表达狂犬病病毒糖蛋白稳转细胞株的构建与应用
2023-07-08李海伦龚志远焦翠翠王福江黄静波白玉洁张海丽李媛媛王化磊
李海伦, 龚志远, 焦翠翠, 王福江,黄静波, 白玉洁, 黄 培, 张海丽, 李媛媛, 王化磊*
(1.吉林大学 动物医学学院 人兽共患病研究教育部重点实验室,吉林 长春 130062;2. 潍坊市畜牧业发展中心,山东 潍坊 262199)
狂犬病是由狂犬病病毒(rabies virus,RABV)感染引起的中枢神经系统人兽共患传染病,对宠物健康和公共卫生安全构成重大威胁。一些国家的狂犬病防控经验表明,动物疫苗免疫是控制人间狂犬病最经济有效的方法。世界卫生组织(World Health Organization,WHO)呼吁全球在2030年前消灭由犬传播的人间狂犬病。常见的动物用狂犬病疫苗主要分为弱毒活疫苗、灭活疫苗、基因工程疫苗3类。弱毒活疫苗容易毒力返强,并对免疫缺陷动物等存在潜在感染风险[1],目前在我国已经被禁止生产和使用[2],在国外主要用于野生动物的口服免疫。当前应用较为广泛的动物疫苗多为灭活疫苗,但其仍存在成本高和免疫效果不够持久的不足。因此研制安全、高效的基因工程疫苗对于狂犬病的综合防控具有重要意义。
RABV基因组编码5种结构蛋白质,分别是核蛋白(nucleoprotein,N)、磷蛋白(phosphoprotein,P)、基质蛋白(matrix protein,M)、糖蛋白(glycoprotein,G)和依赖RNA的RNA多聚酶蛋白(RNA polymerase,L)[3]。G蛋白是唯一存在于RABV粒子表面的蛋白,是RABV的主要保护性抗原,能诱导机体产生体液免疫应答和细胞免疫应答,还参与介导病毒与细胞受体的结合,在病毒致病性和嗜神经性中发挥重要作用[4-5]。G基因缺失的RABV因缺乏病毒传播或感染需要的元件,被称为“复制缺陷型”RABV。复制缺陷型RABV在细胞内能完成一次生命周期,但不能产生子代病毒,因此安全性要优于其他疫苗[6]。此外,复制缺陷型RABV可作为疫苗载体,为研制安全、高效的复制缺陷型活载体疫苗提供了新的思路。本研究拟构建稳定表达RABV-G基因的稳转细胞株,为复制缺陷型RABV的高效增殖和狂犬病新型疫苗的研制奠定基础。
1 材料与方法
1.1 主要试剂真核表达载体YHM-Cas9-SP、Piggy Bac转座子酶辅助质粒、pcDNA3.0-SRV9-G重组质粒、BSR细胞等均保存于吉林大学人兽共患病研究教育部重点实验室病毒病研究室;AgeⅠ、BsrGⅠ限制性内切酶、T4DNA连接酶均购自NEB公司;AxyPrep DNA凝胶回收试剂盒均购自Axygen公司;Primer STAR Max DNA Polymerase购自北京宝日医生物技术公司;DH5α感受态细胞、DNA Marker、SOC培养基均购自TaKaRa公司;去内毒素质粒小量提取试剂盒为QIAGEN公司产品;DMEM培养液、0.25%胰酶和胎牛血清均为Gibco公司产品;Polyjet脂质体转染试剂购自Invitrogen公司;预染蛋白Marker购自Thermo公司;细胞膜染料DIO和鼠源抗RABV-G单克隆抗体购自Merck公司;TRITC标记的山羊抗小鼠IgG购自Abcam公司;HRP标记的山羊抗鼠IgG和FITC标记的山羊抗鼠IgG购自Bio World公司;灭瘟素S盐酸盐(blasticidin)购自大连美仑生物技术有限公司;MinuteTM质膜蛋白和细胞组分分离试剂盒购自Invent公司。
1.2 RABV-G表达载体的构建
1.2.1RABV-G基因的扩增 根据GenBank中公布的RABV SRV9株(GenBank:AF499686.2)G基因序列,分别设计RABV-G基因的PCR扩增引物,上游引物F:5′-ATAACCGGTATGGTTCCTCAGGCTCTCC-3′,下游引物R:5′-ATATGTACATTACAGTCTGGTCTCACCCCC-3′。以pcDNA3.0-SRV9-G质粒为模板,通过PCR扩增RABV SRV9 G基因。PCR扩增体系:上、下游引物(10 μmol/L)各2 μL,模板200 ng,Primer STAR 25 μL,加ddH2O补充至50 μL。PCR反应条件:98℃ 1 min;98℃ 10 s,55℃ 10 s,72℃ 20 s,35个循环;72℃ 7 min。扩增完成后,产物经1%琼脂糖凝胶电泳回收目的条带。
1.2.2重组质粒的构建 将YHM-Cas9-SP载体和目的基因RABV-G用AgeⅠ、BsrGⅠ内切酶酶切,酶切产物经琼脂糖凝胶电泳鉴定,回收目的条带(酶切后YHM载体)与RABV-G目的基因按一定比例于16℃连接过夜。将连接产物转化至DH5α感受态细胞,培养12~16 h后,挑取单克隆菌落,扩大培养后提取质粒双酶切鉴定正确后送生工测序鉴定。将鉴定正确的重组质粒命名为YHM-SRV9-G。
1.3 RABV-G蛋白的表达将BSR细胞接种于96孔细胞培养板中,待细生长至底面积的70%~80%时进行转染,具体步骤:向无菌离心管中加入15 μL DMEM细胞培养液、0.5 μL Polyjet脂质体转染试剂以及0.2 μg YHM-SRV9-G后轻轻混匀,室温孵育15~20 min,然后将上述混合液缓慢加至96孔板培养的BSR细胞中,于37℃、5% CO2细胞培养箱中培养,6 h后换新鲜培养基。转染后48 h弃掉细胞上清,用80%冷丙酮室温固定30 min;弃去液体用PBST清洗3次,使用1%BSA稀释鼠抗RABV-G单克隆抗体(1∶500),37℃孵育1 h;弃去液体用PBST清洗3次,使用含1%BSA 1∶200倍稀释的FITC标记的山羊抗鼠IgG抗体(含1∶500倍稀释的伊文思蓝),37℃孵育1 h;弃去液体用PBST清洗3次,置倒置荧光显微镜下观察荧光信号。
1.4 稳转细胞株的构建及鉴定
1.4.1Blasticidin对BSR细胞的毒性测定 将处于对数生长期的BSR细胞经胰酶消化,按照3×105个/孔接种至6孔板中, 37℃、5% CO2的培养箱中培养12 h后弃掉细胞培养上清,分别换成含0,5,10,20,30 mg/L blasticidin的培养基,每天观察细胞死亡情况,适时更换含有blasticidin、10%胎牛血清的 DMEM培养基,每日观察细胞生长状态。连续培养1周后,选择能使细胞形态未发生明显变化且密度达到70%~80%的质量浓度作为最适筛选质量浓度。
1.4.2稳转细胞株的构建 将BSR细胞接种于6孔细胞培养板中,待细胞密度达70%~80%时,向无菌离心管中加入50 μL DMEM细胞培养液、5 μL Polyjet脂质体转染试剂、1 μg表达RABV-G基因的重组质粒以及1 μg Piggy Bac转座子酶质粒后轻轻混匀,在室温下孵育15~20 min,然后将上述混合液缓慢加至BSR细胞的培养液中,混匀,于37℃、5% CO2细胞培养箱中培养,6 h后换新鲜培养基。转染24 h后,加入20 mg/L的blasticidin进行药物筛选。培养2周后,对细胞板中的细胞进行有限稀释,接种于96孔细胞培养板,添加20 mg/L的blasticidin,继续对细胞进行扩增筛选,直至部分细胞生长成簇,再次利用有限稀释法进行单克隆筛选,如此经2次单克隆纯化后,获得能在blasticidin作用下稳定生长的单克隆细胞株,鉴定正确后扩大培养并冻存细胞。
1.4.3稳转细胞株的PCR鉴定 按照试剂盒说明提取稳转细胞株BSR-G和正常BSR细胞的基因组,进行PCR扩增。PCR扩增体系:上、下游引物(10 μmol/L)各2 μL,模板200 ng,Primer STAR 25 μL,加ddH2O补充到50 μL。PCR反应条件:98℃ 1 min;98℃ 10 s,55℃ 10 s,72℃ 20 s,35个循环;72℃ 7 min。
1.4.4间接免疫荧光鉴定 利用间接免疫荧光(indirect immunofluorescence assay,IFA)鉴定细胞株中BSR-G的表达情况。将BSR-G细胞以合适的细胞密度接种于96孔细胞培养板中,48 h后用80%冷丙酮室温固定30 min;弃去液体用PBST清洗3次,用含1%BSA的PBST 1∶500倍稀释的鼠抗RABV-G蛋白单克隆抗体37℃孵育1 h;用含1%BSA的PBST 1∶200倍稀释的FITC标记的山羊抗鼠IgG抗体(含1∶500倍稀释的伊文思蓝)37℃孵育1 h;弃去液体用PBST清洗3次,置于倒置荧光显微镜下观察荧光信号。
1.4.5激光共聚焦鉴定 将BSR-G细胞传代,按2×104个/孔加入已放置细胞爬片的24孔细胞培养板中,37℃温箱中培养36~48 h后用4%多聚甲醛孵育20 min固定细胞,用PBS冲洗3次,加入含1% BSA的PBS室温封闭30 min;用封闭液1∶500倍稀释鼠抗RABV-G单克隆抗体,37℃孵育1 h;用封闭液1∶500倍稀释TRITC标记的抗鼠IgG 抗体,37℃孵育1 h。用PBST洗涤3次后,用50 μmol/L 的细胞膜绿色荧光探针(DIO)37℃孵育15 min;用PBST洗涤3次后,将含DAPI的抗荧光淬灭剂滴于爬片中央,3~5 min后,用90%甘油封片,使用激光共聚焦显微镜进行观察。
1.4.6Western blot鉴定 将BSR-G细胞传代,按MinuteTM质膜蛋白和细胞组分分离试剂盒说明书提取细胞膜成分,用裂解液进行裂解,经 5×loading buffer 处理后进行10% SDS-PAGE,电泳后将蛋白转移至NC膜,加入用封闭液1∶500倍稀释的鼠抗RABV-G蛋白单克隆抗体37℃孵育1.5 h,加入使用封闭液1∶10 000倍稀释的HRP标记羊抗鼠IgG(H+L),37℃孵育1 h,PBST冲洗3次后,加入显色液进行显影。
1.5 稳转细胞株遗传稳定性的鉴定为了验证BSR-G细胞株中RABV-G蛋白表达的稳定性,将细胞进行连续传代培养, F2、F8及F15代细胞传代时留取部分细胞进行IFA鉴定。
1.6 G基因缺失重组狂犬病病毒rSRV9-△G-eGFP在稳转细胞株中的增殖将BSR-G细胞和正常BSR细胞按合适的密度接种至24孔板,500 μL/孔。将G基因缺失的重组病毒rSRV9-△G-eGFP按MOI = 0.2分别接种单层BSR-G细胞和单层BSR细胞,37℃、5% CO2培养箱培养,分别于病毒感染后每24 h 通过重组病毒带有的绿色荧光信号蛋白eGFP的表达来观察病毒在稳转细胞株和正常细胞中的增殖情况。每24 h收取1次病毒,测定病毒半数细胞感染量(50% tissue culture infectious dose,TCID50),并绘制病毒增殖曲线。
2 结果
2.1 重组质粒的鉴定RABV-G的PCR扩增产物经琼脂糖凝胶电泳,得到1 575 bp特异性条带,与预期大小一致。将RABV-G克隆至YHM载体,用AgeⅠ、BsrGⅠ内切酶进行酶切鉴定。酶切产物经1%琼脂糖凝胶电泳分析后,得到7 173 bp的载体片段和1 575 bp的目的基因条带,与预期大小一致(图1)。测序结果显示,基因片段与目的基因序列一致,表明成功构建了含有RABV-G基因的重组质粒,命名为YHM-SRV9-G。
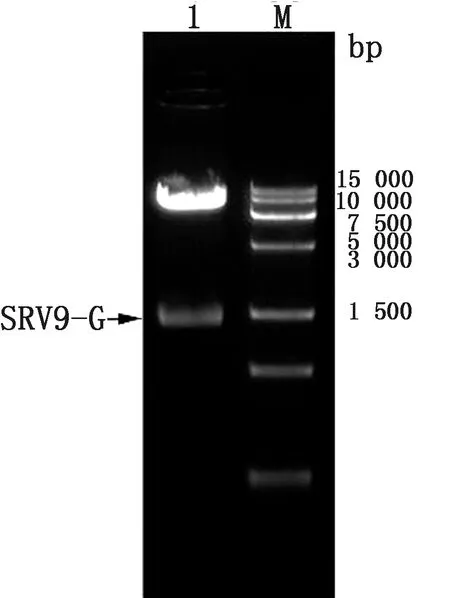
M.Marker;1.YHM-SRV9-G质粒的双酶切产物
2.2 RABV-G蛋白的表达为鉴定G蛋白是否表达,将重组质粒YHM-SRV9-G、空载体质粒分别转染BSR细胞,转染后48 h以鼠抗RABV-G单克隆抗体作为一抗进行IFA鉴定。结果显示,转染YHM-SRV9-G质粒的细胞可检到特异性荧光,表明重组质粒YHM-SRV9-G可在细胞中成功表达RABV-G蛋白(图2)。
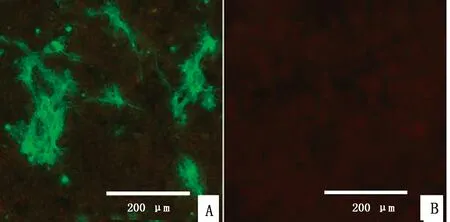
A.重组质粒YHM-SRV9-G;B.空载体质粒YHM-spCas9
2.3 稳转细胞株的筛选及鉴定使用含不同质量浓度blasticidin的培养液培养BSR细胞,通过显微镜观察细胞的生长状态(图3),培养1周后,blasticidin质量浓度为20 mg/L时细胞仍可达到70%~80%密度,且细胞形态未发生改变,因此确定药物筛选最适质量浓度为20 mg/L。将构建的重组质粒YHM-SRV9-G与Piggy Bac转座子酶辅助质粒共转染BSR细胞,经blasticidin药筛及单克隆纯化后获得稳转细胞株。分别选取单克隆传代1,8次,同时提取细胞基因组后进行PCR鉴定,结果显示,细胞株在大约1 575 bp处出现目的条带,对照组无条带,证明RABV-G基因成功插入细胞基因组中(图4)。选取F3代细胞株进行IFA鉴定,结果显示,稳转细胞株可在荧光显微镜下观察到明亮且均一的绿色荧光,证明单克隆纯化后获得的细胞株可以高效表达RABV-G蛋白(图5)。为进一步确定RABV-G在细胞中的表达位置,通过激光共聚焦显微镜进行鉴定(图6),在稳转细胞株的细胞膜上可观察到均匀分布的红色荧光,而且红色荧光信号与商品化的细胞膜染料DIO有很好的共定位,证明RABV-G主要在细胞膜上表达。选取F8代细胞提取膜蛋白进行Western blot鉴定,结果显示,细胞株可在约66 kDa处检测到目的蛋白表达,与RABV-G蛋白大小相符,而BSR细胞对照没有检测到相应的特异性条带(图7),证明RABV-G正确表达且定位于细胞膜。综上结果,表明成功构建了表达RABV-G的稳转细胞株BSR-G。
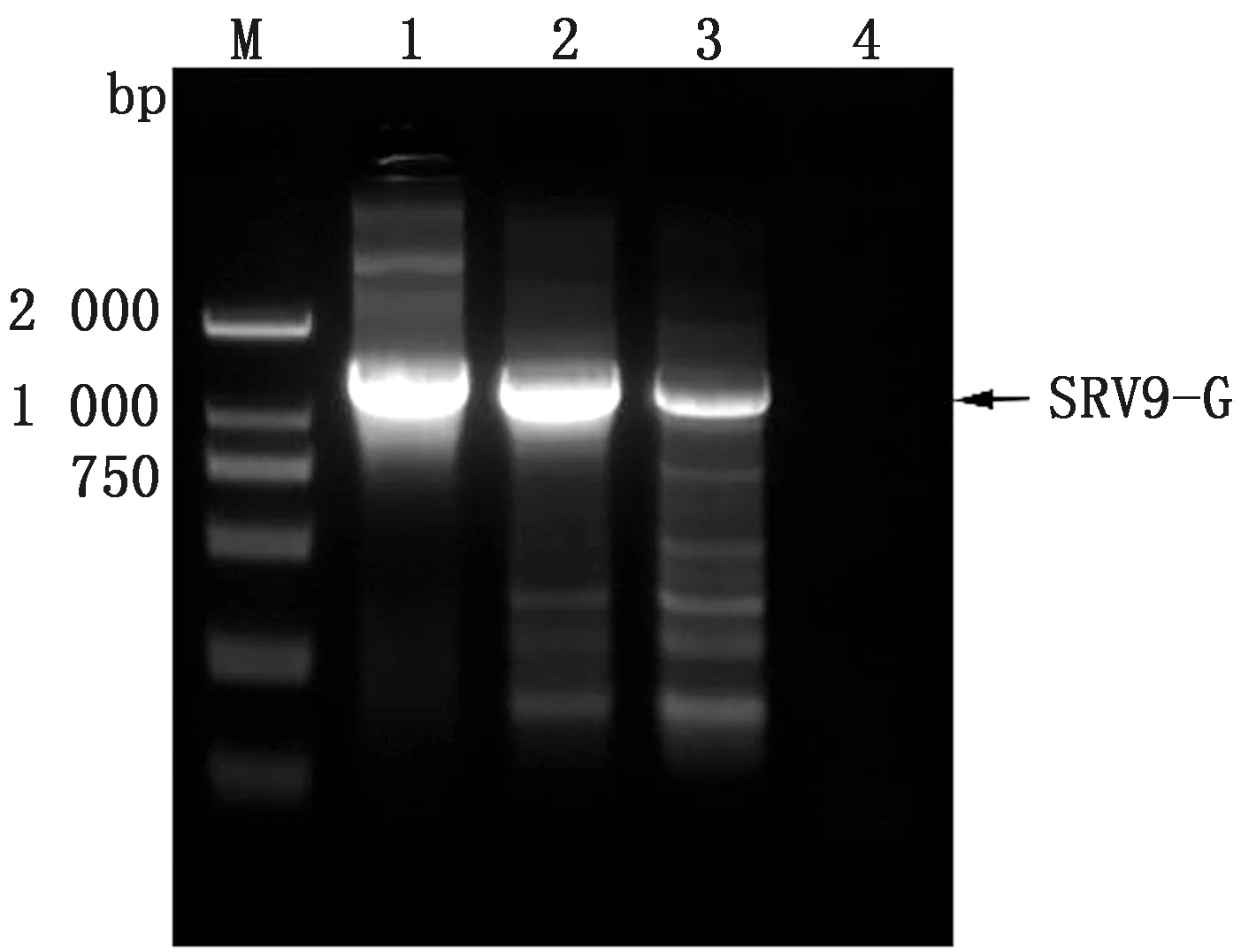
M.DL2000 DNA Marker;1.阳性对照;2.F8代BSR-G细胞;3.F1代BSR-G细胞;4.阴性对照
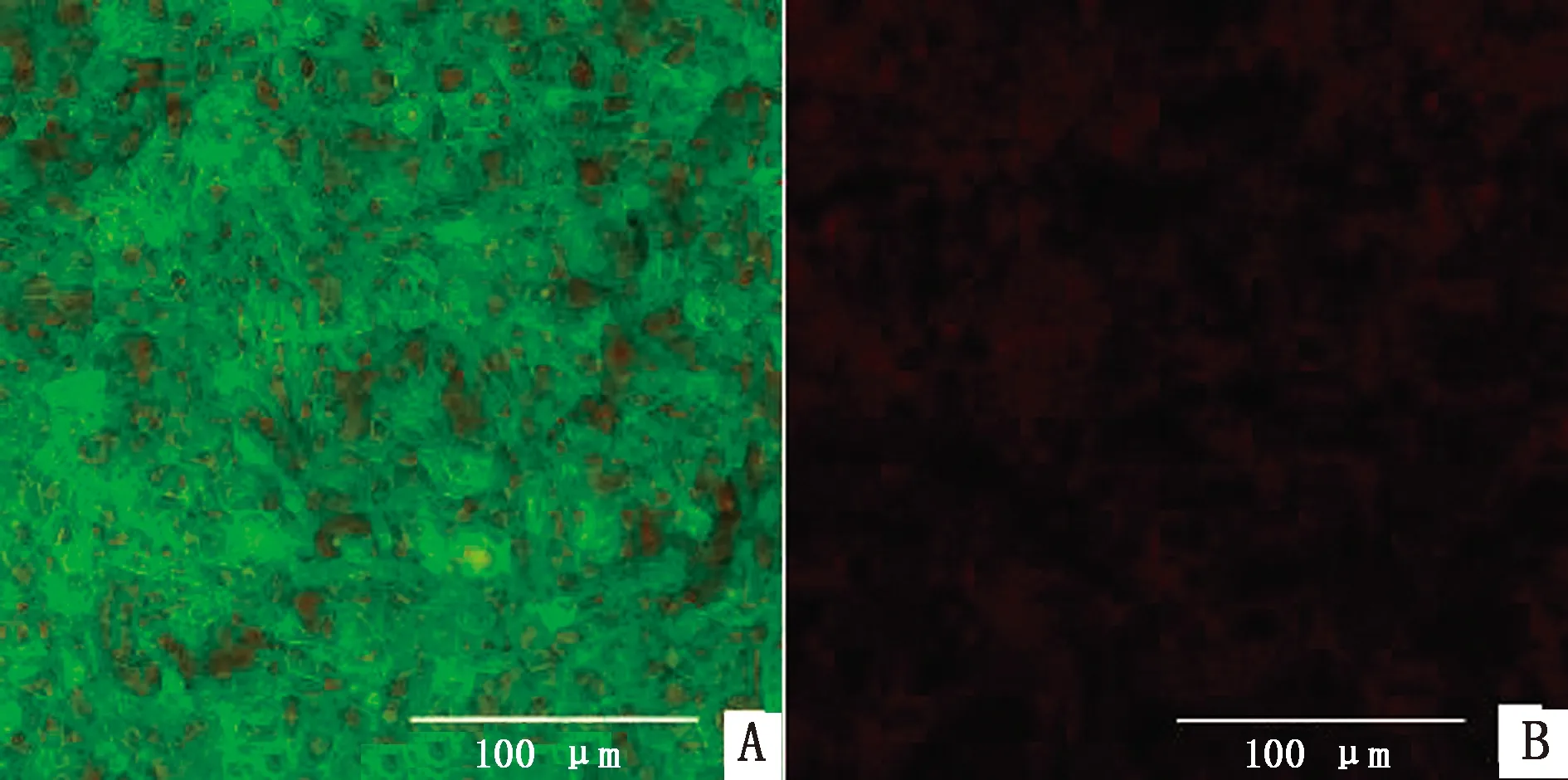
A.BSR-G细胞;B.正常BSR细胞
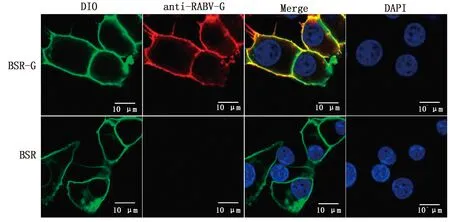
图6 稳转细胞株RABV-G蛋白表达位置鉴定
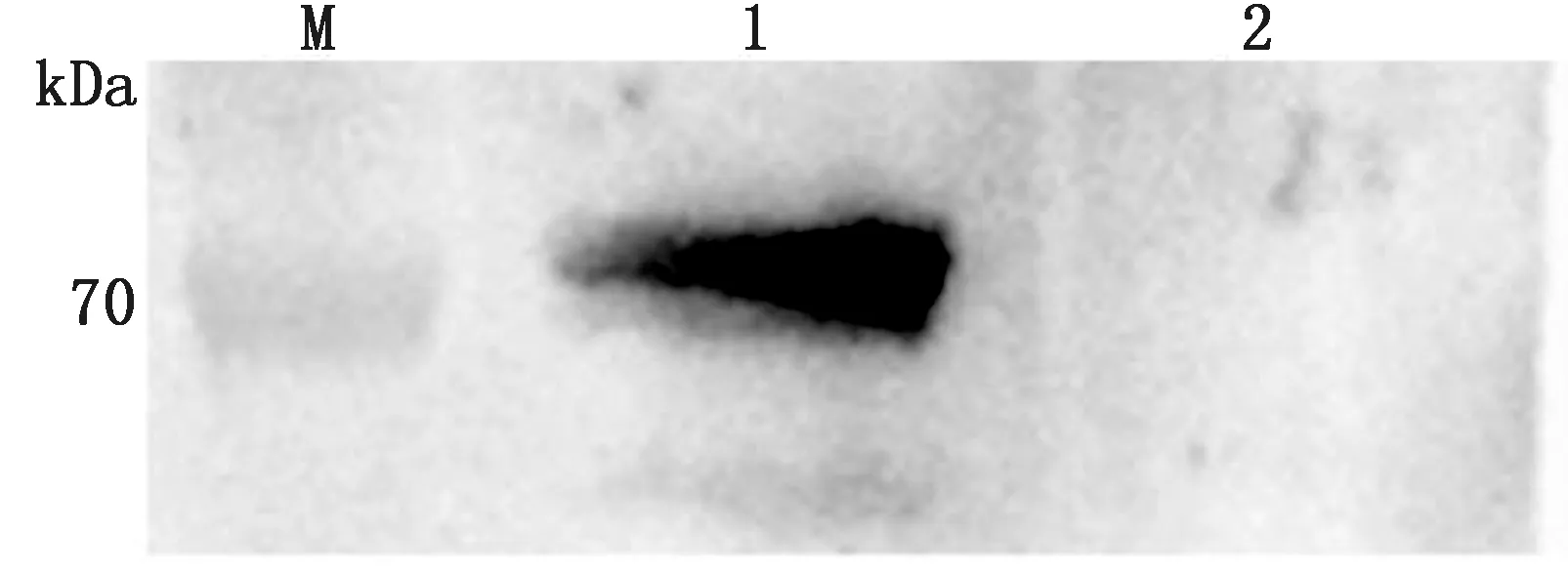
M.蛋白Marker;1.BSR-G细胞;2.BSR细胞
2.4 稳转细胞株的遗传稳定性为了验证稳转细胞株BSR-G中 RABV-G蛋白表达的稳定性,选取F2、F8、F15代次细胞,利用IFA进行检测,结果显示,该细胞株从F2至F15代都能够稳定表达RABV-G蛋白,证明该细胞株能够稳定持续表达外源RABV-G蛋白(图8)。

A.F2代BSR-G细胞;B.F8代BSR-G细胞;C.F15代BSR-G细胞
2.5 复制缺陷型重组狂犬病病毒在稳转细胞株中的增殖将G基因缺失的重组病毒rSRV9-△G-eGFP按MOI=0.2分别单层接种BSR-G细胞和BSR细胞,通过观察绿色荧光的表达评价重组病毒的增殖情况。结果显示,rSRV9-△G-eGFP感染BSR-G细胞后可出现明显增殖趋势,而感染正常BSR细胞后则没有增殖(图9)。每24 h收集病毒感染BSR-G细胞上清培养物,进行病毒滴度测定,结果显示,重组病毒rSRV9-△G-eGFP可在BSR-G细胞增殖,且病毒滴度可达105.3TCID50/mL;感染BSR细胞后,病毒滴度低于可检测值(图10)。由此表明,BSR-G细胞可用于复制缺陷型RABV的增殖培养。

图9 重组病毒rSRV9-△G-eGFP的增殖情况

图10 重组病毒rSRV9-△G-eGFP在BSR-G细胞和BSR细胞上的生长曲线
3 讨论
疫苗免疫是控制传染病暴发和流行的主要措施。传统疫苗主要包括灭活疫苗和弱毒疫苗。灭活疫苗引起的细胞免疫较弱,持续时间短,以及各个抗原成分之间的免疫应答不平衡,往往需要多次免疫才能产生较好的免疫应答;弱毒疫苗存在毒力返强以及散毒的风险,限制了其在临床的进一步应用[7]。针对新发、突发传染病,积极采用新技术、新手段研发安全、高效的新型疫苗才能防患于未然[8]。复制缺陷型病毒疫苗不仅能诱导宿主产生体液免疫和细胞免疫,还具有良好的安全性。本研究构建了一种可用于复制缺陷型重组RABV增殖培养的稳转细胞株,为狂犬病复制缺陷基因工程疫苗的研制奠定了基础。
稳定表达外源蛋白的细胞系在基因和蛋白功能等生物学研究中具有重要作用。ITO等[9]构建了一种M基因缺陷的RABV RC-HL△M,该病毒仅在表达M蛋白的BHK-M细胞株上增殖,将RC-HL△M通过滴鼻免疫小鼠后诱导产生的中和抗体水平与RC-HL株相同。BLANEY等[10]和PAPANERI等[11]基于RABV反向遗传操作系统,构建了表达Ebola GP的RABV G基因缺失的重组病毒BNSP-△G-GP,重组病毒BNSP-△G-GP可在表达RABV-G的BSR-RVG细胞系上复制增殖,在正常BSR细胞上则仅能进行单轮复制。动物试验结果表明重组病毒BNSP-△G-GP可作为RABV和Ebola二联疫苗候选株。KATO等[12]开发的表达MERS-S1的RABV-△P RABV疫苗,同样借助表达RABV P蛋白的BHK-P细胞系进行疫苗候选株的增殖培养。本研究中,G基因缺失的重组病毒rSRV9-△G-eGFP感染BSR-G细胞后可成功增殖,但在BSR细胞中仅能进行单轮复制,表明rSRV9-△G-eGFP具有良好的安全性,可作为复制缺陷疫苗候选株。
构建稳转细胞株(stable-transfection cell line)的基本原理是将外源DNA克隆到具有某种抗性的载体上,重组载体被转染到宿主细胞后定点或随机整合到宿主细胞基因组中,之后根据载体所含有的抗性标志选用相应的药物进行筛选,筛选得到可以稳定表达目的蛋白或者稳定表达特定基因的细胞株。建立稳定表达外源基因的细胞系有多种方法,常见的有质粒转染、慢病毒感染等方式。利用慢病毒载体构建稳转细胞系的方法除了需要构建所需的质粒,还需包装并获得高滴度的病毒,操作过程复杂繁琐,且在操作过程中还要注意生物安全问题,而且慢病毒启动外源基因表达的时间较长[13]。pcDNA3.1载体是构建真核细胞系传统的载体之一,但是此载体缺少细胞基因组整合元件,所以用其构建细胞系的效率较低,而且在稳转细胞的后期传代过程中,常出现外源基因丢失的情况[14]。相比之前2种方法,转座系统具有高容纳量、高效率以及高安全性等优势,而且转座系统中的辅助质粒表达的转座酶使外源基因可稳定插入细胞基因组上,在很大程度上降低外源基因丢失的可能性[15]。本试验选用了在哺乳动物细胞中转座活性更高的Piggy Bac转座子系统[16]。Piggy Bac转座子是可以在原核和真核生物基因组上进行基因位置改变的DNA片段,通过“剪切-粘贴”的机制在转座子和染色体之间进行基因换位。Piggy Bac转座酶识别转座子载体两端的反向重复序列(inverted terminal repeat sequences,ITRS)并将其插入宿主基因组的TTAA位点[17-18]。比较于传统的转座子系统,其有更高的整合效率,更高的目的基因表达效率。此外,也能容纳更大目的片段的插入。与Sleeping Beauty转座子系统相比,Piggy Bac转座子具有更高的转座活性[15-16]。此外,Sleeping Beauty转座后会留下 3 bp的基因转移足迹CAC,而Piggy Bac转座后不会留下基因序列改变痕迹[19-22]。丁晟等[23]将Piggy Bac转座子成功应用于小鼠和哺乳动物细胞基因功能的研究,实现了Piggy Bac转座子在人和小鼠细胞中高效导入外源基因并稳定表达,并成功培育出了带荧光的转基因小鼠。LYNCH等[24]利用Piggy Bac转座系统构建了能持续表达人类免疫缺陷型病毒Ⅰ型(human immunodeficiency virus type 1,HIV1)Gag蛋白的转基因昆虫细胞系,用于HIV-1 Gag病毒样颗粒的制备。本研究构建的稳定表达RABV-G蛋白的细胞株BSR-G,连续进行15次传代后仍可稳定表达RABV-G蛋白,表明BSR-G细胞具有良好的遗传稳定性。
本研究利用Piggy Bac转座子真核表达载体构建了表达RABV-G基因的重组表达质粒,并通过与辅助质粒Piggy Bac转座酶的共转染将RABV-G基因插入到细胞基因组,通过筛选单克隆的方法获得稳定表达RABV-G蛋白的细胞株。将构建的稳转细胞株应用于复制缺陷型重组RABV的增殖培养,使复制缺陷型重组RABV利用细胞表达的糖蛋白完成病毒生命周期,组装出具有单次感染性的复制缺陷型病毒颗粒,并在稳转细胞株上实现高效增殖。综上所述,本研究构建的稳定表达RABV-G蛋白的BSR-G细胞株,可用于复制缺陷型重组RABV的拯救和增殖,为狂犬病新型疫苗和基于RABV载体的复制缺陷型基因工程疫苗的研制奠定了基础。
