甘蔗VAMP相关蛋白ScPVA12与甘蔗花叶病毒P3N-PIPO的互作研究
2023-07-20玉泉馨杨宗桃程光远周营栓焦文迪罗廷绪黄国强张木清徐景升
玉泉馨 杨宗桃 张 海 程光远 周营栓 焦文迪 曾 康 罗廷绪 黄国强 张木清 徐景升,*
甘蔗VAMP相关蛋白ScPVA12与甘蔗花叶病毒P3N-PIPO的互作研究
玉泉馨1杨宗桃1张 海1程光远1周营栓1焦文迪1曾 康1罗廷绪1黄国强1张木清2,*徐景升1,*
1福建农林大学 / 农业农村部福建甘蔗生物学与遗传育种重点实验室 / 国家甘蔗工程技术研究中心 / 教育部作物遗传育种与综合利用重点实验室, 福建福州 350002;2广西大学广西甘蔗生物学重点实验室, 广西南宁 530004
植物囊泡膜蛋白相关蛋白PVA12 (plant vesicle-associated membrane protein (VAMP)-associated proteins homolog 12)属于VAP27家族蛋白, 在细胞中介导内质网囊泡运输以及膜融合。甘蔗(spp. hybrid) PVA12应答甘蔗花叶病毒(, SCMV)侵染尚未见报道。本研究从栽培种新台糖22号(ROC22)中克隆了PVA12基因, 命名为。该基因开放读码框(open reading frame, ORF)的长度为735 bp, 其编码长度为244 aa的蛋白。生物信息学分析表明, ScPVA12是一种不稳定的亲水性脂溶蛋白, C端具有跨膜结构域; 二级结构中无规则卷曲占比最高; 进化树分析表明, 该蛋白在单子叶和双子叶植物中存在明显分化。酵母双杂交(yeast two-hybrid, Y2H)和双分子荧光互补(bimolecular fluorescence complementation, BiFC)试验表明, ScPVA12与SCMV-P3N-PIPO蛋白互作。亚细胞定位试验表明ScPVA12定位于内质网。共定位试验表明ScPVA12与SCMV-P3N-PIPO共定位于内质网。实时荧光定量PCR分析发现,基因在甘蔗各组织中均有表达, 在第8节间中的表达量最低, 在正七叶中的表达量最高; SCMV侵染对基因表达影响显著, 在SCMV胁迫下基因先下调表达, 后期恢复正常水平。
PVA12; 甘蔗花叶病毒; P3N-PIPO; 蛋白互作
甘蔗是我国乃至世界上最重要的糖料作物和经济作物[1-3], 同时甘蔗产业也是我国重要的产业之一[4-5]。甘蔗花叶病在我国各大甘蔗产区普遍发生, 对甘蔗生产危害严重[6-9]。感染甘蔗花叶病的甘蔗植株主要表现为植株矮化、生长迟缓、糖分降低、分蘖减少等, 严重影响甘蔗的产量和品质, 发病严重时减产高达50%[10-11], 对甘蔗生产产生了严重威胁[12-16]。甘蔗花叶病病原的虫媒主要是蚜虫, 蚜虫吸取带病蔗株的汁液后, 以非持久的方式将病毒传播到健康植株[17-18]。在生产中, 带毒甘蔗种茎是甘蔗花叶病在田间传播和扩散的主要途径。因此, 培育和使用抗病品种是防治甘蔗花叶病的根本手段。甘蔗花叶病的病原主要为甘蔗花叶病毒(, SCMV)、甘蔗条纹花叶病毒(, SCSMV)和高粱花叶病毒(, SrMV)[19-20], 且SCMV依然是全世界各大蔗区的主要病原[16,21-23]。SCMV属于马铃薯Y病毒科(Potyviridae)马铃薯Y病毒属(), 由正义单链RNA组成, 编码2个多聚蛋白, 并最终水解成11个成熟蛋白, 这11个蛋白从N端到C端分别为P1、HC-Pro、P3、P3N-PIPO、6K1、CI、6K2、VPg、NIa-Pro、NIb和CP[7,17,20,26-27]。其中, P3N-PIPO是在P3顺反子GAAAAAA插入1个A, 移码编码产生的融合蛋白, 是potyviruses的运动蛋白(movement proteins, MP), 定位在质膜和胞间连丝(plasmodesmata, PD), 其中主要定位在质膜[28]。病毒通过在寄主体内组装病毒复制复合体(virus replication complex, VRC)进行复制, 运动蛋白能够将VRC移动至PD[29], 并且能够修饰PD的结构, 扩大其分子扩散极限(size exclusion limit, SEL), 增加其通透性, 使病毒得以通过细胞壁侵染相邻细胞建立系统性侵染[30-31]。运动蛋白介导的VRC的胞内和胞间运动对病毒建立系统性侵染的至关重要[10,18,32-34]。
植物病毒结构简单, 必须依赖于与寄主因子的互作才能建立系统性侵染[35]。分离鉴定与病毒互作的寄主因子基因对于阐明病毒的侵染机制、选育抗病品种具有重要意义[36]。本课题组以SCMV-P3N- PIPO为诱饵, 筛选甘蔗cDNA酵母文库, 获得了与SCMV-P3N-PIPO互作的植物囊泡膜蛋白相关蛋白PVA12 (plant vesicle-associated membrane protein (VAMP)-associated proteins homolog 12), 并从甘蔗品种ROC22中克隆了其编码基因, 命名为。PVA12是动物VAP33 (VAMP-associated protein of 33 kD)在植物中的10个同源物之一[37], 在植物中最早发现的VAP蛋白分子量为27 kD, 因此将植物VAP蛋白家族命名为VAP27 (VAMP-associated protein 27)[38]。近期有文献将植物VAP27蛋白家族的10个成员重新按VAP27-1至VAP27-10编号命名, 其中VAP27-3与PVA12为同一蛋白的不同命名(TAIR网站登录号同为AT2G45140), 2种命名均沿用至今[37,39-43]。
PVA12属于VAP27蛋白家族, 又名为VAP27-3, 是一种内质网(endoplasmic reticulum, ER)定位蛋白[37]。VAP27蛋白广泛存在于真核生物中, 在哺乳动物、酵母和植物中均有VAP27蛋白的相关报道[44-45]。该家族蛋白高度保守的主精子结构域(major sperm domain, MSD)是VAP27蛋白与其他蛋白发生相互作用的必要功能结构域, 还具有典型的螺旋卷曲结构域(coiled-coil domain, CCD)结构域和C端跨膜结构域(transmembrane domain, TMD), 其氨基酸残基高度保守。VAP27蛋白在植物体内各组织中普遍表达, 在细胞中介导内质网囊泡运输以及膜融合, 在细胞内的脂质运输以及液泡的形成中扮演重要
角色[37,42,46-47]。VAP27蛋白在细胞中与多种不同蛋白互作[43,45], 与其他蛋白共同参与内质网和高尔基体之间的脂质运输[37]、液泡的生物合成[46]、维持内吞作用的稳态[48-49]以及调控自噬体的形成[50], 对植物细胞内的物质运输以及内环境稳态具有重要作用。此外VAP27蛋白还是内质网–质膜结合位点(ER-plasma membrane (PM) contact sites, EPCSs)复合体的重要组分之一, 参与了内质网和质膜之间的物质运输[37,43], VAP27蛋白的缺失会使植物表现出内吞作用延迟、影响内吞膜的稳态, 还会出现根系表型异常[43,51], 表明VAP27蛋白参与维持植物的正常生长发育。近期有研究发现VAP27蛋白还可能参与了植物病毒在寄主体内的侵染[52-53]。
甘蔗中PVA12与SCMV-P3N-PIPO互作尚属首次发现, 深入研究其互作机制, 对于阐明SCMV复制囊泡形成、胞内运输及胞间移动分子机制具有重要的科学意义。本研究通过酵母双杂交(yeast two- hybrid, Y2H)和双分子荧光互补(bimolecular fluorescence complementation, BiFC)试验体系验证了ScPVA12与SCMV-P3N-PIPO的互作, 通过实时荧光定量PCR (Real Time Quantitative PCR, RT-qPCR)分析了基因在甘蔗不同组织中的表达情况以及应答SCMV侵染的表达模式。
1 材料与方法
1.1 材料及处理方法
本试验所使用的甘蔗栽培种ROC22组培苗、SCMV-FZ1病毒株系[24]和本氏烟()由福建农林大学国家甘蔗工程技术研究中心提供。在光周期为16 h光照/8 h暗, 光照强度为200 μmol m–2s–1, 温度28℃, 空气湿度60%的条件下, 采用腋芽快繁技术培养ROC22组培苗。待其出现4~5片完全展开的叶片时, 挑选健壮且长势一致的组培苗在光培养期培养1 h后摩擦接种SCMV, 设置3个重复, 每个重复3株, 使用磷酸缓冲液(pH 7.0)摩擦接种的植株作为对照, 取接种叶片, 使用基因特异性引物(表1)检测接种成功与否。在接种后0 h、1 h、2 h、4 h、8 h、12 h、24 h、48 h、72 h、5 d、8 d、14 d取样。从福建农林大学隔离网室中选取处于伸长期的、株高约1.8 m的健壮且长势一致ROC22植株, 取未成熟叶心叶、形态建成叶正一叶、渐衰叶正七叶、未成熟节间第3节间、形态建成节间第8节间和根用于基因的组织特异性表达试验, 选取3株混合作为1个生物学重复, 设3个生物学重复。取样后用锡箔纸包好后放入液氮中速冻, 然后置–80℃冰箱保存备用。
1.2 RNA提取和cDNA合成
在液氮中将采集的样品充分研磨至粉末状, 使用TRIzol (Invotrigen, 美国)试剂并按照其说明书提取总RNA, 使用1.0%琼脂糖凝胶电泳检测提取RNA质量。利用HiScript III 1st Strand cDNA Synthesis Kit (Vazyme, 中国)并按其说明书将提取的不同组织样品RNA反转录成cDNA。
1.3 ScPVA12基因克隆及生物信息学分析
以筛选酵母文库获得的基因序列为参考,通过同源克隆的方法, 在起始密码子和终止密码子附近设计基因的特异扩增引物(表1)。以反转录获得的cDNA为模板, 使用PrimeSTAR GXL DNA polymerase (TaKaRa Bio, 日本)高保真, 克隆基因, 并送生工生物工程(上海)股份有限公司测序。
通过在线网站及分析软件进行生信分析, 包括蛋白质一级结构及理化性质分析(https://web.expasy. org/protparam/)、蛋白质二级结构分析(https://npsa- prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4. html)、蛋白质三级结构预测(https://swissmodel. expasy.org/)、蛋白信号肽预测(https://services. healthtech.dtu.dk/service.php?SignalP-5.0)和蛋白跨膜区预测(https://services.healthtech.dtu.dk/service. php?TMHMM-2.0)。通过NCBI网站(https://blast.ncbi. nlm.nih.gov/Blast.cgi)与Tair网站(https://www. arabidopsis.org/Blast/index.jsp)进行BLAST比对获得ScPVA12与其他物种的同源序列, 利用DNAMAN V6软件对这些序列进行同源性分析, 利用MEGA 7.0软件的邻接法(Neighbor-Joining)构建系统进化树。通过NCBI网站的CD-Search工具https://www. ncbi.nlm.nih.gov/cdd/?term=)和SMART网站(http:// smart.embl-heidelberg.de/smart/set_mode.cgi?NORMAL=1)进行结构域分析预测。
1.4 ScPVA12的亚细胞定位
参照Cheng等[36]的方法, 利用Gateway方法构建ScPVA12的亚细胞定位载体ScPVA12-YFP和P3N-PIPO的亚细胞定位载体P3N-PIPO-CFP, 使用农杆菌GV3101进行侵染。取健康的本氏烟植株, 参照Cheng等[36]农杆菌侵染方法, 将带有重组质粒的农杆菌注射入健康的本氏烟叶片。48 h后在激光共聚焦显微镜(LeicaTCSSP5I)下观察本氏烟叶片表皮细胞中ScPVA12蛋白的定位。用HDEL-RFP标记内质网。YFP的激发光波长为514 nm, 捕获波长为530~590 nm; RFP激发光波长为552 nm, 捕获波长为590~630 nm; CFP的激发波长为442 nm, 采集波长为450~500 nm。采用数字采集图像, 使用LSM 2.6.3软件进行处理。SCMV-P3N-PIPO-CFP载体来自本课题组前期研究工作[36]。
1.5 Y2H验证ScPVA12与SCMV-P3N-PIPO的互作
按照Cheng等[36]的方法构建ScPVA12的诱饵载体pPR3-ScPVA12, 利用Y2H技术验证其与pBT-STE-P3N-PIPO的互作关系。使用酵母二缺培养基(DDO: 缺少亮氨酸和色氨酸的酵母合成限定基本培养基)和四缺培养基(QDO: 缺少亮氨酸、色氨酸、组氨酸和腺嘌呤的酵母合成限定基本培养基)培养共转酵母菌株NMY51, 在平板培养基上添加5-溴-4-氯-3-吲哚-β-D-半乳糖苷(X-Gal)。以pTSU2- APP/pNubG-Fe65组合为阳性对照, 以pPR3-N/ pNubG-Fe65组合为阴性对照。pBT-STE-P3N-PIPO载体来自本课题组前期研究工作[36]。
1.6 BiFC验证ScPVA12与SCMV-P3N-PIPO的互作
参照Cheng等[36]的方法构建YN-ScPVA12和ScPVA12-YC载体, 利用BiFC技术验证其与YN- SCMV-P3N-PIPO的互作。取健康的本氏烟植株, 参照Cheng等[36]农杆菌侵染方法, 将带有重组质粒的农杆菌注射入健康的本氏烟叶片, 48 h后在激光共聚焦显微镜(LeicaTCSSP5I)下观察。YFP的激发光波长为514 nm, 捕获波长为530~590 nm; 叶绿体的的激发光波长为552 nm, 捕获波长为650~680 nm。采用数字采集图像, 使用LSM 2.6.3软件进行处理。YN-P3N-PIPO和P3N-PIPO-YC载体来自本课题组前期研究工作[36]。
1.7 ScPVA12基因的RT-qPCR表达分析
根据基因序列设计特异性实时荧光定量PCR引物(表1), 以供试材料的cDNA为模板, 以基因和基因(表1)为内参基因, 使用SYBR Green ProHS预混型qPCR试剂盒(含ROX,艾瑞科, 中国), 按其说明书配制反应体系, 在7500型实时荧光定量PCR仪(ABI, USA)进行RT-qPCR。每个样品设置3次技术重复和生物学重复, 以DEPC处理的H2O作为对照, 采用2–ΔΔCt算法计算基因的相对表达量, 使用统计软件SPSS 20对基因表达差异进行显著性分析。
2 结果与分析
2.1 ScPVA12基因的克隆与生物信息学分析
经PCR扩增和测序, 本研究从ROC22中克隆了1条开放读码框(open reading frame, ORF)长度为735 bp的PVA12基因, 编码长度为244 aa的蛋白。将该基因命名为并提交GenBank, 登录号为OP828637。ProtParam分析表明, ScPVA12蛋白分子式为C1178H1924N328O365S9, 分子量为26,810.75 kD; 该蛋白的不稳定指数为63.22, 为不稳定蛋白; 脂溶指数为83.48, 且总体平均亲水性为负值, 因此该蛋白可能是一种亲水性脂溶蛋白; ScPVA12的二级结构预测显示, 该蛋白无规则卷曲、α螺旋和延伸链占比分别为50%、27.87%和22.13%; 信号肽预测表明, ScPVA12蛋白无信号肽, 属于非分泌蛋白; 跨膜结构域预测分析表明, ScPVA12在C端具有一个TMD。ScPVA12与拟南芥AtVAP27-3 (Locus: AT2G45140)氨基酸序列比对结果表明他们的氨基酸序列长度相近且序列保守。结构域分析结果表明ScPVA12与AtVAP27-3都具有VAP27蛋白家族典型的MSD、CCD, 以及C端TMD, 表明ScPVA12属于VAP27蛋白家族并且是VAP27-3的同源物(图1)。
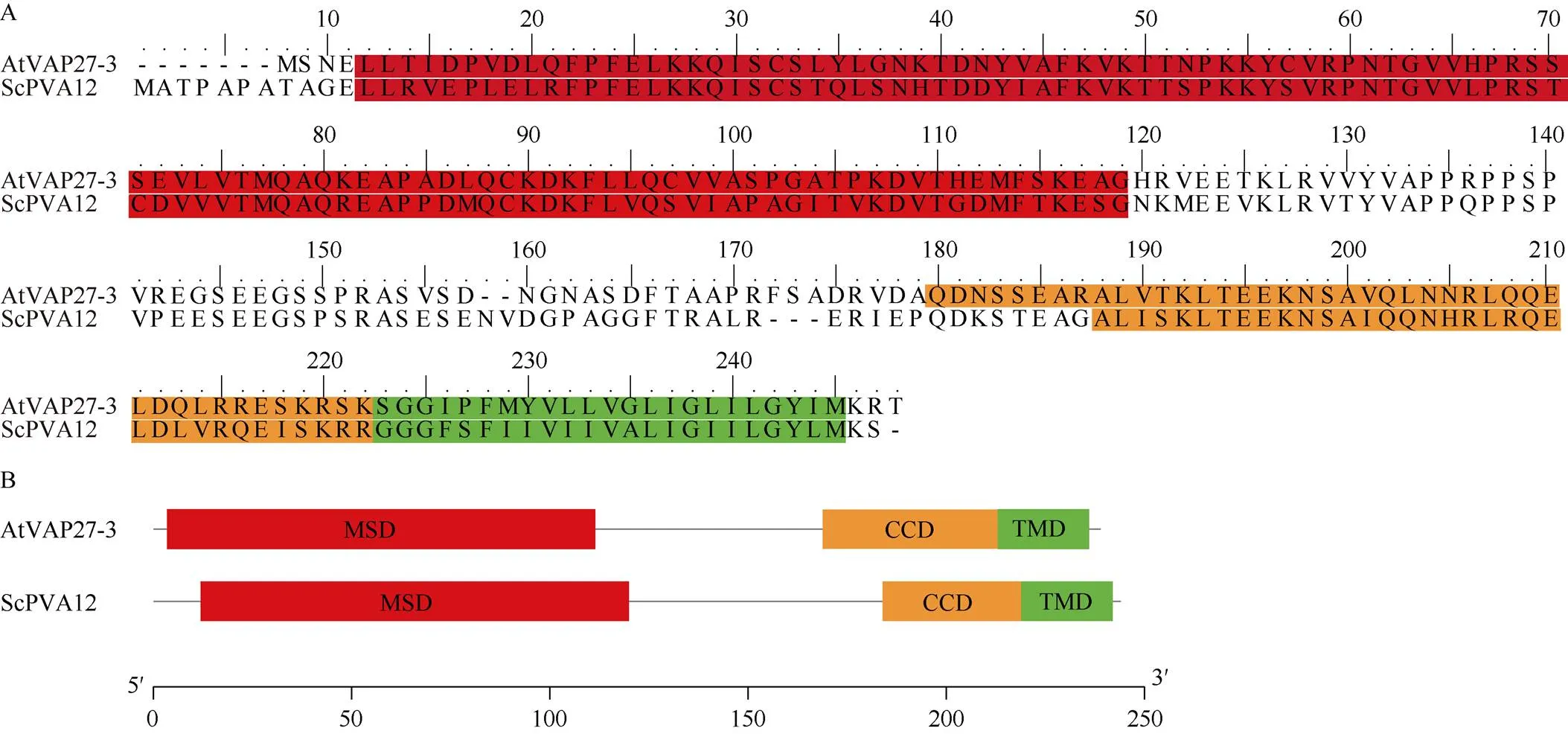
图1 ScPVA12蛋白结构域分析
A: ScPVA12与AtVAP27-3氨基酸比对结果(红色标记为MSD, 黄色标记为CCD, 蓝色标记为TMD)。B: ScPVA12与AtVAP27-3结构域示意图(标尺为250个氨基酸残基)。
A: the amino acid comparison between ScPVA12 and AtVAP27-3 (The red marks indicate MSD, the orange marks indicate CCD, and the green marks indicate TMD. B: the schematic diagram of ScPVA12 and AtVAP27-3 domain (Ruler: 250 amino acids).
2.3 ScPVA12的氨基酸同源性分析和系统进化树分析
通过NCBI网站的Blastp搜索PVA12的单子叶物种同源序列发现, ScPVA12蛋白与高粱(, XP_002445075.1)、南荻(, CAD6266147.1)、玉米(, NP_ 001130606.1)、黍(, RLN03983.1)、谷子(, XP_004972806.1)、二穗短柄草(, XP_003573476.1)、野生二粒小麦(, XP_037458000.1)中的PVA12蛋白相似度分别为: 96%、97%、95%、93%、91%、86%、86%, 表明PVA12蛋白具有高度保守性(图2)。
在NCBI网站检索不同物种的已注释的PVA12同源序列, 利用MEGA软件的Neighbor Joining算法构建PVA12的系统进化树(图3)。结果表明, 单子叶植物南荻()、玉米()、高粱()、弯叶画眉草()、黍()、沼生菰()、稻()、短花药野生稻()、二穗短柄草()、野生二粒小麦植物南荻()、玉米()、高粱()、弯叶画眉草()、黍()、沼生菰()、稻(Group)、短花药野生稻()、二穗短柄草()、野生二粒小麦()和大麦()组成群I, 双子叶植物番木瓜()、拟南芥()、大豆()、可可()、温州蜜柑()和开心果()组成群II。在单子叶植物群中又可以分为2个亚群, 分别为C4植物亚群(亚群I-1)和C3植物亚群(亚群II-2)。这说明在遗传进化上, PVA12蛋白在单子叶和双子叶植物之间, 以及C3和C4植物之间存在明显的分化。

图2 甘蔗ScPVA12与其他单子叶植物PVA12蛋白的氨基酸序列比对
高粱: SbPVA12 (XP_002445075.1); 南荻: MlPVA12 (CAD6266147.1); 玉米: ZmPVA12 (NP_001130606.1); 黍: PmPVA12(RLN03983.1); 谷子: SiPVA12 (XP_004972806.1); 二穗短柄草: BdPVA12 (XP_003573476.1); 野生二粒小麦: TdPVA12 (XP_037458000.1)。
: SbPVA12 (XP_002445075.1);: MlPVA12 (CAD6266147.1);: ZmPVA12 (NP_001130606.1);: PmPVA12 (RLN03983.1);: SiPVA12 (XP_004972806.1);: BdPVA12 (XP_003573476.1);: TdPVA12 (XP_037458000.1).

图3 ScPVA12与其他物种PVA12蛋白的系统进化树分析
群I为单子叶植物群, 其中亚群I-1为C4植物, 亚群群I-2为C3植物; 群II为双子叶植物群。
Group I is monocot, in which I-1 is C4plant subgroup and I-2 is C3plant subgroup. Group II is a dicotyledonous group.
2.4 ScPVA12的亚细胞定位
前人报道PVA12为内质网蛋白[37,53], 因此以HDEL-RFP作为内质网标记进行亚细胞定位试验。试验结果显示, ScPVA12-YFP融合蛋白的黄色荧光信号(绿色伪彩)与HDEL-RFP融合蛋白的红色荧光信号重合, 表明ScPVA12定位于内质网(图4-A), 与文献报道的酵母Scs2p和拟南芥AtPVA12的亚细胞定位结果相同[37,53]。将SCMV-P3N-PIPO-CFP、ScPVA12-YFP以及HDEL-RFP蛋白在健康的烟草叶片中表达, ScPVA12-YFP与SCMV-P3N-PIPO-CFP融合蛋白的青色荧光蛋白信号重合, 表明SCMV- P3N-PIPO和ScPVA12具有共同定位。将3种融合蛋白的荧光信号进行两两组合并使用伪彩进行观察(图4-B), ScPVA12-YFP (绿色伪彩)和HDEL-RFP (红色)组合的荧光信号重合, 表明ScPVA12定位在内质网; ScPVA12-YFP (红色伪彩)和P3N-PIPO-CFP (绿色伪彩)组合的荧光信号重合, 表明ScPVA12与P3N-PIPO具有共同定位; HDEL-RFP (红色)和P3N-PIPO-CFP (绿色伪彩)组合的荧光信号重合, 表明在P3N-PIPO与ScPVA12在内质网上具有共定位。如图4-B Merge中白色箭头所示, SCMV-P3N-PIPO- CFP、ScPVA12-YFP和HDEL-RFP存在共定位。部分SCMV-P3N-PIPO可能受ScPVA12的影响改变了其原本在质膜[36]上的定位。

图4 ScPVA12-YFP在本氏烟表皮细胞中的定位
A: ScPVA12-YFP亚细胞定位; B: ScPVA12-YFP和SCMV-P3N-PIPO-CFP亚细胞共定位。将3种荧光蛋白信号进行两两组合并使用伪彩进行观察共定位情况, 组合分别为ScPVA12-YFP (绿色伪彩)+HDEL-RFP (红色)、ScPVA12-YFP (红色伪彩)+P3N-PIPO-CFP (绿色伪彩)以及HDEL-RFP (红色)+P3N-PIPO-CFP (绿色伪彩)。标尺为25 μm。白色箭头标记共定位位置。
A: the subcellular localization of ScPVA12-YFP; B: the subcellular colocalization of ScPVA12-YFP with SCMV-P3N-PIPO-CFP. Pairwise combination of three fluorescent protein signals and observation of colocalization presented by using pseudo-color, and the combinations were ScPVA12-YFP (green pseudo-color)+HDEL-RFP (red pseudo-color), ScPVA12-YFP (red pseudo-color)+P3N-PIPO-CFP (green pseudo-color) and HDEL-RFP (red)+P3N-PIPO-CFP (green pseudo-color). Bar: 25 μm. The white arrow marks the location of colocalization.
2.5 ScPVA12与SCMV-P3N-PIPO的互作验证
Y2H试验结果表明, 试验组pPR3-ScPVA12/ pBT3-SET-P3N-PIPO与阳性对照pTSU2-APP/ pNubG-Fe65结果相似, 且在滴加X-Gal 8 h后肉眼可见菌落变蓝, 表明ScPVA12与SCMV-P3N-PIPO互作(图5); BiFC试验结果表明, 共注射的组合ScPVA12-YC与YN-P3N-PIPO、组合YN-ScPVA12与P3N-PIPO-YC都分别产生黄色荧光信号(图6), 进一步证明了SCMV-P3N-PIPO与ScPVA12互作。此外, 在BiFC试验中, 2种蛋白互作产生的荧光信号具有明显的点状结构。
2.6 ScPVA12基因的组织特异性表达及应答SCMV侵染的表达模式
荧光定量PCR结果表明,基因在不同组织的表达量具有差异性。基因在叶片中的表达量高于茎和根, 其中正七叶的表达量最高; 正一叶和心叶, 第3节间、第8节间和根的表达量较低, 其中第8节间的表达量最低(图7)。
使用基因特异性引物(表1)检测SCMV接种甘蔗ROC22组培苗叶片, 扩增出目的片段, 表明接种成功。荧光定量PCR结果表明, SCMV侵染对基因表达影响显著, 与ROC22叶片接种SCMV 0 h的对照相比,表达量总体上呈现先下降再上升的趋势。接种SCMV 1 h、2 h、4 h、8 h、12 h、24 h、48 h、72 h时,表达量下调, 在接种1 h时表达量显著下调; 在接种5 d、8 d、14 d时,表达量逐渐恢复到SCMV侵染前的表达水平(图8)。

图5 Y2H检测ScPVA12与SCMV-P3N-PIPO的互作
pTSU2-APP/pNubG-Fe65为阳性对照, pPR3-N/pNubG-Fe65为阴性对照。DDO+X-Gal: 添加了5-溴-4-氯-3-吲哚-β-D-半乳糖苷的缺少亮氨酸(Leu)和色氨酸(Trp)的酵母合成限定基本培养基; QDO+X-Gal: 添加了X-Gal的缺少亮氨酸(Leu)、色氨酸(Trp)、组氨酸(His)和腺嘌呤(Ade)的酵母合成限定基本培养基。
The positive and negative controls are yeast cotransformants with pNubG-Fe65 plus pTSU2-APP and pNubG-Fe65 plus pPR3-N, respectively. DDO+X-Gal: synthetic defined yeast minimal medium lacking Leu and Trp with the treatment of 5-Bromo-4Chlor-3-Indoly1 β-D-Galactopyranoside; QDO+X-Gal: synthetic defined yeast minimal medium lacking Leu, Trp, His, and Ade but plus the X-Gal.

图6 BiFC检测ScPVA12与SCMV-P3N-PIPO的互作
A: YN融合于ScPVA12的N末端, YC融合于SCMV-P3N-PIPO的C末端; B: YC融合于ScPVA12的C末端, YN融合于SCMV-P3N-PIPO的N末端。将YN-ScPVA12和P3N-PIPO-YC (A)、YN-P3N-PIPO和ScPVA12-YC (B)分别共注射到本氏烟叶片中进行瞬时表达, 48 h后激光共聚焦观察。标尺为25 μm。白色箭头标记呈点状结构的荧光信号。
A: the N-terminal half of YFP was fused to the N-terminal of ScPVA12 to generate YN-ScPVA12, while the C-terminal half of YFP was fused to the C-terminal of SCMV-P3N-PIPO to generate P3N-PIPO-YC; B: the C-terminal half of YFP was fused to the C-terminal of ScPVA12 to generate ScPVA12-YC, while the N-terminal half of YFP was fused to the N-terminal of SCMV-P3N-PIPO to generate YN-P3N-PIPO. Plasmids combination of YN-ScPVA12 plus P3N-PIPO-YC (A), YN-P3N-PIPO plus ScPVA12-YC (B) were individually co-injected into N. benthamiana leaves for transient expression. The fluorescent signal was monitored by confocal microscopy at 48 hours post infiltration. Bar: 25 μm. The white arrow marks the fluorescent signal with a dotted structure.

图7 ScPVA12基因在甘蔗不同组织中的表达模式
误差线为每组处理的标准误差(= 3)。柱上不同的小写字母表示在< 0.05差异显著。
The error bars represent the standard error of each treating group (= 3). Bars super-scripted by different lowercase letters are significantly different at< 0.05.

图8 ScPVA12基因应答SCMV侵染的表达模式
误差线为每组处理的标准误差(= 3)。柱上不同的小写字母表示在< 0.05差异显著。
The error bars represent the standard error of each treating group (= 3). Bars super-scripted by different lowercase letters are significantly different at< 0.05.
3 讨论
植物病毒结构简单, 必须依赖于与寄主因子的互作才能建立系统性侵染[24,35,54]。本研究的Y2H和BiFC试验表明ScPVA12与SCMV-P3N-PIPO互作, 说明ScPVA12参与了SCMV的侵染进程。病毒在寄主体内的复制是其建立系统性侵染的关键环节之一,寄主体内的脂质和亚细胞膜对病毒VRC的组装和复制至关重要[53,55]。病毒VRC的组装和积累依赖于细胞器膜上的蛋白质和脂类[56-59]。甾醇是重要的膜组分[60], 研究表明, 在酵母中甾醇特异在病毒复制位点富集, 并能刺激病毒的复制与积累[53]。内质网是蛋白质折叠加工和脂质合成的场所, 病毒可以在内质网上利用脂质进行VRC的组装[53], 有文献报道内质网是侵染玉米的SCMV的重要复制场所[61]。甾醇的转运主要通过氧化甾醇结合蛋白相关蛋白(oxysterol-binding protein (OSBP)-related proteins, ORPs)进行[37,56-59,62]。ORP是重要的脂质转运蛋白(lipid transfer protein, LTP), 通过其特有的ORD结构域招募和结合甾醇类脂类[37,56-59,62]。ORP3a能够特异结合谷甾醇[37,53], 而谷甾醇是质膜和PD的主要甾醇组分[60]。VAP27蛋白通过与ORP3a互作将后者定位于内质网并招募甾醇, 参与脂类运输和代谢, 阻碍VAP27与ORP3a的互作, ORP3a失去内质网定位[37]。因此, VAP27蛋白被认为是VRC在膜上组装的锚定蛋白, 通过间接招募甾醇促进病毒复制和侵染[53,63]。为此, 我们推测SCMV-P3N-PIPO与ScPVA12互作对SCMV侵染具有重要的生物学意义, 一方面通过ScPVA12与ORP3a互作间接招募谷甾醇,便于VRC的形成, 促进病毒复制; 另一方面, 将SCMV-P3N-PIPO定位于内质网, 便于其在内质网内进一步的正确折叠修饰并通过细胞质被膜复合体II (coat protein complex II, COPII)途径进行胞内转运。
膜结合位点(membrane contact site, MCS)是重要的亚细胞结构, MCS在脂质翻转和转运中具有重要作用, 除了参与病毒的复制, 还参与了病毒的胞间移动[62,64]。内质网膜和质膜之间可以形成EPCS, VAP27是EPCS的重要组分, Wang等认为VAP27与ORP互作形成的复合体介导了氧化甾醇的转运[37,62-64]。病毒的胞间移动是以囊泡形式进行的[65-66]。SCMV-P3N-PIPO与ScPVA12互作, 很可能通过后者与ORP3a互作赋予囊泡特殊的甾醇组分并使病毒囊泡定位于EPCS或PD。本课题组前期研究表明SCMV-P3N-PIPO定位在质膜和PD[36], 本研究的共定位试验表明SCMV-P3N-PIPO、ScPVA12和HDEL-RFP可以共定位(图4), 且在BiFC试验中SCMV-P3N-PIPO和ScPVA12互作的荧光信号呈现点状结构(图6), 说明它们可能定位于EPCS。有文献表明EPCS与PD共定位[43], PD富含鞘脂和甾醇[60], 而携带与PD相同的脂类的囊泡则更容易通过PD移动到相邻细胞。明确SCMV-P3N-PIPO和ScPVA12互作的EPCS定位对于深入研究其在病毒侵染中的作用具有重要生物学意义, 因此后续我们将克隆EPCS的标记蛋白NET3C的编码基因, 验证ScPVA12与SCMV-P3N-PIPO是否互作于在EPCS。
病毒在寄主细胞中的复制会引起植物的防御反应, 这可能会影响寄主正常的代谢和激素信号, 并导致发育和形态异常, 甚至导致局部和系统性细胞死亡反应[67-68]。前人的试验表明PVA12具有促进病毒组装VRC的作用[53], 但是在本研究的应答SCMV侵染的表达模式分析结果显示在病毒侵染初期下调表达, 推测可能是植物通过抑制表达的防御反应来减缓病毒在体内的复制, 以达到自我保护的目的。在SCMV侵染后期,又逐渐恢复至未侵染前的表达水平, 这可能是由于甘蔗是SCMV的天然寄主, SCMV建立系统性侵染之后, SCMV不再依赖或利用ScPVA12。目前有关植物VAP27蛋白功能及其应答病毒侵染机制的研究非常有限,在病毒胁迫下的表达水平变化的根本原因仍需进一步明确。后继我们将开展ScPVA12及其蛋白家族的相关研究, 克隆甘蔗的ORP3a并验证其与PVA12的互作, 进一步明确ScPVA12在SCMV侵染甘蔗中的作用。
4 结论
的ORF长度为735 bp, 编码244 aa。亚细胞定位试验表明, ScPVA12蛋白主要定位在细胞内质网; Y2H和BiFC验证表明ScPVA12与SCMV-P3N-PIPO存在互作关系; 共定位试验表明ScPVA12与SCMV-P3N-PIPO在内质网存在共定位; RT-qPCR分析显示,基因在各组织中均表达, 其中正七叶表达量最高, 第8节间表达量最低; 在SCMV胁迫下早期下调表达, 后期逐步恢复正常表达水平。
[1] 刘晓雪, 王新超. 2017/18榨季中国食糖生产形势分析与2018/19榨季展望. 农业展望, 2018, 14(11): 40–46. Liu X X, Wang X C. Domestic sugar production situation in 2017/18 crushing season and its prospect for 2018/19 crushing season., 2018, 14(11): 40–46 (in Chinese with English abstract).
[2] 王明强, 李文凤, 黄应昆, 王晓燕, 卢文洁, 罗志明. 我国大陆蔗区发生的甘蔗病毒病及防控对策. 中国糖料, 2010, (4): 55–58. Wang M Q, Li Y P, Huang Y K, Wang X Y, Lu W J, Luo Z M. Occurrence and controlling strategies on sugarcane viral diseases in Chinese mainland., 2010, (4): 55–58 (in Chinese with English abstract).
[3] 刘燕群, 李玉萍, 梁伟红, 宋启道, 秦小立, 叶露. 国外甘蔗产业发展现状. 世界农业, 2015, (8): 147–152. Liu Y Q, Li Y P, Liang W H, Song Q D, Qin X L, Ye L. Current status and development of the abroad sugarcane industry., 2015, (8): 147–152 (in Chinese with English abstract).
[4] 李明, 田洪春, 黄智刚. 我国甘蔗产业发展现状研究. 中国糖料, 2017, 39(1): 67–70. Li M, Tian H C, Huang Z G. Research on the development status of sugarcane industry in China., 39(1): 67–70 (in Chinese with English abstract).
[5] 翁卓, 黄寒. 中国制糖产业竞争力对比与政策建议—基于对巴西、印度、泰国考察的比较. 甘蔗糖业, 2015, (4): 65–72. Weng Z, Huang H. Comparative analysis on China’s sugar industry competitiveness: based on the comparison of Brazil, India and Thailand sugar industry., 2015, (4): 65–72 (in Chinese with English abstract).
[6] 黄应昆, 李文风, 卢文洁, 罗志明. 云南蔗区甘蔗花叶病流行原因及控制对策. 云南农业大学学报, 2007, 22: 935–938. Huang Y K, Li W F, Lu W J, Luo Z M. The Causes of sugarcane mosaic disease epidemic in Yunnan sugarcane area and the control strategy., 2007, 22: 935–938 (in Chinese with English abstract).
[7] 颜梅新, 黄伟华, 邓展云, 韦金菊. 广西甘蔗花叶病SCMV调查初报. 中国糖料, 2012, (1): 50–51. Yan M X, Huang W H, Deng Z Y, Wei J J. Investigation ofinfecting sugarcane in Guangxi., 2012, (1): 50–51 (in Chinese with English abstract).
[8] 熊国如, 李增平, 赵婷婷, 蔡文伟, 王俊刚, 王文治, 冯翠莲, 张雨良, 张树珍. 海南蔗区甘蔗病害种类及发生情况. 热带作物学报, 2010, 31: 1588–1595. Xiong G R, Li Z P, Zhao T T, Cai W W, Wang J G, Wang W Z, Feng C L, Zhang Y L, Zhang S Z. Primary Investigation to Sugarcane on the Diseases in Hainan Province., 2010, 31: 1588–1595 (in Chinese with English abstract).
[9] 蒋军喜, 谢艳, 阙海勇. 江西甘蔗花叶病病原的分子鉴定. 植物病理学报, 2009, 39: 203–206. Jiang J X, Xie Y, Que H Y. Molecular identification of the pathogen of sugarcane mosaic disease in Jiangxi province., 2009, 39: 203–206 (in Chinese with English abstract).
[10] 梁姗姗, 罗群, 陈如凯, 高三基. 引起甘蔗花叶病的病原分子生物学进展. 植物保护学报, 2017, 44: 363–370. Liang S S, Luo Q, Chen R K, Gao S J. Advances in researches on molecular biology of viruses causing sugarcane mosaic., 2017, 44: 363–370 (in Chinese with English abstract).
[11] 冯小艳, 沈林波, 王文治, 杨本鹏, 王勤南, 周峰, 王俊刚, 熊国如, 张树珍. 中国甘蔗主要杂交亲本病毒性病害的分子鉴定. 分子植物育种, 2018, 16: 6729–6737. Feng X Y, Shen L B, Wang W Z, Yang B P, Wang Q N, Zhou F, Wang J G, Xiong G R, Zhang S Z. Molecular identification of viral diseases in major sugarcane hybrid parents in China., 2018, 16: 6729–6737 (in Chinese with English abstract).
[12] 周国辉, 许东林, 沈万宽. 甘蔗重要病害研究及防治策略. 甘蔗糖业, 2005, (1): 11–16. Zhou G H, Xu D L, Shen W K. On sugarcane major diseases and their controlling., 2005, (1): 11–16 (in Chinese).
[13] 周丰静, 黄诚华, 李正文, 商显坤, 黄伟华, 潘雪红, 魏吉利, 林善海. 广西蔗区甘蔗花叶病病毒种群分析. 南方农业学报, 2015, 46: 609–613. Zhou F J, Huang C H, Li Z W, Shang X S, Huang W H, Pan X H, Wei J L, Lin S H. Analysis of the virus population causingdisease in sugarcane growing area of Guangxi., 2015, 46: 609–613 (in Chinese with English abstract).
[14] 杨荣仲, 周会, 肖祎, 吕达, 廖红香, 陈道德, 刘昔辉, 雷敬超,林垠孚. 甘蔗主要亲本自然条件下抗甘蔗花叶病测定. 中国糖料, 2020, 42(2): 47–52.Yang R Z, Zhou H, Xiao Y, Lyu D, Liao H X, Chen D D, Liu X H, Lei J C, Lin Y F. Testing on sugarcane mosaic resistance of sugarcane major parents under field conditions., 2020, 42(2): 47–52 (in Chinese with English abstract).
[15] Ling H, Huang N, Wu Q, Su Y, Peng Q, Ahmed W, Gao S, Su W, Que Y, Xu L. Transcriptional insights into theinteraction., 2018, 11: 163–176.
[16] Akbar S, Yao W, Yu K, Qin L, Ruan M, Powell C A, Chen B, Zhang M. Photosynthetic characterization and expression profiles of sugarcane infected by(SCMV)., 2020, 150: 279–294.
[17] Wu L, Zu X, Wang S, Chen Y. Sugarcane mosaic virus: long history but still a threat to industry., 2012, 42: 74–78.
[18] 王文治, 马滋蔓, 张树珍, 杨本鹏, 蔡文伟, 顾丽红, 李娇. 甘蔗花叶病的基因工程研究. 生物技术通报, 2009, (1): 22–26. Wang W Z, Ma Z M, Zhang S Z, Yang B P, Cai W W, Gu L H, Li J. Research on genetic engineering of sugarcane mosaic disease., 2009, (1): 22–26 (in Chinese with English abstract).
[19] 张海, 刘淑娴, 杨宗桃, 王彤, 程光远, 商贺阳, 徐景升. 甘蔗PsbS亚基应答甘蔗花叶病毒侵染及其与6K2蛋白的互作研究. 作物学报, 2020, 46: 1534–1545. Zhang H, Liu S X, Yang Z T, Wang T, Cheng G Y, Shang H Y, Xu J S. Sugarcane PsbS subunit response toinfection and its interaction with 6K2 protein., 2020, 46: 1534–1545 (in Chinese with English abstract).
[20] 郑艳茹, 翟玉山, 邓宇晴, 成伟, 程光远, 杨永庆, 徐景升. 甘蔗花叶病毒(SCMV)种群结构分析. 福建农林大学学报(自然科学版), 2016, 45(2): 135–140.Zheng Y R, Zhai Y S, Deng Y Q, Cheng W, Cheng G Y, Yang Y Q, Xu J S. The population structure of(SCMV).(Nat Sci Edn), 2016, 45(2): 135–140 (in Chinese with English abstract).
[21] Yao W, Ruan M, Qin L, Yang C, Chen R, Chen B, Zhang M. Field performance of transgenic sugarcane lines resistant to., 2017, 8: 104.
[22] Filloux D, Fernandez E, Comstock J C, Mollov D, Roumagnac P, Rott P. Viral metagenomic-based screening of sugarcane from Florida reveals occurrence of six sugarcane-infecting viruses and high prevalence of., 2018, 102: 2317–2323.
[23] Yahaya A, Dangora D B, Kumar P L, Alegbejo M D, Gregg L, Alabi O J. Prevalence and genome characterization of field isolates of(SCMV) in Nigeria., 2019, 103: 818–824.
[24] 邓宇晴, 杨永庆, 翟玉山, 程光远, 彭磊, 郑艳茹, 林彦铨, 徐景升. 甘蔗花叶病毒福州分离物全基因组克隆及种群分析. 植物病理学报, 2016, 46: 775–782. Deng Y Q, Yang Y Q, Zhai Y S, Cheng G Y, Peng L, Zheng Y R, Lin Y Q, Xu J S. Genome cloning of twoisolates from Fuzhou and phylogenetic analysis of SCMV., 2016, 46: 775–782 (in Chinese with English abstract)
[25] Xu D L, Park J W, Mirkov T E, Zhou G H. Viruses causing mosaic disease in sugarcane and their genetic diversity in southern China., 2008, 153: 1031–1039.
[26] 沈林波, 吴楠楠, 冯小艳, 熊国如, 赵婷婷, 王文治, 王俊刚, 张树珍. 52个甘蔗品种在广西受病毒侵染情况. 热带作物学报, 2020, 41(1): 116–126. Shen L B, Wu N N, Feng X Y, Xiong G R, Zhao T T, Wang W Z, Wang J G, Zhang S Z. Virus infection situation of fifty-two sugarcane varieties in Guangxi., 2020, 41(1): 116–126 (in Chinese with English abstract).
[27] Olspert A, Chung B Y, Atkins J F, Carr J P, Firth A E. Transcriptional slippage in the positive-sense RNA virus family., 2015, 16: 995–1004.
[28] Wei T, Zhang C, Hong J, Xiong R, Kasschau K D, Zhou X, Carrington J C, Wang A. Formation of complexes at plasmodesmata for potyvirus intercellular movement is mediated by the viral protein P3N-PIPO., 2010, 6: e1000962.
[29] Chai M, Wu X, Liu J, Fang Y, Luan Y, Cui X, Zhou X, Wang A, Cheng X. P3N-PIPO interacts with P3 via the shared N-terminal domain to recruit viral replication vesicles for cell-to-cell movement., 2020, 94: e01898.
[30] Tilsner J, Linnik O, Louveaux M, Roberts I M, Chapman S N, Oparka K J. Replication and trafficking of a plant virus are coupled at the entrances of plasmodesmata., 2013, 201: 981–995.
[31] 胡帆, 雷荣, 廖晓兰. 植物病毒在细胞间转运的机理探讨. 生物学杂志, 2013, 30(6): 81–85. Hu F, Lei R, Liao X L. The mechanism of viral intracellular transportation in plant., 2013, 30(6): 81–85 (in Chinese with English abstract).
[32] Hillung J, Elena S F, Cuevas J M. Intra-specific variability and biological relevance of P3N-PIPO protein length in potyviruses., 2013, 13: 249.
[33] Lin W, Feng Z, Prasanth K R, Liu Y, Nagy P D. Dynamic interplay between the co-opted Fis1 mitochondrial fission protein and membrane contact site proteins in supporting tombusvirus replication., 2021, 17: e1009423.
[34] 崔晓艳, 陈新, 顾和平, 张红梅, 陈华涛, 袁星星. 马铃薯Y病毒属病毒P3和P3-PiPo蛋白功能研究进展. 微生物学通报, 2012, 39(1): 99–105. Cui X Y, Chen X, Gu H P, Zhang H M, Chen H T, Yuan X X. The functional characterization of-encoded P3 and P3-PiPo protein., 2012, 39(1): 99–105 (in Chinese with English abstract).
[35] Wang A. Dissecting the molecular network of virus-plant interactions: the complex roles of host factors., 2015, 53: 45–66.
[36] Cheng G, Dong M, Xu Q, Peng L, Yang Z, Wei T, Xu J. Dissecting the molecular mechanism of the subcellular localization and cell-to-cell movement of theP3N-PIPO., 2017, 7: 9868.
[37] Saravanan R S, Slabaugh E, Singh V R, Lapidus L J, Haas T, Brandizzi F. The targeting of the oxysterol-binding protein ORP3a to the endoplasmic reticulum relies on the plant VAP33 homolog PVA12., 2009, 58: 817–830.
[38] Laurent F, Labesse G, de Wit P. Molecular cloning and partial characterization of a plant VAP33 homologue with a major sperm protein domain., 2000, 270: 286–292.
[39] Pérez-Sancho J, Tilsner J, Samuels A L, Botella M A, Bayer E M, Rosado A. Stitching organelles: organization and function of specialized membrane contact sites in plants., 2016, 26: 705–717.
[40] Takáč T, Šamajová O, Vadovič P, Pechan T, Šamaj J. Shot-gun proteomic analysis on roots ofmutants suggesting the involvement of PLDα1 in mitochondrial protein import, vesicular trafficking and glucosinolate biosynthesis., 2018, 20: 82.
[41] Sutter J U, Campanoni P, Blatt M R, Paneque M. Setting SNAREs in a different wood., 2006, 7: 627–638.
[42] Ichikawa M, Nakai Y, Arima K, Nishiyama S, Hirano T, Sato M H. A VAMP-associated protein, PVA31 is involved in leaf senescence in., 2015, 10: e990847.
[43] Wang P, Richardson C, Hawkins T J, Sparkes I, Hawes C, Hussey P J. Plant VAP27 proteins: domain characterization, intracellular localization and role in plant development., 2016, 210: 1311–1326.
[44] Loewen C J, Levine T P. A highly conserved binding site in vesicle-associated membrane protein-associated protein (VAP) for the FFAT motif of lipid-binding proteins., 2005, 280: 14097–14104.
[45] Murphy S E, Levine T P. VAP, a versatile access point for the endoplasmic reticulum: review and analysis of FFAT-like motifs in the VAPome., 2016, 1861: 952–961.
[46] D’Ippólito S, Arias L A, Casalongué C A, Pagnussat G C, Fiol D F. The DC1-domain protein VACUOLELESS GAMETOPHYTES is essential for development of female and male gametophytes in., 2017, 90: 261–275.
[47] 金红敏, 李立新. 拟南芥SNARE因子在膜泡运输中的功能. 植物学报, 2010, 45: 479–491. Jin H M, Li L X. Role ofSNARE proteins in vesicle trafficking., 2010, 45: 479–491 (in Chinese with English abstract).
[48] Stefano G, Renna L, Wormsbaecher C, Gamble J, Zienkiewicz K, Brandizzi F. Plant endocytosis requires the ER membrane- anchored proteins VAP27-1 and VAP27-3., 2018, 23: 2299–2307.
[49] Wang P, Pleskot R, Zang J, Winkler J, Wang J, Yperman K, Zhang T, Wang K, Gong J, Guan Y, Richardson C, Duckney P, Vandorpe M, Mylle E, Fiserova J, Van Damme D, Hussey P J. Plant AtEH/Pan1 proteins drive autophagosome formation at ER-PM contact sites with actin and endocytic machinery., 2019, 10: 5132.
[50] Wang P, Hawkins T J, Richardson C, Cummins I, Deeks M J, Sparkes I, Hawes C, Hussey P J. The plant cytoskeleton, NET3C, and VAP27 mediate the link between the plasma membrane and endoplasmic reticulum., 2014, 24: 1397–1405.
[51] Zhang Z, Thomma B P. Structure-function aspects of extracellular leucine-rich repeat-containing cell surface receptors in plants., 2013, 55: 1212–1223.
[52] Carette J E, Verver J, Martens J, van Kampen T, Wellink J, van Kammen A. Characterization of plant proteins that interact with‘60K’ protein in the yeast two-hybrid system., 2002, 83: 885–893.
[53] Barajas D, Xu K, de Castro Martín I F, Sasvari Z, Brandizzi F, Risco C, Nagy P D. Co-opted oxysterol-binding ORP and VAP proteins channel sterols to RNA virus replication sites via membrane contact sites., 2014, 10: e1004388.
[54] Lin L, Luo Z, Yan F, Lu Y, Zheng H, Chen J. Interaction between potyvirus P3 and ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO) of host plants., 2011, 43: 90–92.
[55] 王倩. 调控黄瓜花叶病毒复制的寄主脂质因子的研究. 浙江理工大学硕士学位论文, 浙江杭州, 2019. Wang Q. Study on the Regulation of Host Lipid Factors in CMV Replication. MS Thesis of Zhejiang Sci-Tech University, Hangzhou, Zhejiang, China, 2019 (in Chinese with English abstract).
[56] Xu K, Nagy P D. Enrichment of phosphatidylethanolamine in viral replication compartments via co-opting the endosomal Rab5 small GTPase by a positive-strand RNA virus., 2016, 14: e2000128.
[57] Chuang C, Barajas D, Qin J, Nagy P D. Inactivation of the host lipin gene accelerates RNA virus replication through viral exploitation of the expanded endoplasmic reticulum membrane., 2014, 10: e1003944.
[58] Sharma M, Sasvari Z, Nagy P D. Inhibition of sterol biosynthesis reduces tombusvirus replication in yeast and plants., 2010, 84: 2270–2281.
[59] Xu K, Nagy P D. RNA virus replication depends on enrichment of phosphatidylethanolamine at replication sites in subcellular membranes., 2015, 112: E1782–E1791.
[60] Grison M S, Brocard L, Fouillen L, Nicolas W, Wewer V, Dörmann P, Nacir H, Benitez-Alfonso Y, Claverol S, Germain V, Boutté Y, Mongrand S, Bayer E M. Specific membrane lipid composition is important for plasmodesmata function in., 2015, 27: 1228–1250.
[61] Xie J, Jiang T, Li Z, Li X, Fan Z, Zhou T.remodels multiple intracellular organelles to form genomic RNA replication sites., 2021, 166: 1921–1930.
[62] 邹文娇, 葛磊, 予茜. 氧化甾醇结合蛋白相关蛋白家族的研究进展. 植物学报, 2021, 56: 627–640. Zou W J, Ge L, Yu Q. Research advances in oxysterol-binding protein-related proteins., 2021, 56: 627–640 (in Chinese with English abstract).
[63] Wang P, Hawes C, Hussey P J. Plant endoplasmic reticulum- plasma membrane contact sites., 2017, 22: 289–297.
[64] Levy A, Tilsner J. Creating contacts between replication and movement at plasmodesmata: a role for membrane contact sites in plant virus infections?, 2020, 11: 862.
[65] Zhang H, Cheng G, Yang Z, Wang T, Xu J. Identification of sugarcane host factors interacting with the 6K2 protein of the., 2019, 20: 3867.
[66] Cheng G, Yang Z, Zhang H, Zhang J, Xu J. Remorin interacting with PCaP1 impairsintercellular movement but is antagonised by VPg., 2020, 225: 2122–2139.
[67] Hyodo K, Okuno T. Pathogenesis mediated by proviral host factors involved in translation and replication of plant positive-strand RNA viruses., 2016, 17: 11–18.
[68] Addy H S, Nurmalasari, Wahyudi A H S, Sholeh A, Anugrah C, Iriyanto F E S, Darmanto W, Sugiharto B. Detection and response of sugarcane against the infection of(SCMV) in Indonesia., 2017, 7: 50.
Interaction of sugarcane VAMP associated protein ScPVA12 with SCMV P3N-PIPO
YU Quan-Xin1, YANG Zong-Tao1, ZHANG Hai1, CHENG Guang-Yuan1, ZHOU Ying-Shuan1, JIAO Wen-Di1, ZENG Kang1, LUO Ting-Xu1, HUANG Guo-Qiang1, ZHANG Mu-Qing2,*, and XU Jing-Sheng1,*
1Fujian Agriculture and Forestry University / Key Laboratory of Sugarcane Biology and Genetic Breeding, Ministry of Agriculture and Rural Affairs / National Engineering Research Center for Sugarcane / Key Laboratory of Ministry of Education for Genetics, Breeding and Multiple Utilization of Crops, Fuzhou 350002, Fujian, China;2Guangxi Key Laboratory of Sugarcane Biology, Guangxi University, Nanning 530004, Guangxi, China
PVA12 (plant vesicle-associated membrane protein (VAMP)-associated proteins homolog 12) is a member of the VAP27 family of proteins that mediate endoplasmic reticulum (ER) vesicle transport and membrane fusion in cells. Sugarcane (spp. hybrid) PVA12 responding to(SCMV) infection has not been reported. In the present study, the coding gene of PVA12 was cloned from sugarcane cultivar ROC22 and designated as. The open reading frame (ORF) ofwas 735 bp in length, which encoded a protein with 244 amino acids. Bioinformatics analysis showed that ScPVA12 was an unstable hydrophilic liposoluble protein with a transmembrane at the C-terminal domain. The ratio of the random coil ranked the highest in the secondary structure. Phylogenetic tree analysis revealed that the ScPVA12 was differentiated in monocotyledons and dicotyledons. Yeast two-hybrid (Y2H) and bimolecular fluorescence complementation (BiFC) assays showed that ScPVA12 interacted with SCMV-P3N-PIPO. Subcellular localization experiments indicated that ScPVA12 was localized in the ER. Co-localization experiments demonstrated that ScPVA12 and SCMV-P3N-PIPO co-localized in the ER. Real-time quantitative PCR analysis showed thatgene was expressed in all sugarcane tissues, with the lowest expression level in the eighth internode and the highest expression level in the 7th leaves. The relative expression level ofgene was significantly affected under the stress of SCMV.was down-regulated upon SCMV infection and then recovered to the regular level compared with the control.
PVA12;; P3N-PIPO; protein interaction
2022-10-28;
2023-02-10;
2023-02-28.
10.3724/SP.J.1006.2023.24244
通信作者(Corresponding authors):徐景升, E-mail: xujingsheng@126.com; 张木清, E-mail: zmuqing@163.com
E-mail: YuQuanxin_YQX@outlook.com
本研究由国家自然科学基金项目(31971991), 福建农林大学科技创新基金项目(CXZX2019132G)和广西甘蔗生物学重点实验室开放课题(GXKLSCB-202003)资助。
This study was supported by the National Natural Science Foundation of China (31971991), the Science and Technology Innovation Project of Fujian Agriculture and Forestry University (CXZX2019132G), and the Open Project Program of Guangxi Key Laboratory of Sugarcane Biology (GXKLSCB-202003).
URL: https://kns.cnki.net/kcms/detail//11.1809.S.20230227.1407.006.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
