OsIMA1增强水稻对镉逆境的适应性
2023-07-17彭凤路承凯梁岗
彭凤 路承凯 梁岗
摘 要: 铁(Fe)是植物生长发育所必需的营养元素而镉(Cd)是对植物有害的元素且对植物Fe和Cd的吸收存在拮抗作用。OsIMA是一类正调控水稻Fe吸收的一类小肽,其过表达可以促进Fe的积累。为探究OsIMA是否參与水稻对Cd胁迫的适应性,该研究以水稻为研究材料,利用荧光定量PCR分析了OsIMA基因的表达水平,通过遗传转化和CRISPR/Cas9基因编辑技术构建了OsIMA1过表达植物和ima1突变体植物,评估了OsIMA1过表达和突变体植物在Cd逆境条件下的株高,并利用电联耦合等离子体质谱法测量了根和地上部的Fe和Cd含量。结果表明:(1)Cd处理后,OsIMA1和OsIMA2的转录水平上调。(2)OsIMA1过表达植物比野生型植物对Cd胁迫更耐受。(3)ima1功能缺失突变体比野生型植物对Cd胁迫更敏感。(4)OsIMA1过表达植株根系的Cd含量较高,而ima1突变体植株地上部的Cd含量较高。综上所述,OsIMA1通过限制Cd从根向地上部的转运以增强水稻对Cd逆境的适应能力,该研究结果为定向培育耐Cd作物提供了理论参考。
关键词: 水稻, OsIMA, Cd, Fe, 拮抗作用
中图分类号: Q945文献标识码: A 文章编号: 1000-3142(2023)06-1097-08
OsIMA1 enhances tolerance to cadmium stress in rice
PENG Feng1,2 , LU Chengkai1,2, LIANG Gang1,2*
( 1. Key Laboratory of Tropical Plant Resources and Sustainable Use, Xishuangbanna Tropical Botanical Garden, Chinese Academy of
Sciences,
Kunming, 650223, China; 2. College of Life Sciences, University of Chinese Academy of Sciences, Beijing 100049, China )
Abstract: Iron (Fe) is crucial for the growth and development of plants and cadmium (Cd) is toxic to plants. There is an antagonistical mechanism between Fe and Cd uptake in plants. OsIMAs are a class of small peptides, and their overexpression improves Fe accumulation in rice. To explore the role of OsIMA genes in response to Cd stress, we analyzed the expression of two OsIMA genes by qRT-PCR, generated OsIMA1 overexpression plants and CRISPR/Cas9 edited ima1 mutants by genetic transformation, assessed the plant heights of OsIMA1 overexpressing plants and ima1 mutant plants under Cd stress, and measured the Fe and Cd concentration of root and shoot. The results were as follows: (1) Transcriptional levels both OsIMA1 and OsIMA2 were up-regulated by Cd treatment. (2) Overexpression of OsIMA1 gene improved the tolerance of plants to Cd stress. (3) The loss-of-function of OsIMA1 led to the higher sensitivity of plants to Cd stress. (4) OsIMA1 overexpressing plants accumulated more Cd in the root and the ima1 mutants accumulated more Cd in the shoot. To sum up, OsIMA1 improves Cd tolerance by restricting Cd translocation from root to shoot, which provides the theoretical reference for breeding the Cd-tolerant rice.
Key words: rice, OsIMA, cadmium, iron, antagonism
Cd是对所有生物都有害的重金属元素,极易在根系从土壤吸收的过程中与其他二价金属离子(如Fe和Zn)竞争。Cd还可以通过食物链在人体内富集,长期积累可导致骨质疏松症、癌症和肾功能障碍等多种疾病。随着工业化的快速发展以及化肥和农药的过度使用,土壤中不断增加的Cd污染已经严重威胁到粮食安全,增加了对人类健康的潜在风险,并造成了巨大的经济损失(Kirkham, 2006; Wei & Yang, 2010)。因此,研究植物吸收Cd的分子机理并筛选和培育耐Cd胁迫或低Cd累积的农作物,对人类健康和生态保护具有重要意义。
由于Cd和Fe具有相似的水合离子半径,Cd极易通过与Fe竞争并借助Fe离子通道或转运蛋白进入植物体内(Nightingale, 1959; Eide et al., 1996; Schutzendubel & Polle, 2002; Clemens, 2006)。Cd在植物体内的积累导致幼叶明显褪绿、坏死、生长受阻甚至死亡,这与典型的缺Fe症状相似(Das et al., 1997; Cohen et al., 1998; Yoshihara et al., 2006)。相关研究表明,Cd胁迫改变了细胞壁成分,增强了细胞壁对Fe的结合能力,导致Fe滞留在根的质外体中,并抑制Fe从根部到地上部的转运(Xu et al., 2015),这些症状可以通过给植物补给Fe元素得到改善(Bao et al., 2010; He et al., 2017; Huang et al., 2020)。Fe转运体(iron-regulated transporter 1,IRT1)参与Cd的吸收过程(Cohen et al., 1998; Korshunova et al., 1999; Vert et al., 2002; Fan et al., 2014; Mao et al., 2014; Guan et al., 2019; Zhu et al., 2020)。在擬南芥里,IRT1的表达直接受转录因子FIT(FER-like Iron deficient-induced transcription factor)和bHLH Ib亚族成员(bHLH38、bHLH39、bHLH100和bHLH101)的调控(Yuan et al., 2008)。FIT和bHLH38或bHLH39的共表达通过增加根部对Cd的螯合而减少地上部的Cd积累来提高植物对Cd胁迫的耐受性(Wu et al., 2012)。转录因子bHLH104正调控拟南芥Fe稳态,过表达bHLH104则可提高植物对Cd的耐受性(Zhang et al., 2015; Li et al., 2016; Yao et al., 2018)。BTS(BRUTUS)负调控Fe稳态,敲除BTS显著增强了拟南芥对Cd胁迫的耐受性,同时增加了Fe和Cd的积累(Kobayashi et al., 2013; Hindt et al., 2017; Zhu et al., 2020)。在水稻中,2个Fe转运蛋白OsNRAMP1(Natural Resistance-Associated Macrophage Protein 1)和OsIRT1可以介导根系从土壤中吸收Cd,并且过表达OsNRAMP1和OsIRT1可以增加水稻的Cd含量(Lee & An, 2009; Takahashi et al., 2019)。由于Fe和Cd之间存在拮抗作用,因此水稻缺Fe会显著增加根系对Cd的吸收,外源施加Fe则会降低水稻的Cd含量并减缓Cd毒害(Shao et al., 2007)。综上表明,改善Fe的积累和分配可以显著减轻Cd对植物的毒害。因此,全面了解Fe和Cd的拮抗作用有利于指导农业生产和降低Cd对植物的毒性。
小肽泛指小于100个氨基酸的蛋白质,具有短的氨基酸共有基序,参与植物生长发育和非生物等逆境的胁迫响应(Czyzewicz et al., 2013; Matsubayashi, 2014)。IMA(IRON MAN)多肽家族成员的碳末端包含一个17氨基酸的共有基序,在被子植物中高度保守,并且这17个氨基酸就足以代替其全长蛋白发挥作用(Grillet et al., 2018; Li et al., 2021)。近年来,小肽作为激素类信号分子受到广泛的关注,参与器官内通讯、植物发育和胁迫应答(Tavormina et al., 2015; Takahashi et al., 2019)。IMA可能介导了Fe从地上部到根部的长距离信号,参与Fe的吸收、转运和细胞内稳态(Grillet et al., 2018; Li et al., 2021)。Meng等(2022)研究表明,过表达IMA可以提高在拟南芥中对Cd胁迫的适应性。但是,目前关于IMA是否可以提高农作物对Cd胁迫的适应性尚不清楚。本研究探究了水稻OsIMA1调控Cd胁迫方面的作用,以期为未来利用分子育种提高水稻对Cd胁迫的耐受性提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为粳稻品种“日本晴”,种植地点为云南省西双版纳傣族自治州勐腊县勐仑镇中国科学院西双版纳热带植物园的作物保护与育种基地(101°19′ E、21°52′ N)。将水稻种子用双蒸水(ddH2O)浸泡24 h后均匀地散布在吸水滤纸中,在28 ℃人工温室黑暗放置3 d后,转移到1/2MS培养液中萌发生长7 d,转移至不含Cd或含25 μmol·L-1 氯化镉(CdCl2)的1/2MS水培液中培养7 d。每2 d换1次水培液。生长光周期为光16 h、暗8 h的循环。
1.2 转基因植株的构建
利用CRISPR-GE(http://skl.scau.edu.cn/)在线设计OsIMA1的编辑靶点。通过Overlapping PCR构建包含该靶点的sgRNA,并融合至OsU6a启动子的下游,然后将OsU6a-sgRNA克隆至携带Cas9的pMH-SA载体(Liang et al., 2016)。将表达载体转化至农杆菌EHA105菌株中用于水稻转化,并对转基因阳性苗进行PCR测序。对纯合植株进行扩繁,并进一步对T3代植物在进行PCR测序,获得纯合突变植株。
用玉米Ubiquitin启动子驱动OsIMA1全长CDS,获得OsIMA1过表达载体。将构建的质粒转化至农杆菌EHA105菌株中用于水稻转化。选用含潮霉素抗性的转基因植株进行基因表达水平检测,T3转基因植株用来试验分析。
1.3 qRT-PCR
利用水饱和酚法提取水稻根(root)或地上部(shoot)的总RNA。采用RT Primer Mix (oligo dT)和PrimeScript RT Enzyme Mix for qPCR (宝生物,日本)试剂盒将RNA反转录成cDNA,随后使用PrimeScriptTM RT试剂(Perfect Real Time) Kit (宝生物,日本)在Light-Cycler 480实时PCR仪(罗氏,瑞士)上进行qRT-PCR检测。每个基因的定量至少含3次生物学重复。以OsACTIN1和OsOBP作为内参对照,对样品进行归一化处理。
1.4 Fe和Cd含量测定
不同基因型的水稻材料于1/2MS营养液体萌发生长7 d后转移至不含Cd或含25 μmol·L-1 CdCl2的1/2MS水培液培养7 d。分别收取根和叶,并置于65 ℃烘箱干燥7 d。用高通量组织研磨仪打磨样品成粉末。每个样品称取500 mg,加入5 mL硝酸于185 ℃中消解3 h,加入2 mL高氯酸于220 ℃氧化30 min。利用美国Thermo SCIENTIFIC公司的电联耦合等离子体质谱仪(ICP-MS iCAP6300)测定样品元素含量。
1.5 统计分析
所有试驗数据均使用平均值±标准差(x±s)表示。每个试验至少有3次生物学重复。利用SPSS 17.0软件进行单因素方差分析(one-way variance, ANOVA) (P<0.05)。
2 结果与分析
2.1 Cd胁迫引起水稻OsIMA基因表达上调
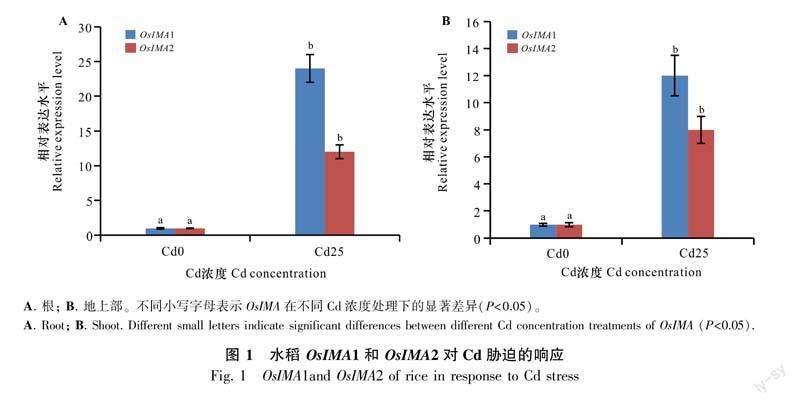
水稻基因组包含两个OsIMA基因(OsIMA1和OsIMA2)。近来的研究表明OsIMA1和OsIMA2受到缺Fe诱导(Kobayashi et al., 2021)。Cd胁迫会严重限制植物的生长发育,从而导致植物表现出缺Fe症状。为研究Cd胁迫是否能影响OsIMA1和OsIMA2的表达,对野生型(wild type,WT)水稻进行Cd处理后,检测了OsIMA1和OsIMA2的表达情况。将在正常培养液上生长7 d的幼苗转移至不含Cd(Cd0)或含25 μmol·L-1 CdCl2(Cd25)的培养液中生长7 d。提取根和地上部的RNA并利用荧光定量PCR检测OsIMA1和OsIMA2的表达水平。结果表明,在Cd胁迫条件下,无论在根还是地上部,OsIMA1和OsIMA2的表达均显著上调(图1)。
2.2 OsIMA1过表达植株对Cd耐受性更高
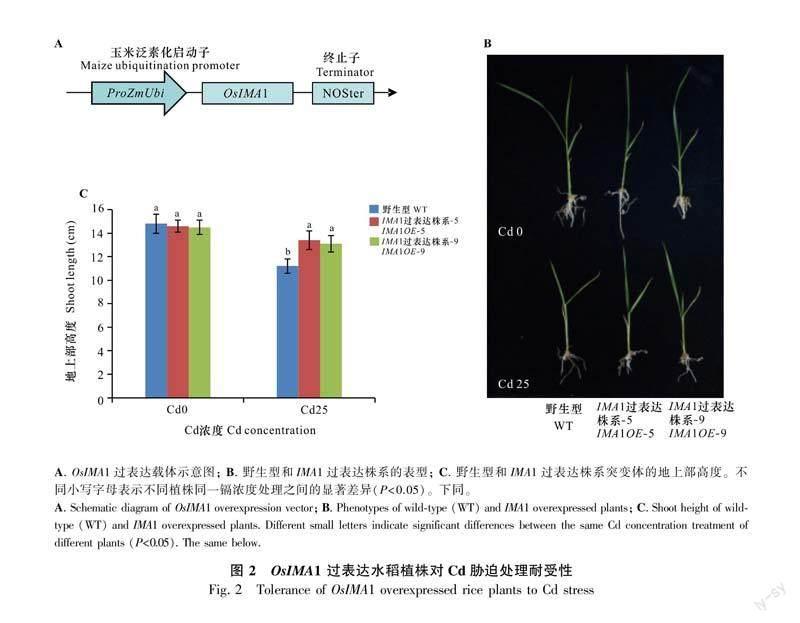
OsIMA1和OsIMA2属于同源蛋白且二者在Fe稳态方面的功能相近。考虑到二者的转录水平均受到Cd处理诱导,本研究接下来以OsIMA1作为代表进行研究。本研究利用玉米的Ubiquitin启动子驱动OsIMA1基因,并且转化了野生型水稻品种“日本晴”(图2:A)。通过定量PCR筛选获得了OsIMA1的过表达转基因水稻植株,评估OsIMA1过表达植株对Cd胁迫的适应能力。在正常水培溶液生长一周的幼苗分别转移到不含Cd (Cd0)或含25 μmol·L-1 CdCl2 (Cd25)的溶液再生长一周,观察和分析其表型。结果表明,在Cd0生长条件下,OsIMA1的过表达植株与野生型植株无明显区别,而在Cd25处理条件下,过表达植株的株高显著高于野生型植株(图2:B,C)。综上表明,OsIMA1过表达增强了植物对Cd胁迫的适应性。
2.3 OsIMA1的功能缺失突变体对Cd胁迫更敏感
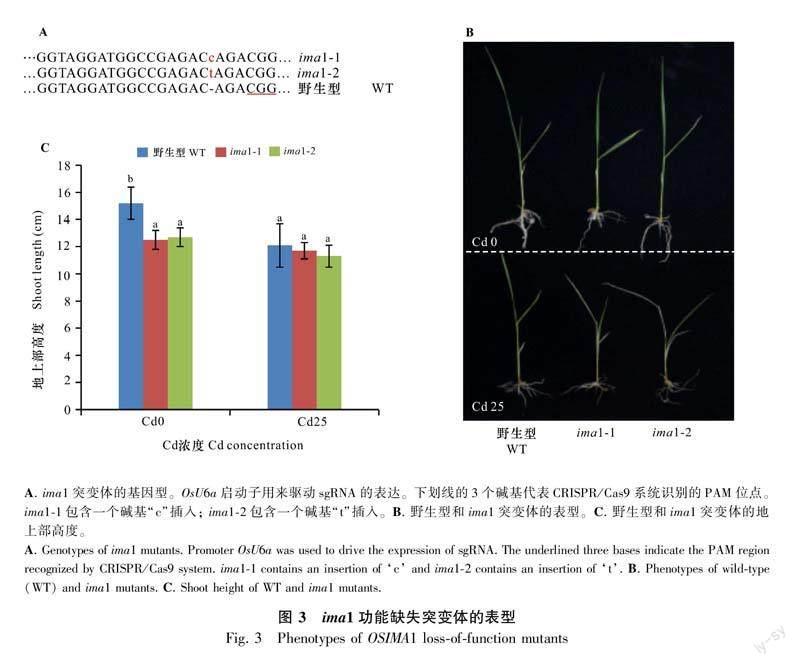
为了进一步研究OsIMA1在适应Cd胁迫方面的作用,本研究利用CRISPR-Cas9基因编辑技术对OsIMA1基因进行了编辑,并获得了两个ima1突变体。本研究在OsIMA1基因的编码区设计了一个特异性的靶点,用OsUba启动子驱动整合了该靶点的sgRNA,并构建了相应的表达载体。通过转化野生型水稻愈伤组织,获得了转基因植株。通过PCR鉴定及后代分离纯化,得到2个纯合的突变体株系ima1-1和ima1-2。2个株系均因基因编码区插入一个碱基导致移码突变(图3:A)。此外,本研究对野生型和2个功能缺失突变体的植株进行了Cd处理试验。在正常水培液生长条件下,2个突变体株系与野生型植株无明显差别(图3:B,C)。在Cd处理条件下,突变体植株表现出幼叶缺绿甚至萎蔫等症状(图3:B)。综上表明,OsIMA1功能缺失增加了植物对Cd的敏感性。
2.4 OsIMA1负调控Cd从根向叶的转运
本研究结果表明OsIMA1过表达植株比野生型更耐Cd胁迫,而ima1突变体则相反。为了进一步分析OsIMA1正调控Cd耐受的分子机制,本研究测定了野生型、OsIMA1过表达株系和ima1突变体的根和叶片中的Cd和Fe含量。先将在正常培养液中生长7 d的幼苗分别转移到不含Cd或含有25 μmol·L-1 CdCl2的培养液中培养7 d,然后分别取根和叶用来检测Fe和Cd的含量。与野生型相比,OsIMA1过表达植株的根和地上部都积累了更多的Fe,而ima1突变体则积累了较少的Fe;OsIMA1过表达植物在根中积累了较多的Cd,而ima1突变体在地上部积累了较多的Cd(图4)。综上表明,OsIMA1负调控Cd从根向地上部的运输。
3 讨论与结论
Cd对动植物的生长发育有害,近年来水稻受到Cd污染的事件频频发生。因此,减少植物的Cd吸收或提高植物对Cd胁迫的适应性,有利于保障粮食安全。Fe是植物生长发育所必需的营养元素,而缺Fe是导致人类贫血症的一个主要原因。水稻作为主粮作物,是人类Fe元素的一个主要来源。因此,培育富Fe稻米有益于改善人类健康。由于Fe和Cd之间的拮抗作用,额外施加Fe可以减少植物对Cd的吸收(Wu et al., 2012; Sebastian & Prasad, 2016)。因此,利用Fe和Cd的拮抗机制可以提高植物对Cd胁迫的适应性。水稻中存在两个OsIMA基因:OsIMA1和OsIMA2,它们的过表达能激活缺Fe诱导基因的表达,从而增加Fe的累积(Kobayashi et al., 2021)。本研究发现,Cd胁迫时OsIMA1和OsIMA2的表达显著上调。Cd处理试验发现OsIMA1的功能缺失突变体提高了水稻对Cd的敏感性,主要表现在植株叶片白化萎蔫。OsIMA1过表达一方面增加了Fe的积累,另一方面却抑制了Cd从根向地上部的转运,这可能是OsIMA1过表达植株对Cd耐受性提高的原因。因此,增加OsIMA1的表达水平是缓解植物Cd毒害的一个有效策略。
Meng等(2022)研究表明,IMA過表达可以提高拟南芥对Cd胁迫的适应能力。本研究发现OsIMA1过表达提高了水稻对Cd胁迫的耐受性。水稻根摄入的Fe和Cd经地上部转运至稻米中。OsIMA1过表达既促进了Fe在根部的累积,又促进了Cd在根部的积累。但是,OsIMA1过表达促进了Fe在地上部积累的同时,也抑制了Cd从根向地上部的转运。这表明OsIMA1可以通过特异性地抑制Cd从根向地上部的转运,提高植物对镉胁迫的适应性。Fe和Cd同属于二价离子,在体内可能被类似的转运蛋白识别。OsIMA1过表达植株根部累积的Fe可能竞争抑制了Cd与这些转运蛋白的结合。OsIMA1这种限制植物从根向地上部转运Cd的能力可能有助于减少稻米中的Cd累积。未来可以测试OsIMA过表达是否可减少稻米中的Cd含量。无论在拟南芥还是在水稻中,IMA均具有增加Fe吸收、增强Cd耐受的生物学功能,揭示了IMA在不同物种中的功能保守。IMA的高度保守性可能有助于打破基因功能物种特异性的限制(Grillet et al., 2018)。因此,未来研究可以测试IMA是否可以提高其他作物的耐Cd能力。
IMA小肽碳末端的17个氨基酸就足以发挥其分子功能(Grillet et al., 2018; Li et al., 2021)。目前,人工合成小分子多肽的技术已经非常流行。因此,未来有望将IMA开发为一种商用多肽,通过施肥的方式提高植物对Cd耐胁迫的适应性。综上所述,IMA可作为减轻植物Cd毒害的一个潜在可利用的小分子。进一步研究IMA调控Cd胁迫的机制,将为基因工程技术改造植物或培育耐Cd胁迫的农作物提供理论依据。
参考文献:
BAO T, SUN LN, SUN TH, 2010. Evaluation of iron on cadmium uptake by tomato, morel and leaf red beet in hydroponic culture [J]. J Plant Nutr, 33(5): 713-723.
CLEMENS S, 2006. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants [J]. Biochimie, 88(11): 1707-1719.
COHEN CK, FOX TC, GARVIN DF, et al., 1998. The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants [J]. Plant Physiol, 116(3): 1063-1072.
CZYZEWICZ N, YUE K, BEECKMAN T,et al., 2013. Message in a bottle: small signalling peptide outputs during growth and development [J]. J Exp Bot, 64(17): 5281-5296.
DAS P, SAMANTARAY S, ROUT GR, 1997. Studies on cadmium toxicity in plants: a review [J]. Environ Poll, 98(1): 29-36.
EIDE D, BRODERIUS M, FETT J, et al., 1996. A novel iron-regulated metal transporter from plants identified by functional expression in yeast [J]. Proc Natl Acad Sci USA, 93(11): 5624-5628.
FAN SK, FANG XZ, GUAN MY,et al., 2014. Exogenous abscisic acid application decreases cadmium accumulation in Arabidopsis plants, which is associated with the inhibition of IRT1-mediated cadmium uptake [J]. Front Plant Sci, 5(1): 721.
GRILLET L, LAN P, LI WF, et al., 2018. IRON MAN is a ubiquitous family of peptides that control iron transport in plants [J]. Nat Plants, 4(11): 953-963.
GUAN MY, ZHU YX, LIU XX, et al., 2019. Induction of S-nitrosoglutathione reductase reduces root cadmium uptake by inhibiting iron-regulated transporter 1 [J]. Plant Soil, 438(1): 251-262.
HE XL, FAN SK, ZHU J, et al., 2017. Iron supply prevents Cd uptake in Arabidopsis by inhibiting IRT1 expression and favoring competition between Fe and Cd uptake [J]. Plant Soil, 416(1): 453-462.
HINDT MN, AKMAKJIAN GZ, PIVARSKI KL,et al., 2017. BRUTUS and its paralogs, BTS LIKE1 and BTS LIKE2, encode important negative regulators of the iron deficiency response in Arabidopsis thaliana [J]. Metallomics, 9(7): 876-890.
HUANG G, DING C, LI Y, et al., 2020. Selenium enhances iron plaque formation by elevating the radial oxygen loss of roots to reduce cadmium accumulation in rice (Oryza sativa L.) [J]. J Hazard Mat, 398(1): 122860.
KIRKHAM MB, 2006. Cadmium in plants on polluted soils: effects of soil factors, hyperaccumulation, and amendments [J]. Geoderma, 137(1): 19-32.
KOBAYASHI T, NAGANO AJ, NISHIZAWA NK, 2021. Iron deficiency-inducible peptide-coding genes OsIMA1 and OsIMA2 positively regulate a major pathway of iron uptake and translocation in rice [J].J Exp Bot, 72(6): 2196-2211.
KOBAYASHI T, NAGASAKA S, SENOURA T,et al., 2013. Iron-binding haemerythrin RING ubiquitin ligases regulate plant iron responses and accumulation [J]. Nat Commun, 4(1): 2792.
KOBAYASHI T, NAGANO AJ, NISHUIZAWA NK. 2021. Iron deficieney-inducible peptide codin genes OsIMA1 and OsIMA2 positiveiy regulate a major pathway of iron uptake and translocation in rice [J]. J Exper Bot, 72(6): 2196-2211.
KORSHUNOVA YO, EIDE D, CLARK WG, et al., 1999. The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range [J]. Plant Mol Biol, 40(1): 37-44.
LEE S, AN G, 2009. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice [J]. Plant Cell Environ, 32(4): 408-416.
LI J, LIU JC, YAN CL, et al., 2019. The alleviation effect of iron on cadmium phytotoxicity in mangrove A. marina. Alleviation effect of iron on cadmium phytotoxicity in mangrove Avicennia marina (Forsk.) Vierh [J]. Chemosphere, 226(1): 413-420.
LI XL, ZHANG HM, AI Q, et al., 2016. Two bHLH transcription factors, bHLH34 and bHLH104, regulate iron homeostasis in Arabidopsis thaliana [J]. Plant Physiol, 170(4): 2478-2493.
LI Y, LU CK, LI CY, et al., 2021. IRON MAN interacts with BRUTUS to maintain iron homeostasis in Arabidopsis [J]. Proc Natl Acad Sci USA, 118(39): e2109063118.
LIANG G, ZHANG HM, LOU DJ, et al., 2016. Selection of highly efficient sgRNAs for CRISPR/Cas9-based plant genome editing [J]. Sci Rep, 6(1): 21451.
MAO QQ, GUAN MY, LU KX, et al., 2014. Inhibition of nitrate transporter 1.1-controlled nitrate uptake reduces cadmium uptake in Arabidopsis [J]. Plant Physiol, 166(2): 934-U730.
MATSUBAYASHI Y, 2014. Posttranslationally modified small-peptide signals in plants [J]. Ann Rev Plant Biol, 65(1): 385-413.
MENG X, LI W, SHEN R, et al., 2022. Ectopic expression of IMA small peptide genes confers tolerance to cadmium stress in Arabidopsis through activating the iron deficiency response [J]. J Hazard Mat, 422(1): 126913.
NIGHTINGALE ER, 1959. Phenomenological theory of ion solvation-effective radii of hydrated ions [J]. J Physiol Chem, 63(9): 1381-1387.
SCHUTZENDUBEL A, POLLE A, 2002. Plant responses to abiotic stresses: heavy metal induced oxidative stress and protection by mycorrhization [J]. J Exp Bot, 53(372): 1351-1365.
SEBASTIAN A, PRASAD MN, 2016. Iron plaque decreases cadmium accumulation in Oryza sativa L. and serves as a source of iron [J]. Plant Biol, 18(6): 1008-1015.
SHAO GS, CHEN MX, WANG WX, et al., 2007. Iron nutrition affects cadmium accumulation and toxicity in rice plants [J]. Plant Growth Regul, 53(1): 33-42.
TAKAHASHI F, HANADA K, KONDO T, et al., 2019. Hormone-like peptides and small coding genes in plant stress signaling and development [J].Curr Opin Plant Biol, 51(1): 88-95.
TAKAHASHI R, ISHIMARU Y, SENOURA T, et al., 2011. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice [J]. J Exp Bot, 62(14): 4843-4850.
TAVORMINA P, DE CONINCK B, NIKONOROVA N, et al., 2015. The plant peptidome: an expanding repertoire of structural features and biological functions [J]. Plant Cell, 27(8): 2095-2118.
VERT G, GROTZ N, DEDALDECHAMP F, et al., 2002. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth [J]. Plant Cell, 14(6): 1223-1233.
WEI BG, YANG LS, 2010. A review of heavy metal contaminations in urban soils, urban road dusts and agricultural soils from China [J]. Microchem J, 94(2): 99-107.
WU HL, CHEN CL, DU J, et al., 2012. Co-overexpression FIT with AtbHLH38 or AtbHLH39 in Arabidopsis-enhanced cadmium tolerance via increased cadmium sequestration in roots and improved iron homeostasis of shoots [J]. Plant Physiol, 158(2): 790-800.
XU SS, LIN SZ, LAI ZX, 2015. Cadmium impairs iron homeostasis in Arabidopsis thaliana by increasing the polysaccharide contents and the iron-binding capacity of root cell walls [J]. Plant Soil, 392(1): 71-85.
YAO XN, CAI YR, YU DQ, et al., 2018. bHLH104 confers tolerance to cadmium stress in Arabidopsis thaliana [J]. J Integr Plant Biol, 60(8): 691-702.
YOSHIHARA T, HODOSHIMA H, MIYANO Y, et al., 2006. Cadmium inducible Fe deficiency responses observed from macro and molecular views in tobacco plants [J]. Plant Cell Rep, 25(4): 365-373.
YUAN YX, WU HL, WANG N, et al., 2008. FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis [J]. Cell Res, 18(3): 385-397.
ZHANG J, LIU B, LI MS, et al., 2015. The bHLH transcription factor bHLH104 interacts with IAA-LEUCINE RESISTANT3 and modulates iron homeostasis in Arabidopsis [J]. Plant Cell, 27(3): 787-805.
ZHU YX, DU WX, FANG XZ, et al., 2020. Knockdown of BTS may provide a new strategy to improve cadmium-phytoremediation efficiency by improving iron status in plants [J]. J Hazard Mat, 384(1): 121473.
(責任编辑 李 莉 王登惠)
收稿日期: 2022-05-28
基金项目: 云南省科技厅科技计划项目(202003AD150007)。
第一作者: 彭凤(1996-),硕士研究生,主要从事植物铁营养代谢研究,(E-mail)1272277231@qq.com。
*通信作者: 梁岗,博士,研究员,主要从事植物矿质营养研究,(E-mail)lianggang@xtbg.ac.cn。
