球药隔重楼的种下分类
2023-07-05任梓萱赵家雯王庆鹤尹鸿翔
任梓萱 赵 灵 赵家雯 王庆鹤 尹鸿翔*
(1.成都中医药大学民族医药学院,成都 611137;2.四川省药品检验研究院,成都 611731;3.成都中医药大学医学技术学院,成都 611137)
重楼属(Paris)为广义百合科(Liliaceae)植物,全世界约有40 余种,分布于欧亚大陆的温带与亚热带地区,我国西南地区是重楼属植物的地理与多样性分布中心[1-2]。球药隔重楼(Paris fargesiiFranch.)隶属于侧膜胎座亚属(SubgenusDaiswa(Raf.)H.Li),与 蚤 休 组(Sect.Euthyra(Salisb.)Franch.)近缘[3],具有雄蕊药隔凸出部分(简称“药凸”)极短,顶面观呈球状或马蹄状膨大的特征,广泛分布于我国秦岭以南,海南以北的地区[1,4]。
一直以来,球药隔重楼种下分类较为混乱,目前主要存在3 种观点:(1)《中国植物志》[5]与Flora of China[6]将球药隔重楼划归到南重楼组(Sect.Eu⁃thya(Salisb.)Franch.)中,并记载具柄重楼(P.far⁃gesiivar.petiolata)为其唯一变种。(2)在《重楼属植物》[1]中,李恒系统根据内轮花被片形态特征差异,确立了宽瓣球药隔重楼(P.fargesiivar.latipetala)、短瓣球药隔重楼(P.fargesiivar.brevipetalata)2 个变种,与球药隔重楼原变种(P.fargesiivar.farge⁃sii)共同构成球药隔组(Sect.FargesianaeH.Li),而具柄重楼则被作为卵叶重楼(P.delavayivar.petio⁃lata)的异名,置于金线重楼(P.delavayi)种下。(3)A Monography of Paris(Melanthiaceae)[7]则认为球药隔重楼种下内轮花被片的长短、宽窄变异幅度交叉重叠,不足以作为划分变种的依据,因此将宽瓣球药隔重楼、短瓣球药隔重楼均作为球药隔重楼的异名处理。
除上述已知变种外,近年来还有更多新类群被陆续发现。2017 年,赵灵等[8]于湖南省双牌县发现了球药隔重楼一新变种,大萼球药隔重楼(P.fargesiivar.macrosepala)。2018 年,在福建省武夷山发现了球药隔重楼的又一新类群(暂称为“球药隔黑斑种群”),之后此类群又陆续在福建省三明[9]、江西省井冈山、贵州省都匀等地被发现。
显然,球药隔重楼的种下分类还存在着较多问题:球药隔重楼种下类群应细分还是合并?仅依据内轮花被片特征进行各变种的划分是否合理?具柄重楼应归属于球药隔重楼还是金线重楼种下?本研究将上述各类群以及近年来发现的球药隔重楼新类群,全部引种至四川省重楼种质资源圃。经过观测发现,它们的形态特征、物候期及甾体皂苷等均存在各自的特点,这很可能是其种下分类的重要支撑依据。基于此,本研究将通过对上述类群的形态特征、物候期特点、甾体皂苷构成、内转录间隔区(ITS)序列和地理分布位置进行系统的梳理和对比,为球药隔重楼的分类研究提供新的证据。
1 材料与方法
1.1 仪器与材料
1.1.1 仪器
高效液相色谱仪(Agilent 1200,美国)、色谱柱(250.0 mm×4.6 mm 5 μm,Cosmosil Cholester,江苏)、超纯水机(UPT-II-10T,四川)、测序仪(3730 XL,美国)、2720 thermal cycler PCR 仪(ABI 2720,美国)、电泳仪(JY 300C,北京)、凝胶成像仪(JY 04S-3C,北京)、离心机(L550,湖南)等。
1.1.2 试验地与试验材料
各类群植株均由课题组自四川、贵州、湖南、江西等地收集(见表1),并由成都中医药大学尹鸿翔副教授鉴定,统一移栽至四川省重楼种质资源圃(30°46′N,103°21′E)。该地区海拔1 300 m,属亚热带湿润季风气候,全年平均气温14 ℃,年平均日照时间641 h,历史最低温−4 ℃,年平均降水量1 012.4 mm,凭证标本保存于成都中医药大学中药标本馆(CDCM)。

表1 样品信息Table 1 Samples information
移栽的植株适应性栽培1 a 后,选取生长状态良好、每年稳定开花结实的成熟植株进行为期3 a的田间观察。观察结束后,取根茎干燥用于测定甾体皂苷(样品信息见表1),取新鲜叶片用于提取DNA 测定ITS 序列,其余重楼属植物ITS 序列均由GenBank获取(见附表1)。
重楼皂苷Ⅴ(纯度97.13%),重楼皂苷H(纯度98.31%)由课题组按照汪瑶等[10]方法制备,重楼皂苷Ⅰ(批号111590-201604;纯度93.60%),重楼皂苷Ⅱ(批号111591-201103;纯度93.40%),重楼皂苷Ⅵ(批号111592-20160;纯度98.00%),重楼皂苷Ⅶ(批号111593-201604;纯度94.00%)购自中国食品药品检定研究院。
1.2 研究方法
1.2.1 形态学研究
对每个类群的形态特征进行为期3 a 的田间观察,并结合野外调查,同时参考《中国植物志》[5]、《中 国 高 等 植 物 图 鉴》[10]、《重 楼 属 植 物》[1]、A Monography of Paris(Melanthiaceae)[7]等 文 献 资料,以及中国数字植物标本馆(https://www.cvh.ac.cn/)的标本资源,进行形态学的对比与整理。
1.2.2 物候期观察
每个类群选择20株健康、长势一致的个体,进行挂牌标记,并观察记录1 a 中出苗、展叶、开花、果裂和倒苗5个生长阶段的具体日期,进行物候期特征分析。
1.2.3 植物化学分类研究
选择重楼属植物常见的6种甾体皂苷:重楼皂苷Ⅰ、重楼皂苷Ⅱ、重楼皂苷Ⅴ、重楼皂苷Ⅵ、重楼皂苷Ⅶ、重楼皂苷H 作为研究的特征性化学成分,按照汪瑶等[11]的方法测定30批样品中各甾体皂苷成分的组成特征,并将所测得数据导入SPSS 23.0软件,选择组间连接的聚类方法,基于平方欧氏距离,建立聚类谱系图。
1.2.4 ITS序列分析
采用植物基因组DNA 提取试剂盒提取表1 中7个类群叶片的DNA,送由成都擎科梓熙生物技术有限公司,按照张开元等[12]的方法进行PCR 扩增及序列测定,并将相关序列上传至GenBank 数据库获取登录号。利用MEGA-X 分析ITS 序列变异位点与遗传距离,采用K2P(Kimura 2-parameter)模型构建NJ(Neighbor-Joining)树,采用K2+G 最佳模型构建ML(Maximum Likelihood)树,设置Bootstrap 1 000 次重复检验各分支支持率,合并支持率<50%的分支。
1.2.5 地理分布研究
通过在四川、湖南、贵州、江西、福建、云南等地的实地调查,同时结合文献资料,对球药隔重楼种下类群的地理分布位置做出补充,并绘制地理分布图。
2 结果与分析
2.1 形态特征
对7 个重楼类群进行连续3 a 的田间定点观察,考察其形态的特异性、一致性和稳定性。结果表明各类群的形态特征区分明显,种群内部稳定一致,并没有因为迁地引种,年限增长而发生改变,形态特征整理如图1~2、表2所示。
2.1.1 球药隔重楼已知类群形态特征
观察结果表明,球药隔组3种重楼不只在内轮花被片上表现出形态差异。首先,球药隔重楼原变种的雄蕊在整个开花过程中始终保持直立、与柱头疏离的特征(如图1A)。而宽瓣球药隔重楼与短瓣球药隔重楼的雄蕊在整个开花过程中弯曲程度较大,在开花始期和开花盛期时会向内包裹住柱头(如图1B~C)。
其次,球药隔重楼外轮花被片的颜色也存在变异,少部分原变种与宽瓣球药隔重楼外轮花被片基部位置都存在一定的紫黑色晕染,但面积较小,并不明显。而在短瓣球药隔重楼中,大部分植株都存在这种现象,部分植株甚至从基部开始逐渐蔓延,使整个外轮花被片呈现出紫黑色或紫绿色(如图2C)。

图2 球药隔重楼种下类群外轮花被片形态A.基部绿色(球药隔重楼原变种);B.基部稍具紫黑色(短瓣球药隔重楼);C.全部紫绿色(短瓣球药隔重楼);D.先端具紫黑色斑块(球药隔黑斑种群)Fig.2 Comparison of morphology of outer tepals of P.fargesii A.Base green(P. fargesii var.fargesii);B.Base slightly purplish black(P. fargesii var.brevipetalata);C.Completely purplish green(P. fargesii var.brevipetalata);D.Apex with purplish black patch(Black patch population)
具柄重楼形态则与3种球药隔重楼差别较大,其药隔突出部分并未像球药隔一样膨大,而是呈尖头状;花基数3~7,相较于球药隔重楼范围更大,这些特征都更加接近于金线重楼(如图1G~H)。此外,课题组在野外考察时,曾在贵州纳雍、四川宝兴均发现了具柄重楼与金线重楼的过渡类型(如图3H),其外轮花被片绿色并未反折,表现为具柄重楼的特征;内轮花被片呈紫色,药凸较长,表现为金线重楼的特征。

图3 球药隔重楼及金线重楼种下类群花被片形态A.球药隔重楼原变种;B.宽瓣球药隔重楼;C.短瓣球药隔重楼;D.大萼球药隔重楼;E.球药隔黑斑种群;F.具柄重楼;G.金线重楼;H.具柄重楼与金线重楼过渡类型;I.外轮花被片大小对比(I-1.大萼球药隔重楼、I-2.球药隔重楼原变种,I-3.宽瓣球药隔重楼、I-4.短瓣球药隔重楼)Fig.3 Comparison of morphology of tepals between P.fargesii and P. delavayi A.P. fargesii var.fargesii;B.P. fargesii var.latipetala;C.P. fargesii var.brevipetalata;D.P. fargesii var.macrosepala;E.Black patch population;F.P. fargesii var.petiolata;G.P. delavayi var.delavayi;H.Transition type of P. fargesii var.petiolata and P. delavayi var.delavayi;I.Outer tepals size contras(tI-1.P. fargesii var.macrosepala,I-2.P. fargesii var.fargesii,I-3.P. fargesii var.latipetala,I-4.P. fargesii var.brevipetalata)
2.1.2 球药隔重楼新类群形态特征
新变种大萼球药隔重楼具有极为宽大的外轮花被片(图3:I-1),叶−萼长度比(外轮花被片与叶片之比)为1.69~2.30,宽度比为2.41~3.36,远小于其他球药隔重楼;同时其药隔突出部分形态独特,为中间具明显裂痕的蝶形与双肾形态(如图1E~F)。
球药隔黑斑种群主要特征为外轮花被片正面近先端处有一块明显的紫黑色斑块(如图2D),极易区分辨认。少部分球药隔重楼外轮花被片也具紫黑色的现象,但与之不同的是,已知的球药隔重楼各类群仅从基部开始呈现紫黑色晕染,并没有出现规则的色斑。而新类群则具有界限明显的紫黑色斑块,位于近先端1/3处的位置。通过进一步观察发现:在整个花期中,其雄蕊常排列成规则的三角形或者矩形,与柱头疏离,药隔突出部分为扁平的纽扣形状(如图1D),且花基数较小,多为3~4。
2.2 物候期特征
根据种质资源圃连续3年以来的田间观测,总结出7个类群重楼的物候期特征(见表3)。结果显示,具柄重楼的花期接近5月,倒苗期为12月,与金线重楼接近。大萼球药隔重楼物候期比其他类群提前1个月,是所有球药隔重楼中开花和倒苗最早的类型。宽瓣球药隔重楼倒苗时间最晚,地面植株可在接近0 ℃的环境下自然越冬,翌年3月才倒苗,是抗寒能力最强的类型。球药隔重楼原变种、短瓣球药隔重楼、球药隔黑斑种群的物候期大致相同。

表3 球药隔重楼及金线重楼种下类群物候期特征Table 3 Phenological characteristics of P.fargesii and P. delavayi
2.3 甾体皂苷构成
测得各类群根茎的甾体皂苷组成及含量如表4 所示,从表中可以看出,具柄重楼与金线重楼因不含重楼皂苷Ⅵ而与球药隔重楼类群相区分,同时,两者6 种重楼皂苷总含量都<1%,明显低于球药隔重楼。球药隔原变种、大萼球药隔重楼、球药隔黑斑种群主要含偏诺皂苷类(重楼皂苷Ⅶ、H、Ⅵ)成分;宽瓣球药隔重楼、短瓣球药隔重楼则同时含有偏诺皂苷与薯蓣皂苷类(重楼皂苷Ⅱ、Ⅰ、Ⅴ)成分。

表4 6种甾体皂苷含量测定结果Table 4 Determination results of six steroidal saponins
聚类结果显示(如图4),在欧式距离为4时样品被分为了6类,其中具柄重楼与金线重楼聚为一类;宽瓣球药隔重楼自身聚为一类;球药隔重楼原变种、大萼球药隔重楼与球药隔黑斑种群聚为2类;短瓣球药隔重楼虽然被分成了2类,但也在欧式距离大于4时聚在了一起,且距离其他支系较远。

图4 基于6种甾体皂苷含量的聚类谱系YB.球药隔重楼原变种;KB.宽瓣球药隔重楼;DB.短瓣球药隔重楼;DE.大萼球药隔重楼;HB.球药隔黑斑种群;JB.具柄重楼;JX.金线重楼Fig.4 The clustering dendrogram based on the content of six steroidal saponins YB.P. fargesii var.fargesii;KB.P. fargesii var.latipetala;DB.P. fargesii var.brevipetalata;DE.P. fargesii var.macrosepala;HB.Black patch popula⁃tion;JB.P. fargesii var.petiolata;JX.P. delavayi var.delavayi
2.4 ITS序列分析
2.4.1 变异位点分析
将表2中球药隔重楼、金线重楼等22条ITS序列进行比对分析。结果显示22 条序列总长度为623~625 bp,GC 含量为53.9%~54.3%,共有40 个变异位点,包含信息位点28 个。其中ITS1 序列GC平均含量51.4%,变异位点20个,信息位点15个;5.8S 序列GC 平均含量49.6%,变异位点4 个,信息位点3 个;ITS2 序列GC 平均含量54.2%,变异位点16 个,信息位点10 个。球药隔重楼变异位点主要集中在ITS1 区域,金线重楼与具柄重楼变异位点位置大致相同,变异位点详细信息见图5。

图5 球药隔重楼及金线重楼种下类群变异位点分析Fig.5 Analysis of variation loci of P.fargesii and P. delavayi
2.4.2 遗传距离计算
根据K2P模型计算的遗传距离见表5。具柄重楼与金线重楼平均遗传距离(0.003 3)远小于与其余5种球药隔重楼之间的平均遗传距离(0.025 5)。宽瓣、短瓣、大萼球药隔重楼种间最小遗传距离为0.006 5,种内最大遗传距离为0;球药隔黑斑种群种间最小遗传距离为0.008 2,种内最大遗传距离为0;球药隔重楼原变种的种间平均遗传距离为0.018 1,种内平均遗传距离为0.009 8,以上结果表明5种球药隔重楼可以相互区分。
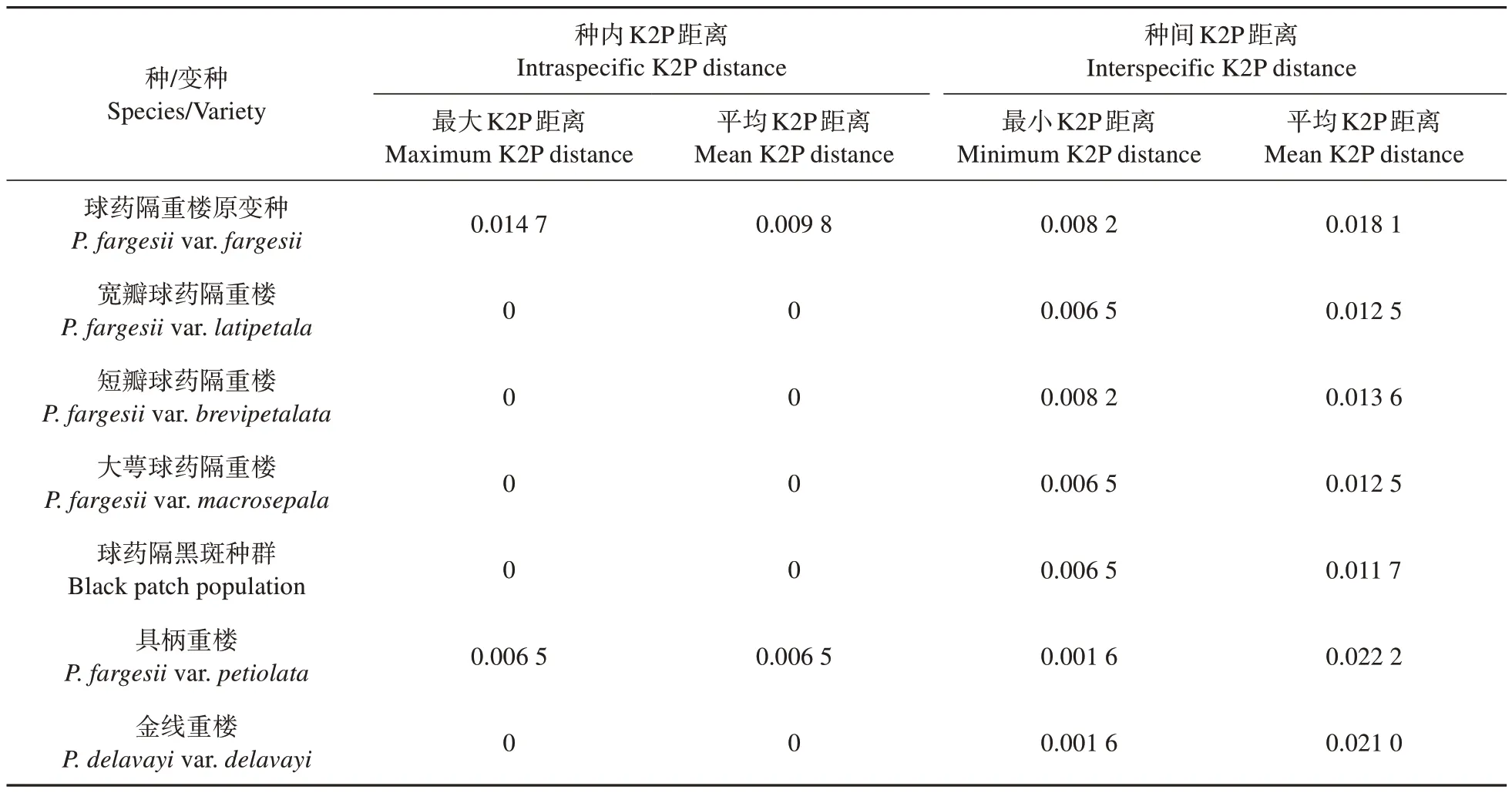
表5 球药隔重楼及金线重楼种下类群K2P距离Table 5 K2P distance of P.fargesii and P. delavayi
2.4.3 系统发育分析
重楼属植物的ITS 序列在重楼的分类鉴定中具有准确性好、鉴定效率高的特点[13-15],因此本研究基于ITS 全序列,选择所研究的7 种重楼及同属近缘植物进行系统发育分析,以在整个重楼属框架下阐明各变种的系统位置。NJ 树结果显示(见图6A),球药隔重楼原变种与大萼球药隔重楼各自聚为一支后相聚,支持率分别为87%、63%;短瓣球药隔重楼、球药隔黑斑种群、宽瓣球药隔重楼各自聚为一支,支持率分别为86%、95%、85%;而具柄重楼与金线重楼以52%的支持率相聚,且在重楼属中距离球药隔支系较远。ML 树(见图6B)与NJ 树的结果高度一致,且球药隔、金线重楼各分支支持率均高于NJ树。

图6 基于ITS序列的重楼属NJ(A)与ML(B)系统发育树Fig.6 Phylogenetic trees of NJ(A)and ML(B)based on ITS sequence of Paris L.
2.5 地理分布范围
地理分布结果显示,7种重楼主要分布在我国秦岭以南的西部地区,其中四川盆地周边山区的种群密度较大,为重楼的多样性分布中心。短瓣球药隔重楼分布范围最广,主要分布在重庆、贵州、四川东部、湖北西部、广西北部等地,几乎包含了除具柄重楼外所有球药隔重楼的分布位置,并且也是唯一分布于台湾省的球药隔重楼;球药隔重楼原变种零星分布在贵州、重庆、湖北等地,种群数量远远不如短瓣球药隔重楼;宽瓣球药隔重楼仅在四川屏山,贵州贵定、都匀,湖北鹤丰、恩施等地少量分布。可以看出,3个变种皆具有各自的范围,而不是相互混杂的均匀分布。其中新类群大萼球药隔重楼分布于湖南双牌,球药隔黑斑种群主要分布于江西、福建,二者均分布在球药隔重楼已知地理分布范围的边缘地带(图7)。

图7 球药隔重楼与金线重楼种下类群地理分布范围Fig.7 Geographic distribution of P. fargesii and P. delavayi
具柄重楼主要集中分布在四川、贵州、重庆、湖北与陕西,相对于短瓣球药隔重楼偏向北方。但实际分布范围比短瓣球药隔重楼更大,在云南西部、广西南部、安徽南部亦有分布,只是分布点位相对较少。金线重楼主要分布位置更偏西北,从云南西部一直到湖北东部均有分布,但偏南方向的贵州、湖南、广西则分布较少。
3 讨论
3.1 根据内轮花被片划分球药隔组的合理性
种内变异是植物进化过程中普遍存在的现象,植物种下类群的划分一直以来都存在着各种不同的观点。一部分学者倾向于利用更加精确合理的术语去划分种下的等级,而另一部分学者却放弃了细致的划分,认为种内的变异并没有一个明确的界限[16],球药隔重楼也是如此,李恒系统根据内轮花被片的变异进而确立了2 个变种,而A Monography of Paris(Melanthiaceae)却描述内轮花被片变异是属于正常的变化范围之内,应当作为一个种内变异进行合并描述。观察发现,原变种内轮花被片长度在4.0~8.0 cm,与另两种重楼的长度范围并没有互相重叠;同样,宽瓣球药隔重楼内轮花被片宽度在3.5~6.5 mm,另两种重楼宽度皆<1 mm,因此内轮花被片的形态是一个稳定的性状。除此之外,3 种重楼在不同花期时的雄蕊、外轮花被片均存在稳定差异,并且其物候期、化学成分和ITS序列都能互相区分,并且具有各自的地理分布范围,种间界限清晰。这证明了李恒系统对球药隔重楼种下分类具有一定的合理性,但仅仅将单一性状作为各变种划分的证据还不够充分与客观[17],应当通过多种分类学指标进行综合评价,以提供更有力的支撑。
3.2 球药隔重楼种下新类群
大萼球药隔重楼与球药隔黑斑种群是近年来被发现的新类群,处于球药隔重楼地理分布的边缘地带。大萼球药隔重楼具有宽大的外轮花被片与深裂的双肾型药凸,特征鲜明,且在同样的环境下,大萼球药隔重楼物候期与其他球药隔重楼区分明显。除此之外,在2种系统发育树中大萼球药隔重楼与球药隔重楼原变种相聚,说明其与原变种亲缘关系最近,推测可能是由原变种所演化形成。
球药隔黑斑种群内轮花被片短小的特征更接近于短瓣球药隔重楼,但其外轮花被片先端的斑块、扁平状药凸、偏小的花基数均与短瓣球药隔重楼明显区分,并且在NJ 与ML 发育树中均独立聚为一支,推测可能是一新变种,但还需要进一步对其子代是否会产生性状分离进行验证。
球药隔2 种新类群的性状特征在以往的文献资料中并无记载,提示球药隔种群正在经历复杂的变异与演化,充分体现了球药隔重楼种下变异的多样性。
3.3 具柄重楼分类学地位
具柄重楼在叶片形态上与球药隔重楼较为相似,因此在药材生产上常常被误作球药隔重楼使用[18],Flora of China中也将其作为球药隔重楼的一个变种。本研究发现,具柄重楼尖头状的药凸形态与金线重楼相似;花期与倒苗期接近于金线重楼;在甾体皂苷的聚类谱系图与系统发育树中均与金线重楼聚为一类,并且在2种系统发育树中均与球药隔重楼的分支距离较远。除此之外,我们在贵州纳雍、四川宝兴所发现的过渡类型,表现出了两者共有的形态特征。这些证据都表明了具柄重楼与金线重楼具有更近的亲缘关系。已有研究表明,金线重楼与具柄重楼在叶绿体基因psbAtrnH、trnL-trnF序列所构建的系统发育树中近缘[19],本研究结果与之一致。故根据李恒系统将具柄重楼处理为金线重楼变种更为合理。
3.4 球药隔重楼地理分布与演化规律
从地理分布图得知,以我国北方的秦岭与南方的南岭为界限,由西部的四川、云南一直延伸到东部的台湾,都是球药隔重楼的分布范围。《重楼属植物》[1]认为,球药隔重楼主要分布在中国华中地区,并且在华东和台湾西部是一个分布的间断区,原因是第四纪冰川作用的影响,而在福建武夷山和三明市发现的球药隔黑斑种群,填补了上述间断的空白。其中短瓣球药隔重楼的分布范围最广,宽瓣球药隔重楼与球药隔原变种分布范围相对较窄,球药隔黑斑种群发现4 处分布区域,大萼球药隔重楼仅发现1处分布区域。
李恒[1]认为,在同等级的范围内,迁徙距离最远往往生存历史较长,因此短瓣球药隔重楼应该是球药隔重楼中最为原始的种群,其次是宽瓣球药隔重楼与原变种,并且随着种群的迁移与扩散,在分布区边缘产生了新的年轻种群,即大萼球药隔重楼与球药隔黑斑种群。根据形态观察发现,球药隔黑斑种群在球药隔重楼中花基数最小,普遍为3~4,而李恒[20]系统认为重楼花基数的演化是从大到小进行的,再次证明了这是一个较为进化的种群。
甾体皂苷类物质为重楼属植物主要的药效物质基础[21-22],并且在自然界植物中的分布存在一定的特征性,这使得该类化学物质具有一定的植物化学分类意义[23]。周荣汉等[24]认为,植物化学的性状也分为原始和进化,植物次生代谢产物的氧化水平在一定程度上能反映分子的进化趋势与水平,处于一级氧化水平的薯蓣皂苷元比二级氧化水平的偏诺皂苷元原始。化学成分显示,仅短瓣球药隔重楼与宽瓣球药隔重楼含有较多薯蓣皂苷类成分,在一定程度上证明了二者的原始性。
根据上述信息,可以初步推断球药隔重楼种内的演化规律为:短瓣球药隔重楼→宽瓣球药隔重楼→球药隔重楼原变种→大萼球药隔重楼、球药隔黑斑种群。但要进一步了解其进化过程还需要结合更多分子水平的研究。
致谢感谢四川省草原科学研究院赵文吉助理研究员拍摄并提供图片3H。
