大岗山植物群落物种多样性与群落结构研究
2023-07-11张雨桐刘华陈永富赵峰凌成星曾浩威马永康
张雨桐 刘华 陈永富 赵峰 凌 成星 曾浩威 马永康


关键词:物种多样性;群落结构;物种组成;大岗山
中图分类号:S718.54 文献标识码:A
随着时代的发展和人类活动的影响,生物多样性正以前所未有的速度丧失。2019 年,一个科学家小组得出结论,现在大约有100 万动植物物种面临着灭绝的威胁,甚至有一部分物种将在未来几十年内灭绝[1]。生物多样性为生态系统提供着保护,免其受到气候突变、病虫害及自然灾害等不利环境条件的影响[2]。此外,已有研究表明,生物多样性与生态系统的服务功能之间存在联系,生物多样性高的生态系统提供着更为丰富的服务功能[3-4]。为了采取有效的措施,减缓生物的灭绝速度,保护受到威胁的生物多样性,对生物多样性的研究刻不容缓。
物种多样性(species diversity)是指地球上动物、植物、微生物等生物类的丰富程度,其代表着物种演化的空间范围和对特定环境的生态适应性,是进化机制的最主要产物及生物有机体本身多样性的体现[5]。一个区域内不同物种的数量是生物多样的关键组成部分,珍稀濒危物种的生长或外来物种的引入都可能导致本土物种的生物多样性丧失[6-8]。物种多样性指数为量化多样性提供了依据[9],陈廷贵等[10]比较了15 种常用的多样性指数,结果显示,在综合反映多样性变化上香农-威纳( Shannon-Wiener) 多样性指数和辛普森(Simpson)多样性指数效果最好,这2 个指数也是研究者们最常用的多样性指数。皮诺(Pielou)均匀度指数以Shannon-Wiener 指数和Simpson 指数为基础,是常用的物种均匀度测定方法[11]。
群落结构在组成、功能和动态变化等方面表现出的差异性[12],不仅反映着群落对环境的适应性,也影响着生态系统的功能[13]。群落的组成结构可以从多方面反映区域植物多样性的特征,分析群落的组成及区系成分是认识群落特征、变化、发展趋势和生物多样性的基础[14]。在大部分南方森林生态系统中,局部小环境差异大,从而使得不同群落乔灌草多样性表现为草本层>灌木层>乔木层、灌木层>乔木层>草本层或乔木层最高等多种类型[15]。在不同森林生态系统中,乔灌草物种多样性通常呈现明显分异,三者为负相关关系[16]。由此可见,物种多样性是表示群落特征的重要指标,在反映植物群落的结构类型、演替阶段和稳定程度等方面具有重要意义[17-18]。
当前,我国林业生态效益的发展逐渐从获取直接的经济效益向改善生态环境质量获取间接效益转变[16]。随着全国生态建设的持续推进,“近自然森林”成为现代林业建设的重点。在次生林和人工林经营过程中,通过调整树种配置及比例、林分结构等,使其接近生态效益高的天然林分,以达到快速提升林分生态功能的效果,实现近自然化经营[16, 19]。研究特定气候条件下的天然次生林,对于区域森林生态系统管理和近自然经营具有指导性意义[15]。此外,次生林可以直接快速地观察到群落动态和受干扰的群落中森林结构的变化,为研究群落组成提供了基础。β 多样性中的物种相似性和物种转化率在衡量2 个或多个地点之间的群落组成变化上受到广泛应用,探究β 多样性可以为次生林演替过程提供指导意义[20]。
中国的亚热带地区与世界同纬度相比,是除沙漠以外最暖热的区域,雨量也远超同纬度地区,因此亚热带森林生态系统有着丰富的物种、较高的生产力以及强大的生态系统的生态功能。大岗山地处中亚热带地区,山区内的气候、土壤等环境差异形成了森林的空间异质性,异质性程度越高,则意味着更多的小生境和小气候条件,从而满足更多具有不同生态位的物种共存[21]。对森林进行科学合理地经营与保护,发挥其潜在自然物质的快速恢复能力、产生巨大的物质产品和生态文化服务,是我国森林经营迫切需要解决的问题[22]。
大岗山的植物群落种类丰富且保存完好,是较为理想的研究場所,近年来的研究偏向单一的植物群落或群落功能,如常绿阔叶林的物种组成[23]、杉木人工林的土壤肥力和碳储量等[24-25],对区域性的群落结构及物种多样性的研究较少。本研究以大岗山主要山区的植物群落为研究对象,通过调查分析区域内整体及各个样带之间的物种多样性和群落结构,描述群落的组成,探寻其分布及变化规律,为揭示此类森林的生物多样性潜力及维持机制等提供理论依据。
1 材料与方法
1.1 研究区概况
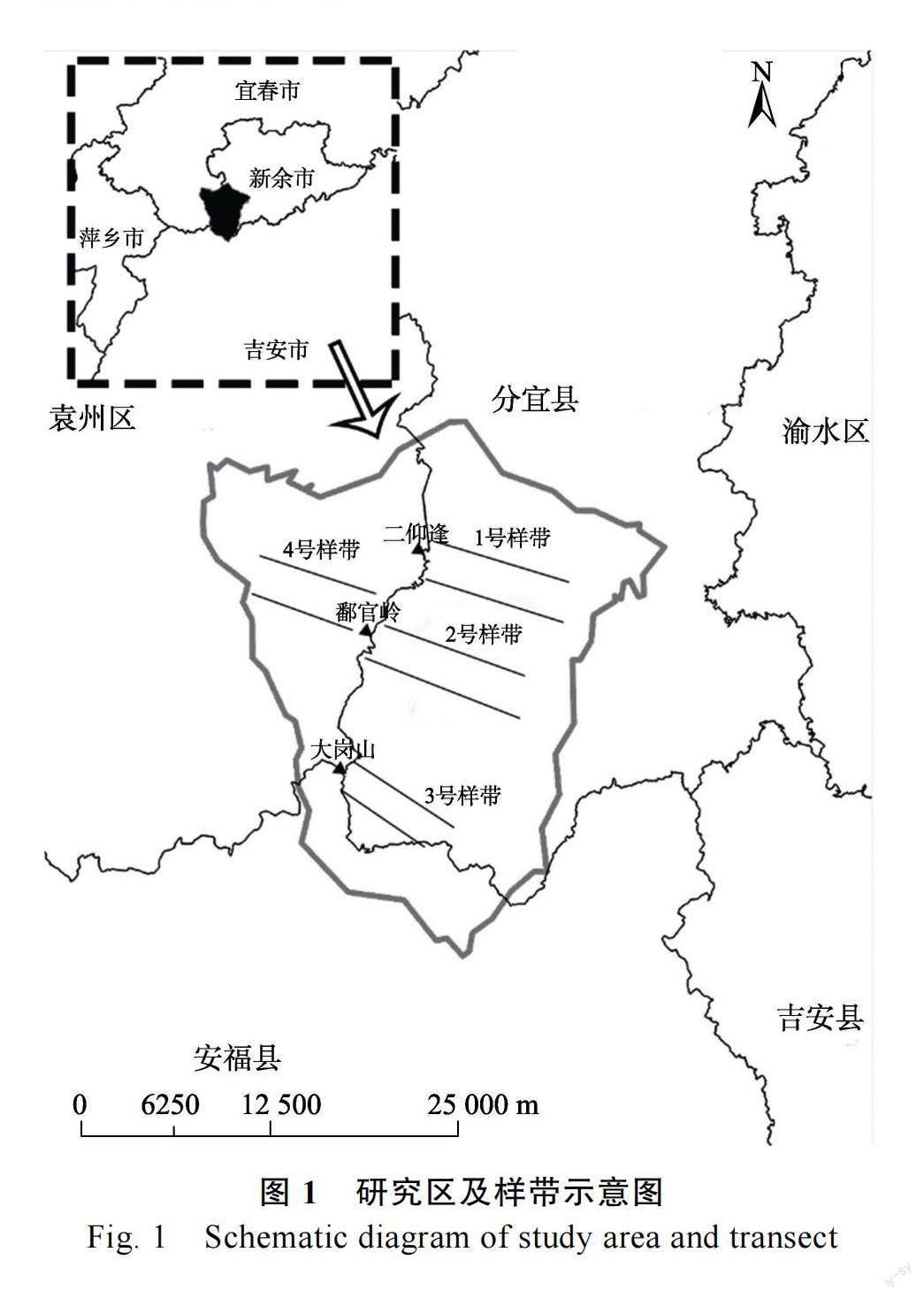
研究区主要位于江西大岗山自然保护区内(图1),研究区范围114°30′~114°37′E,27°34′~27°43′N,海拔116~1031 m,坡度<43°,土壤类型为红壤和黄壤,由低海拔至高海拔土壤类型及亚类的分布依次为红壤、黄红壤、黄壤和黄棕壤,其中红壤分布最广,分布海拔为110~470 m。研究区位于中亚热带,气候类型属于亚热带季风湿润气候,年均气温16 ℃,1 月平均最低温2.1 ℃,7 月平均最高温26.5 ℃,降水主要集中在4—8月,年均降水量1591 mm[26]。该区域雨量充沛,日照充足,气候温和,无霜期长,森林覆盖率高,植被类型主要由常绿阔叶林、杉木林、毛竹林等构成,保护区虽受人为干扰较小,但也并非完全未被破环的原始林,经过几十年的自然演替,顶级次生林、次生林林冠几乎完全封闭[27]。截至1999 年7 月大岗山亚热带林业试验中心的调查统计,该区现有植物1888 种,分属254 科847 属,其中木本植物780 种,隶属760 属,占中国被子植物种类的24%;裸子植物22 属,占中国裸子植物种类的65%;蕨类植物36 属,占中国蕨类种类的16%[23]。自然保护区内还包含国家级重点保护的珍稀动植物40 多种,如猴、獐、鹿、兔、石樟、银杏、水杉等[28]。
1.2 样地设计与调查方法
样地设计:2021 年7 月,以研究区具有代表性的3 个山峰为起点,根据植被覆盖类型沿海拔变化的情况向山峰南、北2 个方向设置样带,共计4 个样带(图1),在每条样带上,按照海拔梯度设置5 条等距离水平样线,在每个样线上设置3 个大样方,大样方垂直投影面积为20 m30 m,各样方间距在50 m 以上,共计60 个大样方。沿途增设2 个含有珍稀濒危树种的样方,实际调查62 个样方。其中1~15、61 和62 号样方位于1 号样带;16~29 号样方位于2 号样带;30~43、59、58 号位于3 号样带;44~57 号位于4 号样带。在每个大样方的4 个角及中央各设置1 个中样方(共5 个)进行灌木植物调查,每个5 m5 m;在每个矩形中样方的内角设置1 个小样方进行植物调查,每个1 m1 m。
样地调查:记录各个样地坐标、海拔、坡位、坡向等信息;对大样方内胸径大于等于5 cm 的乔木树种进行每木检尺,记录乔木层树种名称、胸径、树高等;中样方内调查并记录灌木及胸径小于5 cm 的乔木树种的名称、高度、基径、分布及株数等;小样方内调查并记录草本、苔藓等植物的名称、盖度、高度、分布及生长状况等。
1.3 植物组成成分
裸子植物依据郑万钧[29]分类系统,被子植物依据哈钦松[30]分类系统,种子植物地理分布以吴征镒[31]的中国种子植物属的分布区类型为依据,珍稀濒危植物参考《中国生物多样性红色名录》和江西珍稀植物代表物种[32]。
1.4 数据处理
根据野外调查数据,统计各样带以及区域乔木层、灌木层、草本层植物的科、属、种、个体多度、频度、平均胸径等内容,并计算指标指数,具体如下,所有数据利用Excel 软件进行整理计算和图表绘制。
1.4.1 物种重要值 重要值表示物种在群落中重要性的指标,其计算公式为:
式中,Pi为群落中第i 个物种的重要值,相对密度=某种植物的密度/全部植物的总密度;相对频度=该物种的频度/所有种的频度总和;相对显著度=样方中该种个体胸高断面积和/样方中全部个体胸高断面积总和。
1.4.2 物種多样性 采用α多样性指数法分别计算大岗山植物群落整体及各样带的乔木层、灌木层、草本层的物种丰富度(S)、Shannon-Weiner多样性指数(H)、Simpson 多样性指数(D)和Pielous 均匀度指数(J),计算公式如下:
式中,a 和b 分别为2 个基本观测单元内观测到的物种数量;c 为2 个基本观测单元内观测到的共有物种数量,g(H)和l(H)分别为沿生境变化(H梯度)增加和失去的物种数量。
2 结果与分析
2.1 群落种类、区系组成与分布
根据样地调查统计,合计调查17 979 个独立个体,其中乔木层共有4058 个,灌木层9596 个,草本层4325 个,植株密度4833 株/hm2,总计120科286 属486 种,其中蕨类植物39 种,隶属17科27 属,裸子植物隶属3 科5 属,总计7 种,被子植物共计100 科254 属440 种。前5 位的数量优势科中蔷薇科(Rosaceae)排名第1 位,共计31 种,隶属11 属;第2 位为樟科(Lauraceae)26 种,隶属7 属;第3 位为茜草科(Rubiaceae),13 属20 种,第4 位为壳斗科(Fagaceae),6 属18 种;豆科(Fabaceae)排名第5 位,共计17 种,隶属13 属。单种属190 个,占总数66.43%,研究区属种系数(植物属数与植物种数之比)为58.85%。属种系数略高,说明生境条件稳定,一致性高,也说明植物分化程度低,植物物种多样性以单种或寡种占优势。区域内包含国家重点保护及江西特有植物共7 种,分别为南方红豆杉、闽楠、胡桃、香樟、华木莲、伯乐树和篦子三尖杉,濒危等级及保护等级见表1。濒危种中,南方红豆杉频度株数最高,但伯乐树和江西特有种华木莲均只发现1 科,只在大岗山零星分布。
根据中国种子植物属的分布类型进行分布统计,在大岗山286 个属当中,种子植物有259 个属,其中仅紫叶兰为自墨西哥引进品种,其所在紫露草属未出现在中国种子植物属的分布类型中,余下258 个属分布结果见表2。
大岗山属于亚热带,具有过渡带的性质,群落植被也具有丰富的区系成分,其中以热带区系的植物占优势,所有热带成分分布(分布类型2~7)占种子植物属的49.35%,共128 个属,包含悬钩子属(Rubus)、木姜子属(Litsea)、山胡椒属(Lindera)等数量优势属,其中泛热带分布的属最多,共计54 属,占种子植物属的21.00%,表明植物种类上与热带植物具有密切的亲缘关系。其他分布类型中,北温带分布最多,共计37属,占14.40%,包含栎属(Quercus)、李属(Prunus)等数量优势种,东亚分布略低于北温带分布,为35 属,占13.62%,验证了大岗山所处中亚热带地区具有热带和温带的过渡地带性,此外还有9 个中国特有分布属,占3.50%,分别是伯乐树属(Bretschneidera)、喜树属(Camptotheca)、蜡梅属(Chimonanthus)、巴豆藤属(Craspedolobium)、杉木属(Cunninghamia)、青钱柳属(Cyclocarya)、箭竹属(Fargesia)、大血藤属(Sargentodoxa)、四棱草属(Schnabelia)。
乔木层、灌木层和草本层重要值排名前5 位的物种信息见表3。其中,杉木(Cunninghamialanceolata)在乔木层占据绝对优势,杉木为群落建群种和优势种,调查发现,大岗山还拥有部分杉木人工林。木荷(Schima superba)重要值排名第2,为群落次优势种,此外,在群落灌木层中,木荷也具有较高重要值,排名第4 位,证明其具有较强的更新能力。灌木层中,杜茎山(Maesajaponica)数量最多,油茶(Camellia oleifera)的出现频率最高,但综合来看,杜茎山的重要值最高,为灌木层的主要优势种。在草本层中,荩草( Arthraxon hispidus)数量最多,齿头鳞毛蕨(Dryopteris labordei)出现频率最高,但由于箭竹(Fargesia spathacea)更新能力强,长势快等生理优势,重要值排名最高,成为草本层的主要优势种。
从群落组成的种类及分布分析,有98 个种的乔木树木处于更新阶段,调查期间均在灌木层中,且乔木层中的主要树种均在林下出现幼苗,木荷和杉木更是在灌木层种占据一定优势,表明群落结构基本稳定,杉木还将保持优势地位。
2.2 物种多样性分析
大岗山植物群落灌木层拥有最高的多样性和均匀度,H、D、J 指数分别为4.46、0.97、0.98。草本层次之,H、D、J 指数分别为3.94、0.96、0.97,乔木层最低,H、D、J 指数分别为3.77、0.94、0.94。草本层与乔木层相差不大,灌木层与其他两层相差明显。整体多样性表现灌木层>草本层>乔木层,H 指数最低的乔木层也达到3.77,大岗山植物群落整体均具有较高的物种多样性。
按照各个样带划分,大岗山植物群落乔木层、灌木层和草本层的H、D、J 指数计算结果如表4。可以看出,灌木层和草本层在4 条样带上的变化相似,多样性较为稳定,4 条样带相差并不大,均匀度上3、4 号样线略高于1、2 号样线,但总体上看灌木层仍高于草本层。无论是多样性还是均匀度,乔木层在样带4 上均有明显的劣势,远低于其他样带。样带4 中接近纯林的杉木林和毛竹林在乔木层占据了绝对优势,导致样带4 乔木层整体的多样性下降。
从各样地植被层来看(图2),草本层物种丰富度最高的是47 号样地,含29 种物种,灌木层物种丰富度最高的是53 号,含57 种,乔木层物种丰富度最高的是37 号,含25 种,总体物种丰富度以46 号最多,含86 种,均不处在同一样地,这与各样地的生态环境变化有關。不同的环境对不同植被层的生物多样性有一定影影响,同时也有一定的人为因素,如44 号的荒草地,没有乔木层,以及61、62 号的杉木纯林,乔木层只有杉木,没有算在多样性计算当中。
2.3 群落相似性分析
由表5 可知,各样带之间的S?rensen 指数(SI)和Cody 指数(βC)相似性系数范围为0.50~0.66,均值为0.59,3、4 号样地相似性最低,为0.50,1、2 号样地相似性最高,为0.66,物种转换率平均为105.00,1 号样带和2 号样带物种转化率最小,为80.00;3 号和4 号样带物种转化率最大,为139.50。
3 讨论
在调查的62 个样地中,总计物种486 种,隶属120 科286 属,其中乔木层144 种,隶属52 科87 属;灌木层283 种,隶属114 科154 属;草本层182 种,隶属99 科133 属。灌木层科、属、种数均高于乔木层和草本层。在森林生态系统中灌木与乔木相互作用,影响上层乔木生长、下层幼苗更新以及草本群落结构[33]。因此,灌木层的结构和空间分布对于维持森林生态系统稳定性具有关键作用。
在整个森林群落中,蔷薇科种数最多,其次是樟科、茜草科、壳斗科、豆科。与李海静[34]对大岗山常绿阔叶林植物区系及多样性的研究相比较,当时的主要优势种和建群种是壳斗科、樟科,壳斗科和樟科仍然保持着常绿阔叶林的优势地位。此外,群落中还拥有珍贵的残遗植物和孑遗植物,如胡桃(第三纪残遗植物)、伯乐树(古老残遗植物)、篦子三尖杉(孑遗植物)等,说明了大岗山森林群落的植物起源具有古老性。
一个群落的植物区系与其周围群落的植物区系存在紧密联系,复杂的植物区系组成能够为物种之间的交流、分化、形成与演变提供更多的变异可能[35]。大岗山处于中亚热带,具有热带和温带的过渡地带性,与李海静[34]的研究相比,群落植被属的热带成分有所增加,温带成分减少,热带分布成分已经占据了接近一半的比例。由此可以看出,中亚热带的植物群落具有以亚热带植物为主,热带成分与温带成分并存,且植物区系由温带成分向热带成分过渡的特点。
各个植被层的Shannon-Weiner 多样性指数、Simpson 多样性指数和Pielous 均匀度指数均以灌木层最高,说明层间植物丰富,群落虽然已经接近顶级群落,几乎完全郁闭,但林下灌木生长以及乔木树种的更新能力依然很强,使得灌木层在物种丰富度上具有一定优势,并且对环境变化具有一定的抵抗能力与恢复能力。草本层适应能力强,也具有较高的物种多样性,群落的主体乔木层反而具有一定劣势,分析原因,密度对种子和幼苗存活率的影响及对于维持物种多样性发挥着重要作用,光照和土壤养分等可用资源的竞争也影响着物种多样性的维持[36]。
各样带之间物种多样性表现与整体相似,没有明显的区域差异,仅4 号样带的乔木层表现出劣势,分析其原因为4 号样带中存在部分杉木纯林以及荒草地影响了4 号样带整体物种多样性。各样带相似性指数范围为0.50~0.66,指数相差0.16,说明样带间差异较小,群落较为稳定。但整体相似性不高,处于中等偏上水平。从物种转化率来看,3、4 号样带物种差异较大,1、2 号样带差异较小,这与物种丰富度上的变化表现基本吻合。各样带之间形成了各自的小生境,具有一定的环境异质性,使得各样带之间的物种及物种多样性上产生了差异,但总体上环境比较稳定,人为干扰较小,群落本身较高的丰富度也对环境干扰有一定的抵御能力。
引发物种灭绝的原因有很多,但有2 种被格外注意。一种为当地特有或珍稀物种的整个栖息地被破坏,栖息地的丧失导致物种灭绝;另一种为外来物种的引入,通过引发物种间相互竞争资源,致使本地物种的丰度和分布下降甚至灭绝[37]。大岗山植物群落受国家政策保护,人为干预较小,也鲜少发生大面积自然灾害,为物种提供了稳定且安全的生存环境。丰富的物种多样性以及稳定的群落结构使得群落本身具备了抵御外来物种入侵的能力。
从历史上看,大多数森林都经历了不同程度的人类活动,植物群落受到破碎化和气候变化的影响,在区域形成小生境。人们对环境异质性如何影响群落的物种组成、结构和功能知之尚少,环境异质性又是制定生物多样性保护计划的关键因素。本研究仅根据积累的多年来有关生态因素对物种多样性和群落结构影响的研究[38-40],分析影响大岗山各植被层物种多样性间的差异,下一步将会对物种多样性的维持机制展开深入研究。探寻以不损害生物多样性的方式实施生物多样性保护及可持续利用。
