4种润楠属植物的抗寒性与观赏特性评价
2023-07-11龙凤英刘春玲陈晓娟费永俊
龙凤英 刘春玲 陈晓娟 费永俊


摘要:为研究润楠属植物的抗寒性,检测了4种润楠属植物在自然低温胁迫下的生理指标:叶绿素、相对电导率(REC)、丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、游离脯氨酸(Pro)、可溶性蛋白(SP),分析其抗寒性。采用隶属函数法综合评价4种润楠属植物的抗寒性,结果为:宜昌润楠>刨花润楠>红梗润楠>小果润楠。采用层次分析法(AHP)对4种润楠属植物的观赏性进行综合评价,结果为:宜昌润楠>刨花润楠>小果润楠>红梗润楠。
关键词:润楠属;低温胁迫;隶属函数法;观赏性评价;层次分析法
中图分类号:S792.24文献标识码:A文章编号:1004-3020(2023)03-0027-11
Evaluation of Ecological Adaptability and Ornamental Characteristics of Four Species of Machilus
Long Fengying Liu Chunling Chen Xiaojuan Fei Yongjun
(1.College of Horticulture and Landscape Architecture,Yangtze UniversityJingzhou434000;2.Shuangzhu National Forest Farm of Zhuxi County in Hubei ProvinceShiyan442300)
Abstract:In order to study the cold resistance of Machilus plants,the physiological indexes of four species of Machilus under natural low temperature stress were detected: chlorophyll,relative conductivity (REC),malondialdehyde (MDA),superoxide dismutase (SOD),catalase (CAT),peroxidase (POD),free proline (Pro),and soluble protein (SP).The subordination function method was used to comprehensively evaluate the cold resistance of the four species of Machilus.The results were as follows: Machilus ichangnsis>M.pauhoi>M.rufipes>M.macrocarpa.The results of the comprehensive evaluation of the ornamental properties of four species of Machilus by using the analytic hierarchy process (AHP) were as follows:M.ichangnsis>M.pauhoi>M.microcarpa>M.rufipes
Key words:Machilus;low temperature stress;membership function method;ornamental evaluation;analytic hierarchy process
樟科Lauraceae潤楠属Machilus的植物为常绿乔木或灌木,中国约68种3变种,本属有许多优良的观赏树种[1]。宜昌润楠M.ichangnsis、小果润楠M.microcarpa、红梗润楠M.rufipes、刨花润楠M.pauhoi皆为常绿乔木,且新叶会呈现不同的颜色,具有较高的观赏价值。
润楠属植物主要分布在中国南方,可通过对其抗寒性的研究,根据其低温环境的生理表现,探究出这4种润楠属植物是否适合向北方引种。本研究选取的4种植物均属于春色叶树种,且树形优美,部分树种有特殊的香味,有较高的观赏价值,但在园林运用中较为少见。采用层次分析法(AHP)对4种润楠属植物进行观赏性评价,筛选出适应性强、观赏性高的植物,以期为润楠属植物资源开发和园林植物造景提供更多选择。
目前,润楠属植物在资源现状与保育[2]、群落调查[3]、栽培繁育[4]、抗逆性研究[5]、化学成分分析[6]等方面有丰富的研究,但将生态适应性和观赏共同研究的较少。
抗寒性方面,贺心茹等[7]研究了CaCl2溶液对黔桂润楠M.chienkweiensis幼苗抗寒性的影响,结果表明喷施浓度为15 mmol·L-1的CaCl2溶液可以明显提高黔桂润楠的抗寒性。袁婵璐[5]研究了CaCl2对三种润楠属幼苗抗寒性的影响,结果表明喷施CaCl2能提高植物的抗寒能力。何舒怀等[8]研究了外源水杨酸对广东润楠M.kwangtungensis抗寒性的影响,结果表明喷施10 mmol·L-1 SA溶液对增强广东润楠幼苗抗寒性有明显的促进作用。黎明等[9]研究了不同低温处理对宜昌润楠生理指标和叶片超微结构的影响,结果表明宜昌润楠能抵御一定的低温环境,但在-5 ℃时会出现明显的冻害症状。郭文妹[10]研究了2种润楠属植物的抗逆性,结果表明梨润楠M.pomifera与刻节润楠M.cicatricosa都具有较好的耐寒能力。
观赏性方面,吴林源等[11]利用AHP法,从花、叶、果及其他方面等10个指标对11种华南地区常见的润楠属植物进行观赏性综合评价,最终刨花润楠综合得分最高。张冬生等[12]对野生刨花润楠的9个观赏性指标进行评分并在层次分析法基础上运用熵技术修正各性状的权重,结果显示叶片形状是刨花润楠观赏价值评价的主要影响因素,其中叶大小、叶形和叶色的权重排在前三。胡南辉等[13]用Delphi法确定刨花润楠9项观赏指标的权重,再采用非整秩次RSR法对25株野生刨花润楠的观赏性进行综合评价,结果表明非整秩次RSR法可推广应用于植物观赏性综合评价中。张冬生等[14]利用ColorImpact软件对野生刨花润楠春梢图片色彩进行RGB数值提取与分析,结果表明刨花润楠春梢颜色丰富,值得作为园林景观树种推广种植。
本次试验在前人的研究基础上,对4种润楠属植物的抗寒性和观赏性进行分析与评价。
1材料和方法
1.1试验材料
试验材料为湖北省荆州市华中农高区太湖长江大学农业科技产业园内7 a生,长势相近的宜昌润楠、小果润楠、红梗润楠、刨花润楠的当年生侧枝的成熟叶片。
1.2试验地概况
试验地位于湖北省荆州市(111°15′~114°05′E,29°26′~31°37′N)。荆州位于湖北省南部,为冲积平原,属于亚热带季风气候,光照和热量充足,雨水充沛,有良好的气候资源供农作物和植物生长。
1.3试验方法
分别在2021年10月和2021年12月31日采集宜昌润楠、小果润楠、红梗润楠、刨花润楠的当年生侧枝的成熟叶片,置于-40 ℃冰箱保存,后续测定生理指标。采集的叶片未用液氮速冻,装在塑料自封袋内进行保湿,3次重复。12月25日~12月31日最低温分别为-1 ℃、-1 ℃、0 ℃、1 ℃、0 ℃、0 ℃和2 ℃。
1.4测定方法
叶绿素总含量(Chl)的测定[15]:采用分光光度法测定,取剪碎混匀的叶片鲜样0.2 g置于25 mL试管中,加10 mL 80%乙醇于暗处室温浸提24 h,至叶片褪色。用80%乙醇定容后,取提取液在波长663 nm和645 nm下测定吸光度,以80%乙醇为空白对照。
叶绿素总含量(mg·g-1Fw)=(20.21A645+8.023A663)×提取液总量(mL)÷样品鲜重(g)×1 000。
相对电导率(REC)的测定[15]:采用蔡庆生的方法,使用电导仪测定。
相对电导率(%)=(E1-E0)/(E2-E0)×100%,式中,E0为蒸馏水的值,E1为叶片煮沸前的值,E2为叶片煮沸后的值。
丙二醛(MDA)含量的测定[15]:采用硫代巴比妥酸法色测定,取1 g鲜叶加石英砂和2 mL 10%三氯乙酸研磨成匀浆,再加8 mL 10%三氯乙酸研磨成匀浆,4 000 r·min-1离心10 min后,取上清液。取2 mL上清液和2 mL 0.6%硫代巴比妥酸溶液加入试管,摇匀后沸水浴中反应15 min,迅速冷却后离心,再取上清液分别在波长532 nm、600 nm和450 nm下测定吸光度。
丙二醛含量(μmol·g-1)=[6.45(A532-A600)-0.56A450](μmol·mL-1)×提取液总量(mL)÷样品鲜重(g)。
抗氧化酶活性测定:超氧化物歧化酶(SOD)采用氮蓝四唑(NBT)光还原法测定[15];过氧化氢酶(CAT)采用紫外吸收法测定[15];过氧化物酶(POD)活性采用愈创木酚氧化比色法测定[15]。
有机溶质测定:游离脯氨酸含量(Pro)采用茚三酮显色法测定[15];可溶性蛋白含量(SP)采用考马斯亮藍染色法测定[15]。
1.5数据处理
1.5.1显著性分析
对测得的数据使用Excel 2018进行统计分析,使用SPSS 25.0进行单因素方差分析,比较不同物种在胁迫前的差异,显著性水平设定为P=0.05,多重比较选择Duncans方法;使用配对T检验法比较同一物种在低温胁迫前和低温胁迫后的差异。
1.5.2隶属函数分析
采用隶属函数法综合评价润楠属植物的抗寒性,选取Chl、REC、MDA、SOD、CAT、POD、Pro、SP等指标计算分析。
隶属函数:U(Xijk)=(Xijk-Xmin)/(Xmax-Xmin);
反隶属函数:U(Xijk)=1-(Xijk-Xmin)/(Xmax-Xmin)。
式中,U(Xijk)为第i个品系第j个取样阶段第k项指标的隶属度,且U(Xijk)∈〔0,1〕;Xijk表示第i个品系第j个取样阶段第k个指标测定值;Xmax、Xmin为所有参试品系中第k项指标的最大值和最小值[16]。
1.5.3层次分析
采用SPSSPRO进行层次分析(AHP)。先构建出影响楠木观赏性的评价指标体系:目标层A(4种润楠属植物观赏性评价体系)、约束层B(影响评价的主要因子:花果、叶、树形、其他)、标准层C(具体的评价标准:花量、果实大小、果柄颜色、新叶色、新梢/叶柄颜色、绿期、树形、冠幅、株高、滞尘能力、繁殖难度、耐寒性)、最底层D(4种待评价的润楠属植物)。
由于基地的树苗处于幼年期,所以花量、果实大小、果柄颜色、株高、冠幅的数值参照中国植物志中4种润楠属植物的描述和图片进行评分;新叶色、新梢/叶柄颜色、绿期的数值参考2021年4月29日拍摄的图片(图1)进行评分;滞尘能力[17]、繁殖难度[11]、耐寒性(以本研究为依据)等指标参考研究文献进行评分。再参照1~9比例标度法确定出各评价标准的权重,然后参考相关观赏性评价文献资料[11-13]制定具体的评价指标以及评分标准,最后对4种润楠属植物进行评分,使用加权后的得分进行观赏性排序。
2结果与分析
2.1低温胁迫的影响
2.1.1低温胁迫对植物叶绿素含量的影响
如表1所示,在常温时,刨花润楠的叶绿素含量最高,其次是小果润楠、红梗润楠、宜昌润楠;在低温胁迫后,小果润楠叶绿素含量最高,其次是红梗润楠、刨花润楠、宜昌润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠叶绿素含量的变化率=(低温胁迫后观测值/低温胁迫前观测值-1)×100%,分别为-58.28%、55.42%、14.08%、-53.52%。小果润楠经过低温胁迫后,叶绿素含量上升的最多,可见低温胁迫促使小果润楠产生更多的叶绿素以维持光合作用。宜昌润楠在受到低温胁迫后叶绿素含量的变化率最大,可见其在光合作用方面受低温胁迫的影响较大(表1)。
2.1.2低温胁迫对植物相对电导率(REC)的影响
如表2所示,低温胁迫前,刨花润楠的REC最大,其次是小果润楠、红梗润楠、宜昌润楠;低温胁迫后,小果润楠的REC最大,其次是红梗润楠、刨花润楠、宜昌润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠REC的变化率分别为-22.45%、26.62%、-8.90%、-28.42%。可見,仅小果润楠的REC上升了,其他三种润楠REC下降的原因可能是这三种润楠的抗寒性较强,在气温回升时细胞的质膜透性开始恢复正常,使渗透速度下降[18]。
2.1.3低温胁迫对植物丙二醛(MDA)的影响
如表3所示,低温胁迫前,红梗润楠的MDA含量最高,其次是小果润楠、宜昌润楠、刨花润楠;低温胁迫后,宜昌润楠MDA的含量最高,其次是刨花润楠、小果润楠、红梗润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠MDA的变化率分别为215.79%、29.50%、-45.70%、305.60%。可见,刨花润楠和宜昌润楠MDA含量上升幅度较大,表明低温对两者的伤害较大;红梗润楠MDA含量下降可能是物种差异的原因[19]。
2.1.4低温胁迫对植物超氧化物歧化酶(SOD)的影响
如表4所示,低温胁迫前,刨花润楠的SOD活性是最高的,其次是宜昌润楠、红梗润楠、小果润楠;低温胁迫后,刨花润楠的SOD活性是最高的,其次是红梗润楠、宜昌润楠、小果润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠SOD活性的变化率分别为-19.29%、-58.61%、9.26%、5.34%。小果润楠SOD活性的变化率最大,说明小果润楠SOD活性受低温胁迫的影响最大,SOD活性下降明显表明其叶片已无法抵御低温伤害;低温胁迫激发了刨花润楠和红梗润楠的保护性应激反应,使SOD酶活性有所上升。
2.1.5低温胁迫对植物过氧化氢酶(CAT)的影响
如表5所示,低温胁迫前,CAT的活性最高的是红梗润楠,其次是小果润楠、宜昌润楠、刨花润楠;低温胁迫后,4种润楠属植物CAT的活性整体呈下降趋势,CAT活性最高的是宜昌润楠,其次是红梗润楠、刨花润楠、小果润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠CAT活性的变化率分别为-11.22%、-58.35%、-41.94%、-9.64%。可见,低温胁迫对小果润楠CAT活性的影响最大,其CAT酶活性下降显著。
2.1.6低温胁迫对植物过氧化物酶(POD)的影响
如表6所示,低温胁迫前,POD活性最高的是刨花润楠,其次是宜昌润楠、小果润楠、红梗润楠;低温胁迫后,POD活性最高的也是刨花润楠,其次是宜昌润楠、红梗润楠、小果润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润POD活性的变化率分别为136.10%、-2.75%、25.05%、9.77%,只有小果润楠的POD活性降低。低温胁迫前后,刨花润楠的POD活性始终是显著高于其他三种植物,说明低温胁迫对于刨花润楠的POD影响不大;宜昌润楠POD酶活性显著升高,表明受到低温刺激宜昌润楠能够快速提高POD酶活性来保护细胞。
2.1.7低温胁迫对植物游离脯氨酸(Pro)的影响
如表7所示,低温胁迫前,刨花润楠的Pro含量最高,其次是小果润楠、宜昌润楠、红梗润楠;低温胁迫后,刨花润楠的Pro含量最高,其次是宜昌润楠、红梗润楠、小果润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠其Pro活性的变化率分别为32.47%、-10.89%、43.43%、11.71%。低温胁迫后,红梗润楠、宜昌润楠和刨花润楠的Pro含量有不同程度的升高,仅小果润楠的Pro含量下降了,可见低温胁迫对小果润楠的伤害更加严重。
2.1.8低温胁迫对植物可溶性蛋白(SP)的影响
如表8所示,低温胁迫前,SP含量最高的植物是宜昌润楠,其次是红梗润楠、刨花润楠、小果润楠;低温胁迫后,SP含量最高的是宜昌润楠,其次是刨花润楠、小果润楠、红梗润楠。低温胁迫后,宜昌润楠、小果润楠、红梗润楠和刨花润楠的SP含量的变化率分别为19.49%、19.24%、-33.46%、21.03%。可见,仅红梗润楠的SP含量降低,说明红梗润楠受低温胁迫的伤害较大。
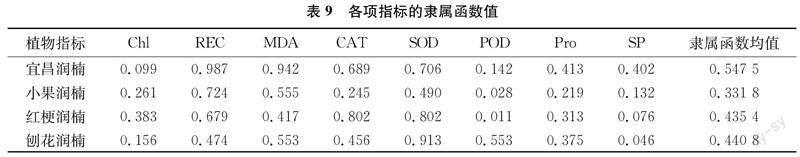
2.1.9隶属函数法综合评价抗寒性
Chl、SOD、POD、Pro和SP在逆境下的含量(或活性)变化与植物自身的抗寒性呈现正相关,用隶属函数计算;与植物抗寒性呈负相关的MDA和REC用反隶属函数计算。用隶属函数均值对4种润楠属植物进行排序,可知抗寒性的强度依次是:宜昌润楠>刨花润楠>红梗润楠>小果润楠(表9)。
2.2观赏性评价
2.2.1构建AHP评体系
根据4种润楠属植物现场情况以及文献资料[10-12]的描述,筛选出观赏特征的优良指标,采用理论分析法和专家咨询法制定评价指标体系(表10)。
2.2.2指标权重和评分标准的确定
参照1~9比例标度法,分别对评价体系中的约束层、标准层各因素的重要性进行两两比较,并借助SPSS PRO构建判断矩阵,进行一致性检验,计算出各个指标的权重。由表11可知,对目标层的权重排序第一的是新叶色(30.680%),其次为树形(15.776%)、花量(12.121%),说明新叶色、树形和花量是影响这4种润楠属植物观赏性的重要因素。
表12中数据来源于有关楠木观赏性评价的文献[11-13]、中国植物志和楠木有关滞尘性[17]、耐寒性[5,9]以及繁殖相关的文献。
2.2.3观赏性评价
通过阅览中国植物志图库、中国自然标本馆等网站信息和现场观察(2021年4月29日拍摄的照片),对4种润楠属植物的12个指标进行评分,再通过加权求出最后的得分,最后根据得分进行排序。4种润楠属植物的综合评分相差不大,都具有一定的观赏价值,观赏性排序为:宜昌润楠>刨花润楠>小果润楠>红梗润楠。可见,宜昌润楠在这4种润楠属植物中的观赏性与抗寒性均最高,最适宜运用于园林观赏中(表13)。
3结论与讨论
3.1抗寒性评价
Chl的含量在冬季会减少,因为低温胁迫会导致叶绿素合成受阻。宜昌润楠和刨花润楠经过低温胁迫后,Chl的含量减少了,但小果润楠和红梗润楠的Chl含量升高了。与本研究不同的是,在袁婵璐[5]的研究中低温胁迫后宜昌润楠的Chl含量升高了,可能是袁婵璐的低温处理的时间较短,因而出现不同的结果;小果润楠和红梗润楠的Chl含量的变化与袁婵璐[5]研究中的粉叶润楠表现一致。小果润楠和红梗润楠的Chl含量升高也可能是在气温回升时Chl的含量在逐步提高[17]。
植物受到低温伤害时,细胞质膜透性会有不同程度增大,电解质会不同程度外渗,从而导致相对电导率提高[20],可用相对电导率反映植物的细胞膜伤害程度,进而判断其抗寒性。此次研究中宜昌润楠的相对电导率变化最小,说明细胞膜未受到严重损伤,可见宜昌润楠表现出良好的抗寒性。
MDA是膜脂过氧化过程的最终分解产物,其含量可以反映膜系统受伤害的程度和植物的抗逆性[21]。此次研究中,受到低温胁迫后宜昌润楠、刨花润楠和小果润楠的MDA含量明显升高,大量积累的MDA可能导致细胞膜功能受阻。相反的是红梗润楠的MDA含量下降了,可能是不同物种在长期低温环境中的MDA含量变化规律不同[19]。与本研究表现相同的是,在高福元等[19]的研究中,美国红栌在持续低温初期(6~9 d)MDA含量下降,随着低温胁迫时间延长又迅速升高,可能是胁迫初期植物保护酶产生的防御反应,酶活力增强,清除活性氧和自由基的能力增强。王微[22]对紫花苜蓿几个抗寒生理指标的研究中出现MDA含量无变化规律的现象,指出抗寒反应的生理机制比较复杂,单从某一个生理指标来鉴定抗寒性比较困难。
SOD、CAT和POD是三种保护性酶[23]。本研究中,除CAT活性表现相同外,其他酶在低温下的表现不一样,可能是CAT对低温比较敏感。不同的物种,SOD和POD活性在低温下的表现不一样,可能是物种本身抗寒性不同,有不同的抵御逆境的对策。在黎明等[9]的研究中宜昌润楠SOD和CAT活性在低温胁迫下表现出先上升后下降的趋势,在5 ℃时二者均达到峰值,而后开始下降。与本研究表现一致,说明低温胁迫表现出对宜昌润楠SOD和CAT酶的伤害。在郑家基等[24]的研究中,香蕉的POD活性随低温程度加强呈现下降趋势,与宜昌润楠的表现不同,与小果润楠POD活性相同。可见,不同物种的POD活性对低温的反应不同,这可能是植物适应逆境条件的一种积极的代谢适应对策[21]。郭祥泉[25]认为随着低温胁迫时温度的降低,SOD酶活性有一个温度拐点,在拐点前其活性急剧上升,在拐点后又下降,以此来定量研究不同树种及不同个体的低温临界值。可能红梗润楠和刨花润楠的SOD酶活性的温度拐点与宜昌润楠、小果润楠的不一致。
低温胁迫下植物通过积累可溶性糖、可溶性蛋白、脯氨酸等物质降低细胞渗透势,保护生物膜,维持细胞结构提高植物抗寒性[26]。在李高志[27]对三种楠属植物的抗寒性研究中,浙江楠在低温胁迫过程中脯氨酸含量呈现先升后降的趋势,本研究中只有小果润楠的Pro含量下降了,可能温度过低致使其Pro含量下降。低温胁迫下,只有红梗润楠的SP含量下降了,与黄枝润楠[5]在低温胁迫下的表现一致。何开跃等[28]认为:脯氨酸的积累可能是植物对低温的一种适应或者细胞受损的一种表现,与植物的耐寒能力无明确相关关系。
因为植物抗寒生理生化变化较为复杂,且不同指标存在交互作用,所以根据单一的指标判断植物的抗寒性具有局限性[29],只有对这些指标进行综合分析,才能准确地判断各物种的抗寒强弱。目前用于抗寒性综合评价分析的方法有聚类分析法、主成分分析法和隶属函数法等,其中隶属函数法应用较为广泛。本研究采用隶属函数法对4种润楠属植物的抗寒性进行综合评价,结果表明宜昌润楠的抗寒性最强,其次为刨花润楠、红梗润楠、小果润楠。徐文斌等[30]在2015~2016年冬春武汉植物园61种樟科植物冻害调查中表示宜昌润楠是属于抗冻的树种,其调查数据表明宜昌润楠在冻害中的表现好于刨花润楠,与本次抗寒性综合评价结果一致。
3.2观赏性评价
結合隶属函数法的结果来看,宜昌润楠的抗寒性是最强的,加上其在树形、树干、叶色和花等方面都有良好的景观效果,表明宜昌润楠适合广泛推广。在实际应用时,可采用孤植、列植或群植等配植方法应用于城市道路、主题公园、生活居住区以及园林绿化。刨花润楠的抗寒性较好,对土壤要求不严,树形美观,树干通直,树叶繁茂,春季时叶紫红,冬季可观黑果。刨花润楠也是非常重要的经济树种,是制造蜡烛、肥皂蚊香、祭香的主要原料[31],其枝叶和果还可提取芳香油。红梗润楠的抗寒性不如前面两种,其花序轴及花梗呈红紫色,花大,花期和果期都比较早,可作为园林中观果观叶树种;其根皮可入药,有舒筋活络、消肿止痛、解毒等效用。小果润楠的新叶鲜红,新梢粉红,树形秀丽,有较好的观赏效果。小果润楠在湖北省的生长情况良好,可以作为园林中良好的绿化树种,且小果润楠的种子繁殖容易,生长迅速,也是极具开发利用价值的重要用材树种。
参考文献
[1]中国科学院中国植物志编辑委员会.中国植物志(第三十一卷)[M].北京:科学出版社,1982:7-63.
[2]徐振东,杨曼,胡蝶,等.润楠属主要树种的资源状况、研究现状和开发利用前景[J].长江大学学报(自科版),2016,13(3):13-17.
[3]汪杨,王奥,朱司甲,等.神农架地区楠木种群结构及分布特征[J].福建林业科技,2017,44(4):1-5,27.
[4]张冬生,张锡洲,谢金兰,等.刨花润楠播种育苗技术探讨[J].园艺与种苗,2016(2):13-14,17.
[5]袁婵璐.CaCl2对三种润楠属树种幼苗抗寒性研究[D].荆州:长江大学,2017.
[6]里艳茹,黄胜男,寻园,等.粗壮润楠化学成分及其活性研究[J].中南药学,2022(10):2279-2285.
[7]贺心茹,陈建,胡蝶,等.CaCl2溶液处理对黔桂润楠幼苗抗寒性的影响[J].长江大学学报(自然版)理工卷,2019,16(2):65-68,77.
[8]何舒怀,马永征,费永俊.外源水杨酸对广东润楠抗寒性的影响[J].江苏农业科学,2018,46(8):155-158.
[9]黎明,杨芳绒,商卫卫.低温对宜昌润楠生理指标及叶片超微结构的影响[J].西北林学院学报,2010,25(4):19-22.
[10]郭文妹.两种润楠属植物幼苗的抗逆性研究[D].广州:华南农业大学,2017.
[11]吴林源,黄稚清,丁释丰,等.基于AHP法的润楠属树种观赏性评价[J].福建林业科技,2019,46(4):4.
[12]张冬生,谢金兰,叶雪兰,等.基于层次分析法与熵技术法的野生刨花润楠观赏性评价[J].广东林业科技,2016,32(1):23-30.
[13]胡南辉,邓春亮,苏振伟,等.基于非整秩次RSR法的刨花润楠观赏性评价[J].湖南理工学院学报:自然科学版,2018,31(3):6.
[14]张冬生,陈聪,叶雪兰,等.梅州地区野生刨花楠春梢色彩的调查与分析[J].广东园林,2016,38(137):77-79.
[15]张治安,陈展宇.植物生理学实验技术[M].长春: 吉林大学出版社,2008.
[16]徐超,苏敬,王雨亭,等.强冷空气过程中3种柑橘光合生理特性的变化及抗寒性综合评价[J].果树学报,2022(12):2309-2318.
[17]黄承石,潘珈瑶,王东,等.5种润楠属植物叶表皮结构和滞尘能力研究[J].河南科技大学学报:自然科学版,2019,40(2):7.
[18]商卫卫.宜昌润楠的耐低温生理研究[D].郑州:河南农业大学,2009.
[19]高福元,张吉立,刘振平,等.持续低温胁迫对园林树木电导率和丙二醛含量的影响[J].山东农业科学,2010(2):47-49,81.
[20]杨向娜,魏安智,杨途熙,等.仁用杏3个生理指标与抗寒性的关系研究[J].西北林学院学报,2006(3):30-33.
[21]张海燕,樊军锋,高建社,等.5个欧美杨无性系抗寒性测定与评价[J].西南林业大学学报 (自然科学),2020,40 (2):36-43.
[22]王微.低温胁迫下紫花苜蓿几个抗寒生理指标的变化规律[D].长春:东北师范大学,2009.
[23]李冰,张敬敬,高秀瑞,等.低温胁迫下不同基因型西瓜抗寒性综合评价[J].中国瓜菜,2019,32(4):16-19.
[24]郑家基,谢厚钗,肖燕惠,等.香蕉越冬期ATPase和保护酶的活性及膜脂过氧化[J].福建农业大学学报,1996(4):35-39.
[25]郭祥泉 .用膜透性、SOD 酶活性和膜脂肪酸成分探讨林木抗寒性[J].西南林业大学学报 ,2012,32(4):6-11.
[26]任军,黄志霖,曾立雄,等.低温胁迫下植物生理反应机理研究进展[J].世界林业研究,2013,26(6): 15-20.
[27]李高志.三种楠属植物的抗寒性研究[D].杭州:浙江农林大学,2016.
[28]何開跃,李晓储,黄利斌,等.福建柏抗寒生理指标变化研究(英文)[J].南京林业大学学报(自然科学版),2002(5):10-14.
[29]王红平,董铁,刘兴禄,等.5 个苹果砧木品种枝条的低温半致死温度及耐寒性评价 [J].果树学报,2020,37 (4):495-501.
[30]徐文斌,詹玉玲,吴风璨,等.2015~2016年冬春武汉植物园61种樟科植物冻害调查[J].湖北林业科技,2016,45(3):36-39,78.
[31]胡希华.刨花润楠的优良特性及育苗栽培技术[J].湖南林业科技,2006(1):65-66.
(编校:唐岚)
