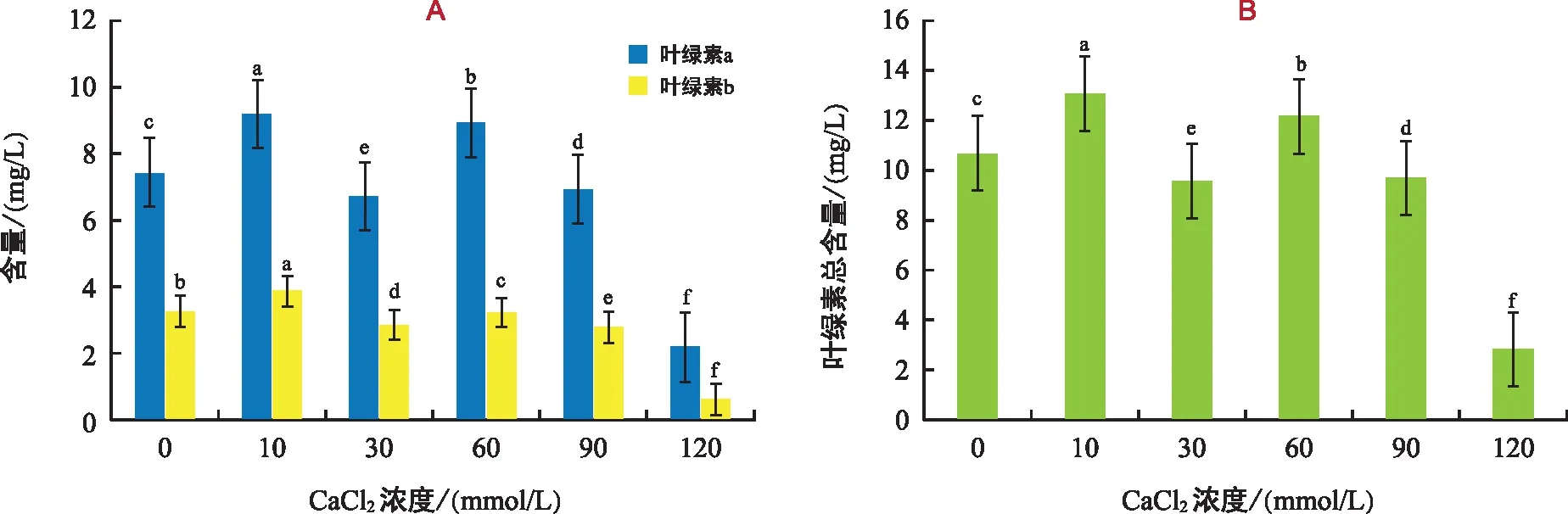
注:不同字母表示各处理组在0.05水平差异显著。下同。图2 钙胁迫对黑骨藤幼苗叶绿素含量的影响Fig.2 Effect of calcium stress on chlorophyll content of seeding of P. forrestii
2.6 不同浓度CaCl2溶液对黑骨藤幼苗谷丙转氨酶活性的影响
由图3可知,黑骨藤叶片中谷丙转氨酶(GPT/ALT)的活性随着钙胁迫浓度的增大而升高,60~90 mmol/L处理组的变化不明显,120 mmol/L处理组的酶活性最高,组间差异极显著。在胁迫实验中,转氨酶会发生催化作用,谷丙转氨酶的含量也会随之升高,说明植物的组织受到了损害。
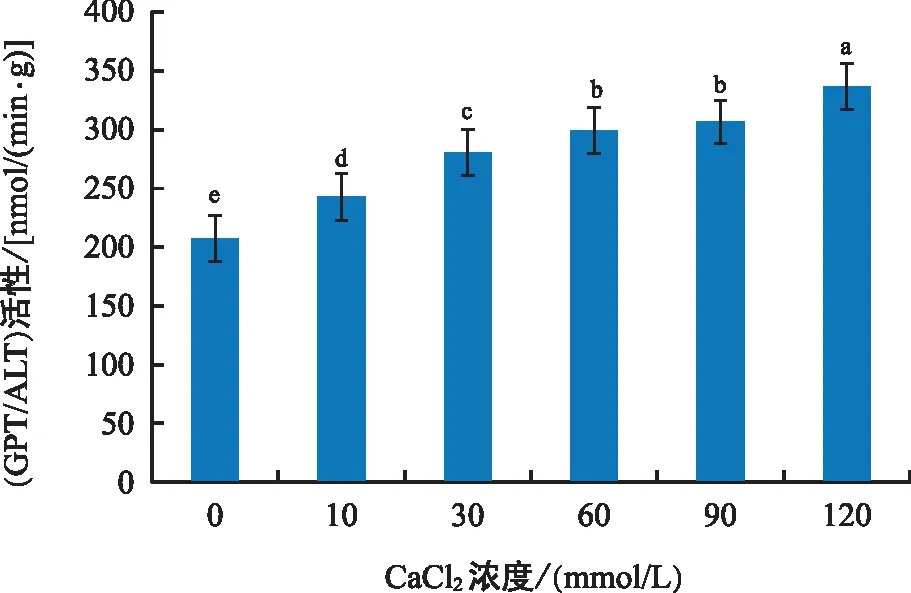
图3 钙胁迫对GPT/ALT活性的影响Fig.3 Effect of calcium stress on GPT/ALT activity
2.7 不同浓度CaCl2溶液对黑骨藤幼苗谷草转氨酶活性的影响
由图4可知,黑骨藤叶片中谷草转氨酶(GOT/AST)的活性随着钙胁迫浓度的增大而呈升高的趋势,其中,120 mmol/L处理组的酶活性最高,组间差异极显著。当植物的组织受到损害时,转氨酶会发生催化作用,谷草转氨酶的含量也会随之升高。
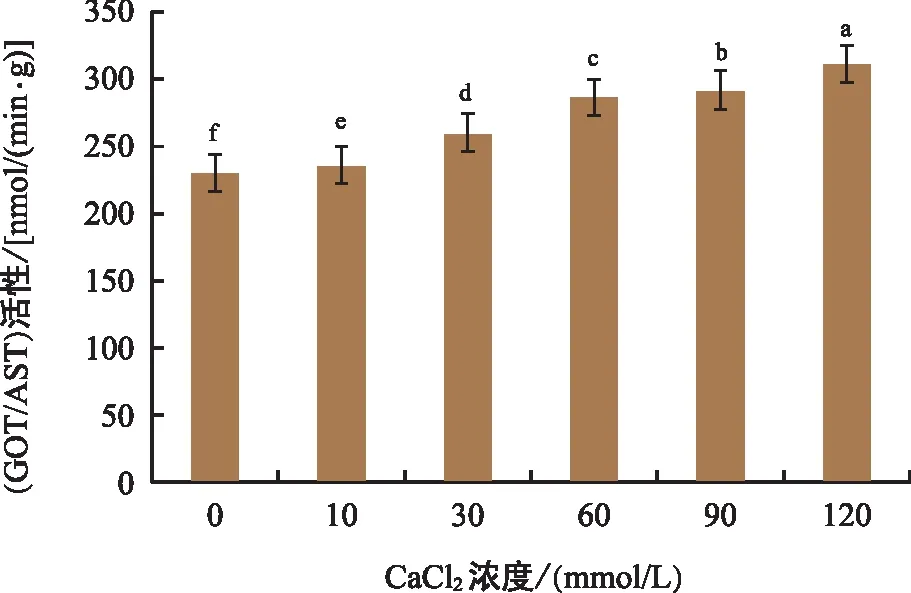
图4 钙胁迫对黑骨藤幼苗GOT/AST活性的影响Fig.4 Effect of calcium stress on GOT/AST activity of seeding of P. forrestii
2.8 不同浓度CaCl2溶液对黑骨藤幼苗POD活性的影响
由图5可知,钙胁迫对黑骨藤幼苗POD活性的影响,随着钙胁迫浓度的增加而呈逐渐升高的趋势,10~60 mmol/L钙胁迫处理组下黑骨藤幼苗叶片的POD活性趋于稳定,90 mmol/L后出现升高的趋势,其中120 mmol/L处理组中黑骨藤幼苗叶片的POD活性最高。与对照组相比,各钙胁迫处理组POD活性均具有极显著差异。POD是反映植物组织老化的一项生理指标,其含量越高,说明植物组织衰老越严重,同时也说明其在生长的过程中产生了活性氧自由基。

图5 钙胁迫对黑骨藤幼苗POD活性的影响Fig.5 Effect of calcium stress on peroxidase activity of seeding of P. forrestii
2.9 不同浓度CaCl2溶液对黑骨藤幼苗可溶性糖含量的影响
由图6可以看出,随着钙胁迫浓度的不断增大,黑骨藤幼苗叶片的可溶性糖含量呈先升后降再升的趋势,60 mmol/L处理组的可溶性糖含量为第一个峰点,90 mmol/L处理组可溶性糖含量出现降低,其中120 mmol/L处理组的可溶性糖含量最高。与对照组相比,各钙胁迫处理组的可溶性糖含量差异极显著,说明黑骨藤幼苗生长在钙胁迫环境下受到了渗透胁迫。
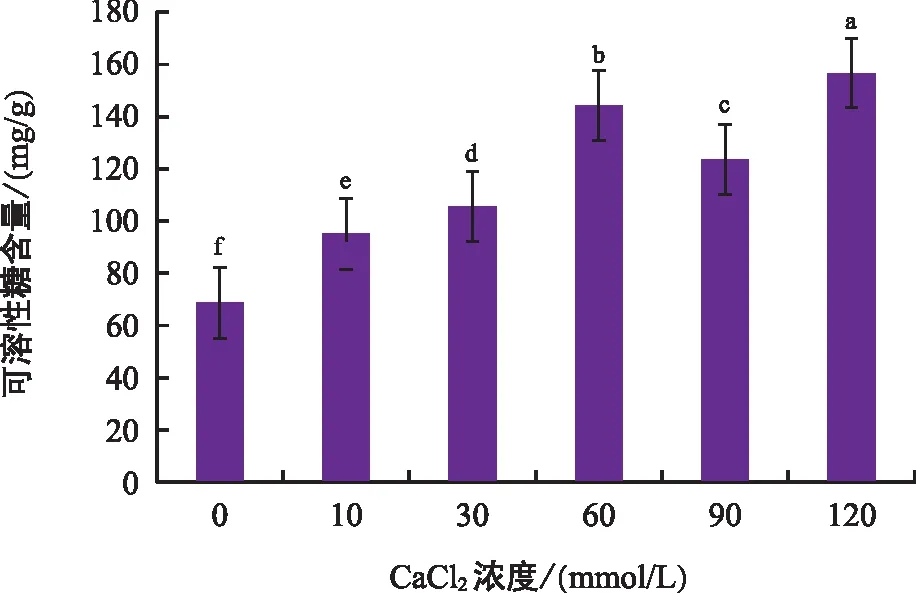
图6 钙胁迫对黑骨藤幼苗可溶性糖含量的影响Fig.6 Effect of calcium stress on soluble sugar of seeding of P. forrestii
2.10 不同浓度CaCl2溶液对黑骨藤幼苗SOD活性的影响
由图7可知,黑骨藤幼苗叶片SOD活性随着钙胁迫浓度的升高呈逐渐升高的趋势,其中,120 mmol/L处理组的酶活性最高,组间差异极显著。说明随着钙胁迫浓度的增加,黑骨藤幼苗体细胞受到了氧损伤。
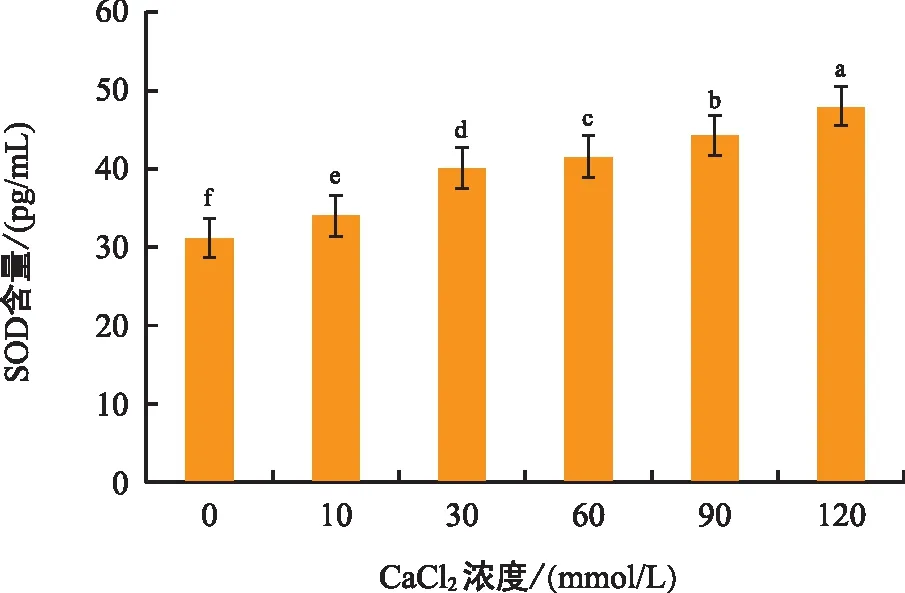
图7 钙胁迫对黑骨藤幼苗SOD活性的影响Fig.7 Effect of calcium stress on superoxide dismutase activity of seeding of P. forrestii
3 讨论与结论
3.1 讨 论
3.1.1不同浓度钙处理对黑骨藤种子萌发的影响
虽然钙离子在一定程度上能维持和调控离子平衡以适应逆境环境的影响,但是当钙离子浓度过高时会抑制植物的生长。有研究发现,钙浓度过高会抑制种子的萌发,对种子的萌发产生抑制作用[11-13]。檀龙颜等[14]通过对金荞麦种子的钙胁迫研究发现,较高浓度的钙离子会对种子产生离子毒害作用,抑制金荞麦种子的萌发和生长。本研究表明,对照组黑骨藤种子的萌发情况最好,但随着钙浓度的增加黑骨藤种子萌发受到抑制。本实验中未出现处理组种子不发芽的情况,从浓度的影响程度来看,10~30 mmol/L时钙胁迫强度对黑骨藤种子萌发影响小,这与徐拾佳等[15]在钙离子条件下对矮牵牛种子萌发影响的研究结果一致;而60~120 mmol/L时对其萌发影响较大。本研究表明,黑骨藤种子对钙离子有一定程度的耐受性,但浓度过高对黑骨藤种子的影响就加大,抑制种子的萌发。
就生长指标来看,实验结果表明,低浓度钙胁迫对黑骨藤种子的生长受到的影响小,但当钙浓度达到120 mmol/L时,种子仅露出胚根生长,且其他生长指标均为零,高钙浓度条件下严重抑制了黑骨藤种子萌发后的生长。从真叶生长情况来看,随着钙浓度的增大,黑骨藤种子真叶生长情况(叶长、叶宽、真叶片数)逐渐受到抑制,表明钙浓度过高会抑制黑骨藤种子萌发真叶的生长,进而影响其光合作用。综上分析,说明黑骨藤种子对钙离子有一定程度的耐受力,但是钙浓度过高反而抑制其萌发。所以钙浓度应控制在小于或等于30 mmol/L的范围内,更有利于黑骨藤种子的萌发生长。
3.1.2不同浓度钙处理对黑骨藤幼苗生长的影响
贵州是典型的喀斯特地貌,喀斯特地区的普遍特征是极度缺乏氮、磷、钾的高钙土壤环境[16],当植株生长在这样的环境时,土壤的盐分过多对植物造成一定的危害,同时对它的发育也会产生一定的影响[17]。钙胁迫对植物的生长发育活动有一定的影响,随着钙胁迫强度的增加,其对植物正常的生长会产生一定的负面作用,比如说降低植株的生长高度等。彭博等[18]研究表明,在不同浓度钙胁迫的影响下,狭叶香蒲的根系、株高、叶片等均受到不同程度的抑制。刘梦龙等[19]研究表明,在钙胁迫环境下大白菜的根系、叶片等均受到不同程度的抑制作用。已有研究表明,在钙胁迫环境下,可通过植株的株高及分枝变化反映其生长情况[20]。本研究中,钙胁迫对黑骨藤幼苗的生长影响直观表现是植株高度的变化、新叶片数量和侧枝产生的情况,随着钙胁迫浓度的增大,黑骨藤幼苗的侧枝数、新叶片数量以及株高均随之减小。根系是胁迫环境最直接的受害部位,根据试验结果得知,在不同浓度钙胁迫环境下生长的黑骨藤幼苗根长和根粗均受到了不同程度的抑制,这与植物的根直接接触到盐溶液的原因有关,因为植物的根部最先能感受到盐碱逆境的胁迫信号,是为了其能够适应逆境所产生的生理反应[21]。随着钙含量的增加,当CaCl2浓度达到90 mmol/L和120 mmol/L时,黑骨藤幼苗出现枯死、萎黄的现象,说明在种植栽培黑骨藤时土壤含钙量不宜过高,否则会因钙浓度过高而影响到黑骨藤幼苗的生长与产量。
钙胁迫对植物的生长会产生一定的负面作用,随着钙胁迫浓度的增加,对植株的直观影响主要表现在其生长速度以及新叶片数逐渐下降,从而影响植株的光合作用[17]。柴胜丰等[22]采用不同浓度的CaCl2对金茶花生理指标影响进行实验研究,结果表明,在不同浓度CaCl2胁迫环境下,两种金茶花的生理指标主要表现在叶绿素a、叶绿素b和叶绿素总量的下降,说明在高钙环境的影响下对其光合作用有影响。本研究表明,黑骨藤幼苗叶片的叶绿素含量随钙浓度的升高总体呈下降趋势,尤其在120 mmol/L时含量最低,表明高浓度钙胁迫会影响黑骨藤叶片的光合作用。可溶性糖是光合作用最终产物之一,同时也是植物在逆境下常见的渗透调节物质[23]。黑骨藤幼苗中可溶性糖含量随钙浓度的增加而呈逐渐升高的趋势,说明黑骨藤幼苗在高钙环境下受到了渗透胁迫。
SOD在活性氧代谢中具有重要作用,可以有效阻碍氧自由基对植物造成的毒害作用[24],廖汝玉等[25]研究表明,高钙环境能够引起枇杷小苗叶片中的过氧化氢含量、SOD和POD酶活性增加,表明在高钙胁迫的环境下枇杷小苗产生了氧自由基,并且通过抗氧化酶系统来进行清除。本研究表明,黑骨藤幼苗POD、SOD活性随钙浓度的增大而逐渐升高,说明植物脂膜受到了氧化损伤,即产生了活性氧自由基,也进一步说明了黑骨藤幼苗在生长过程中主要依靠抗氧化酶系统来清除氧自由基;谷丙转氨酶和谷草转氨酶是催化谷氨酸与丙酮酸、草酰乙酸之间的转氨基作用的两种酶,其转氨作用可逆,在氨基酸代谢反应中具有重要作用[26],谷丙转氨酶和谷草转氨酶活性随钙浓度的不断增大呈逐渐升高趋势,说明黑骨藤幼苗在生长过程中通过转氨作用进行代谢反应。
综上所述,在钙胁迫环境下,黑骨藤幼苗对钙离子表现出较强的耐受性,对其幼苗的生长发育及生理指标的变化情况进行分析,进一步证实了其对喀斯特地区富含钙的土壤环境有一定的适应能力。根据调查得知,黑骨藤种子在萌发生长期是喜阴植物,野外分布的黑骨藤多以幼苗为主,分布稀疏,且幼苗难以成长发育为成年植株,反映了其对喀斯特地区土壤含钙环境的适应较弱。
3.2 结 论
本研究探讨了不同浓度钙胁迫对黑骨藤种子萌发及幼苗生长的影响,为黑骨藤的引种繁育研究提供一定的数据参考。研究表明,黑骨藤种子萌发和幼苗在生长发育过程中,CaCl2浓度在低于30 mmol/L的条件下对黑骨藤种子萌发及幼苗生长的影响不显著,黑骨藤种子及幼苗对弱钙环境具有一定的适应性。因此,今后进行黑骨藤种植时,应提前对种植地的土壤含盐量进行分析,选择合适的土壤条件,从而避免在高钙胁迫浓度下对黑骨藤种子萌发和幼苗生长的危害,影响黑骨藤种植产业的发展。
