发酵对米粉生产的降镐作用及其微生物学机理
2023-07-04徐文华张文钊侯海军魏文学盛荣
徐文华 ,张文钊 ,侯海军 ,魏文学 ,盛荣
(1. 中国科学院亚热带农业生态研究所,亚热带农业生态过程重点实验室,湖南 长沙 410125;2. 中国科学院桃源农业生态试验站,湖南 常德 415700;3. 中国科学院大学,北京 100049)
水稻是我国的主要粮食作物之一,据统计,近年来稻谷产量约占全国粮食总产量的30%,水稻的安全生产在我国粮食安全中具有举足轻重的地位。镉是一种重金属元素,在土壤中含量较低,主要以水溶态、吸附态、络合态和矿物态等形态存在[1]。由于采矿炼矿和工厂排污等问题导致部分稻田镉含量超标而生产出“镉大米”。近些年,大米镉含量超标事件屡见不鲜,给水稻的安全生产带来新的挑战。根据环境保护部和国土资源部2014 年发布的《全国土壤污染状况调查公报》显示,全国土壤总超标率为16.1%,其中镉污染超标点位最多,达到7%。在一些污染地区的大米样品调查中,约56%~87%的大米样品超过中国食品安全限值0.2 mg/kg(《食品安全国家标准 食品中污染物限量(GB2762-2017)》),其平均镉含量达到0.33~0.69 mg/kg[2]。镉是人体的非必需元素,膳食摄入是非吸烟人群接触镉的主要来源,镉进入人体后会选择性积累在各器官中,生物半衰期长达几十年之久,其毒性表现为引发人体多种综合症,包括贫血、高血压、心力衰竭、肝肺损伤、肾功能衰竭等,被列为第一类致癌物质[3]。因此,稻米镉超标已成为影响国家粮食安全和人体生命健康的重要隐患,如何实现稻谷的安全生产和镉超标稻米的安全利用备受社会和学者关注。
目前,针对“镉大米”问题主要有三个解决方向:一是从源头入手,即采取物理[4-6]、化学、生物修复等方法降低土壤中的镉含量。生物修复又包括动物修复、植物修复、微生物修复[7];二是通过编辑相关基因或蛋白调控水稻中镉的转运和积累来选育新型水稻品种[8-10];三是对已收获的镉含量超标稻米进行再加工使镉含量达标[11]。其中土壤修复以及水稻选育都需要较长的时间进程,而对“镉大米”进行加工降镉不仅能使部分镉超标大米实现安全利用,减少浪费,而且周期短,见效快,具有重要的现实意义。关于稻米加工降镉的方法主要包括物理法、化学法以及生物法。物理法是指利用物理过程将稻米镉含量降低的方法,包括加水浸泡[12]和砻谷加工[13]等方法,操作简单且对大米品质影响较小,但降镉效果一般难以使镉含量较高的稻米镉含量达标。化学法多采用酸碱浸泡使镉元素从稻米中迁出,降镉率较高且耗时较短,但若应用于实际生产则需消耗大量的酸碱,废酸废碱处理不当可能会造成环境污染,加工成本也相对较高,且是否存在因使用化学试剂导致试剂残留进而引发食品安全问题还有待进一步研究[14-15]。生物法主要指微生物发酵法,稻米发酵是食品生产过程中较常见的技术,广泛应用在米粉、米酒、米饮料等生产制作过程。研究表明,某些种类的微生物可以通过将金属离子结合到其细胞表面或将其转运储存在其细胞内从而达到去除发酵体系中重金属的效果[16],如芽孢杆菌[17]、发酵乳杆菌、植物乳杆菌等[18-20]。研究人员通过对几种传统发酵微生物菌种的初步探索发现,利用乳酸菌或罗伊氏乳杆菌+发酵乳杆菌+植物乳杆菌混菌发酵可有效降低大米和果蔬汁中的镉含量[21-24]。此外,微生物发酵还可能通过降解大米中的蛋白质,使结合态镉游离出来[25]。而且,利用微生物发酵不仅能有效消减稻米中的镉,还能提升大米的食用口感[26]。因此,利用微生物发酵降镉有望成为“镉大米”安全利用的重要手段。然而,目前关于大米发酵降镉的主要功能微生物类群及其关键作用机制还缺乏清晰的认知,严重制约我们对降镉微生物资源的挖掘利用和稻米高效、安全降镉技术的研发。
鲜湿米粉口感好,食用方便,作为主食或早餐深受我国广大消费者的喜爱。前期调查研究发现,常德市一米粉厂虽然使用了镉轻度超标大米为原料,但生产出的米粉镉含量远低于国家标准,其中大米的发酵过程可能是降镉的关键步骤。本项研究基于常德米粉加工过程的降镉现象展开,探究稻米自然发酵过程中微生物的组成结构,揭示稻米镉含量降低过程中微生物的动态变化,通过传统的微生物分离方法获得优势菌株并研究其降镉功能及作用机制,为大米发酵液中高效降镉菌株的开发利用提供参考。该项研究将丰富镉超标大米的降镉微生物资源和降镉技术,为镉超标大米安全利用提供科学依据。
1 材料与方法
1.1 试验材料
供试稻米:米粉发酵生产过程微生物种群演变及降镉作用研究采用早籼米(镉含量约为0.49 mg/kg),原始米样、成品米粉、经1、3、4 d 自然发酵的米样均采集于常德市一米粉厂;为进一步探讨分离出的发酵功能微生物菌株的降镉能力,实验室模拟发酵试验采用镉重度超标的晚籼米(镉含量约为1.05 mg/kg),取自镉污染试验基地。
发酵液:2020 年12 月,采集于米粉厂中经1、2、3、4 d 发酵的大米发酵原液(分别记作D1、D2、D3 和D4),每个发酵时间点的发酵液分别取3次重复,米粉厂原料为早籼米。
LB 琼脂培养基每升含量:蛋白胨10.0 g、酵母膏粉5.0 g、氯化钠5.0 g、葡萄糖1.0 g、琼脂15.0 g,最终pH 7.0 ± 0.2。
MRS 琼脂培养基每升含量:蛋白胨10.0 g、牛肉膏粉5.0 g、酵母膏粉4.0 g、葡萄糖20.0 g、吐温80 1.0 mL、碳酸氢二钾2.0 g、乙酸钠5.0 g、柠檬酸三铵2.0 g、硫酸镁(MgSO4·7H2O)0.2 g、硫酸锰(MnSO4·4H2O)0.05 g、琼脂15.0 g、最终pH 6.2 ± 0.2。
1.2 试验方法
1.2.1 样品中镉含量测定 采集于米粉厂的发酵过程米样、米粉产品以及模拟发酵试验的米样的处理参考GB5009.15-2014《食品安全国家标准 食品中镉的测定》中的干试样制备。
米样中镉含量的测定:参考GB5009.15-2014《食品安全国家标准 食品中镉的测定》中的湿式消解法,使用电感耦合等离子体质谱联用仪ICP-MS7900(美国安捷伦科技有限公司)测定消化液中的镉含量。
试样中镉含量的计算公式为:
式中:CCd为样品中镉含量(mg/kg),Cs为样品消化液中镉含量测定值(μg/L),C0为试剂空白溶液中镉含量的测定值(μg/L),V为样品消化液定容总体积(mL),m为样品质量(g)。
1.2.2 发酵液中微生物种群组成分析 取3 mL 发酵液于离心管中,离心后弃上清液浓缩至1 mL。采用FastDNA™ Spin Kit for Soil DNA 提 取 试 剂 盒(MP,美国)提取发酵液DNA。使用1%琼脂糖凝胶电泳和微量分光光度计检验DNA 质量,检验合格后用于发酵液微生物种群组成分析。采用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)以 及806R(5′-GGACTACHVGGGTWTCTAAT-3′)对16S rRNA 基因V3-V4 可变区进行PCR 扩增,反应体系(50 μL)为:10 μmol/L 引 物 各2 μL,2×PCR mix(Takara)25 μL,DNA 模板(20 ng/μL)2 μL,无菌水19 μL。扩增程序为:98 ℃,2 min;98 ℃ 10 s,60 ℃ 30 s,72 ℃ 45 s,30 个循环;72 ℃,10 min。使用AxyPrep DNA 凝胶回收试剂盒(AXYGEN 公司)切胶回收PCR 产物,Tris-HCl 洗脱,2%琼脂糖电泳检测。参照电泳初步定量结果,将PCR 产物用QuantiFluor™-ST 蓝色荧光定量系统(Promega公司)进行检测定量,之后按照每个样本的测序量要求,进行相应比例的混合。构建Miseq 文库,送至Illumina Miseq PE300 平台(上海美吉生物医药科技有限公司)进行测序分析。
1.2.3 发酵液中微生物的分离鉴定 取第3 d 的发酵液(D3),进行梯度稀释,分别吸取1×10-4和1×10-5稀释度样品100 μL 涂布于MRS 和LB 固体培养基,每个稀释度设3 个重复。于35 ℃生化培养箱倒置培养2~5 d。观察并挑选外观形态特征不同的单菌落在相应的培养基上划线分离,经过2~3 次划线纯化后得到单一菌株,对分离出的单菌落进行菌体形态观察,并对形态特征有明显差异的菌株进行16S rRNA 基因序列分析。
16S rRNA 基因PCR 扩增和测序引物采用27F(5′-AGAGTTTGATCMTGGCTCAG-3′) 或8F(5′-AGAGTTTGATCCTGGCTCAG-3′) 和1492R(5′-TACGGCTACCTTGTTACGACTT-3′),25 μL 菌落PCR 反应体系:2×Easy Taq Master Mix 12.5 μL(含有Taq DNA 聚合酶、dNTPs、缓冲液、MgCl2、溴酚蓝),10 μmol/L 引物各1 μL,ddH2O 10.5 μL,用无菌牙签挑取单菌落到反应体系中。PCR 扩增程序为:95 ℃预变性5 min;95 ℃ 45 s,50 ℃ 45 s,72 ℃90 s,35 个循环;72 ℃ 10 min。PCR 产物使用1%琼脂糖凝胶电泳检测。检验合格后将PCR 产物送至上海生工生物工程技术服务有限公司测序。通过NCBI-BLAST(http: //www.ncbi.nlm.nih.gov),将 获得的16S rRNA 基因序列与GenBank 数据库中的序列进行比对获取目标菌株的分类学信息。
1.2.4 菌株降镉效率试验 制备接种菌液:从固体培养基上挑取单菌落接种至装有20 mL 相应液体培养基的50 mL 锥形瓶中,设置不接种对照,置于35 ℃恒温培养箱震荡培养6~7 h 至液体培养基浑浊,离心后去上清液,将菌体沉淀用无菌水洗涤两次,最后用无菌水重悬菌体。利用全自动生长曲线分析仪测定调整菌液OD600值为0.8,即为接种菌液。
发酵体系:在洁净工作台中,将50 g 晚籼米加入250 mL 锥形瓶中,再加入75 mL 灭菌的去离子水,按1%接种量接入菌液(即0.75 mL),对照组加入等量灭菌的去离子水,封瓶膜封口。体系中料液比(g/mL)为1.0:1.5。在整米发酵与大米粉发酵对比试验中,考虑到大米粉的吸水性较高,发酵体系料液比(米水比例(g/mL))调整为1:3,具体如下:将50 g 粉碎过筛的晚籼米粉或整米分别装入灭菌的250 mL 锥形瓶,加入150 mL 灭菌的去离子水,按1%接种量接入菌液(即1.5 mL)。对照组加入等量灭菌的去离子水。
1)单菌株降镉效率试验。初步排重后选取不同菌种进行单菌株降镉效率的试验,同时设置不接菌的对照。每个菌株的发酵体系设置三次重复,置于35 ℃生化培养箱培养48 h。采集发酵后的米样测定其镉含量。
2)高效菌株组合降镉效率试验。选取单菌株降镉试验中降镉效率较高的菌株,进行两株菌、三株菌、四株菌的组合发酵试验。同时设置不接菌的对照,不同组合的发酵体系中总的菌液接种量为1%,多菌株接种菌液中各菌株所占的比例相等。每个组合的发酵体系设置三次重复。置于35 ℃生化培养箱培养48 h。处理发酵后的米样后测定其镉含量。
发酵后米样的处理:发酵结束后倾倒发酵液,在锥形瓶中加入300 mL 左右的去离子水将米样洗涤四次。洗涤后的米样置于培养皿上,在70 ℃的电热鼓风干燥箱中烘干。烘干后的米样一半保存于自封袋中,另一半用粉碎机粉碎,过35 目筛后保存于自封袋中。粉碎的米样用于后续镉含量的测定(参考GB5009.15-2014《食品安全国家标准 食品中镉的测定》中的干试样制备)。
降镉效率的计算公式为:
式中:W表示降镉效率(%),W0是原料大米中的镉含量,W1是发酵后大米的镉含量。
1.2.5 探究发酵降镉的微生物机理 选择L4 和L4+L8 两种菌液分别接种至整米发酵体系和大米粉发酵体系中,控制总接种量为1%,探究大米形态对降镉效率的影响。
收集发酵结束后的发酵液,对发酵液中pH 值与乳酸含量进行测定。pH 值的测定:使用pH 计对发酵液的pH 值进行测定。乳酸含量的测定:发酵液通过一次性细菌过滤器(0.22 μm)过滤后注入上样瓶中,使用气相色谱仪7890A(美国安捷伦科技有限公司)进行检测。探究发酵液pH 值、乳酸含量与降镉效率的关系。
1.2.6 数据统计与分析 采用Excel 软件对数据进行统计分析。采用Excel 和Origin 软件进行绘图。显著性差异采用SAS 9.4 软件进行分析,采用单因素(one-way ANOVA)和Duncan 法进行方差分析和多重比较。测序获得的序列用MEGA7.0 软件进行系统发育分析。所有分离菌株的16S rRNA 基因序列均已提交GenBank 数据库,并获得了相应的序列登录号。
2 结果与分析
2.1 发酵过程米样及米粉产品镐含量变化
在常德米粉加工过程中,稻米会在发酵池中加水浸泡静置4~5 d,且发酵液(前一批发酵产物)的接种是米粉制作不可或缺的步骤。通过采集测试分析米粉厂中不同发酵时间点的米样以及成品米粉中的镉含量发现(图1),随着发酵时间的延长,米样中的镉含量逐渐降低。其中从第1 d 到第3 d,米样中的镉含量降低了约37%;到第4 d,米样中镉含量降低了近80%,降幅可达0.25 mg/kg,稻米中残留的镉含量仅为0.06 mg/kg,远低于国家标准0.20 mg/kg。将发酵过的稻米加工成米粉的过程中,镉含量也有所下降,进一步降至0.04 mg/kg。可见,发酵过程对稻米中镉含量的降低发挥了巨大作用,因此推测,发酵液中的微生物在稻米降镉过程中发挥了至关重要的作用。
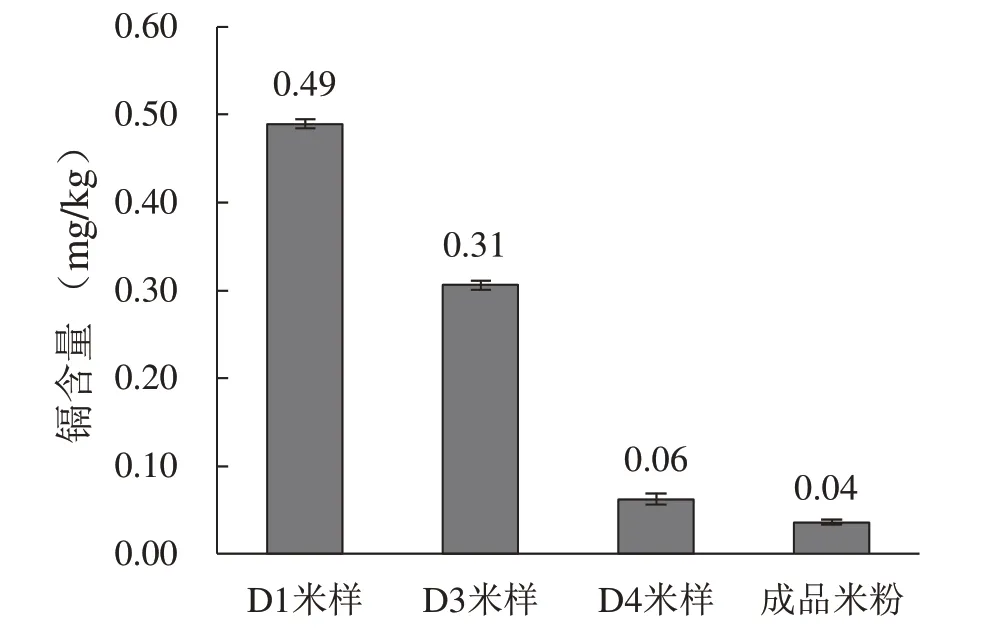
图1 米粉厂不同发酵时间和成品样品中的镐含量Fig. 1 Cadmium content in the rice noodles and rice samples during the fermentation process
2.2 发酵过程细菌种群结构演变及菌株分离与分子鉴定
通过分析不同发酵时间大米发酵液中细菌的种群组成结构特征发现(图2),稻米发酵前三天(D1,D2,D3)的发酵液细菌属水平组成结构比较相似,乳杆菌属的物种总丰度占比几乎达100%,为大米发酵液中的绝对优势细菌属。但第4 d(D4)的发酵液的细菌组成结构发生了显著变化,主要表现在乳杆菌属的相对丰度急剧下降至9.08%,而乳球菌(Lactococcus)、丛毛单胞菌(Comamonas)迅速增长为优势菌属,其相对丰度分别占总丰度的33.04%和31.08%。

图2 不同时间发酵液中种和属分类水平下细菌群落结构特征Fig. 2 Characteristics of bacterial community structure at the level of species and genus in diff erent samples
进一步从种水平上分析发现,前三天发酵液中的优势菌种主要是一些未分类的乳杆菌(unclassif iedLactobacillus)、黏膜乳杆菌(Lactobacillus mucosaeLM1)、解 淀 粉 乳 杆 菌(Lactobacillusamyloνorus,Lactobacillusamylolyticus)、唾液乳杆菌(Lactobacillus saliνarius)等。第1 d(D1)发酵液中的优势菌种为未分类的乳杆菌和黏膜乳杆菌,其占比约为90%;第2 d(D2)发酵液中未分类的乳杆菌和黏膜乳杆菌占比分别下降20%和10%左右,解淀粉乳杆菌所占比例较第1 d 提高了20%左右;第3 d(D3)发酵液中未分类的乳杆菌和黏膜乳杆菌的占比较第2 d 有所提高但仍低于第1 d,而解淀粉乳杆菌含量相对减少。然而在第4 d(D4)的发酵液中乳杆菌属的未分类的乳杆菌、解淀粉乳杆菌和黏膜乳杆菌等急剧减少,取而代之出现大量乳球菌(Comamonastestosteroni)和丛毛单胞菌(Comamonas kerstersii)以及稳杆菌属(Empedobacter)、库尔特氏杆菌属(Kurthia)、莫氏假单胞菌(Pseudomonas mosselii)等致病菌或腐败菌,这些菌成为优势类群。此结果说明,在米粉生产发酵体系中,前三天的发酵状况趋于稳定,菌种组成稳定,乳杆菌为主要的发酵菌种,但在第4 d 时致病菌或腐败菌大量出现,如果继续发酵将可能影响发酵效果和米粉品质。结合2.1 中的结果,推测前三天发酵液中微生物即乳杆菌,在降镉效应中发挥了主要作用。
2.3 大米发酵液中微生物的分离鉴定
为进一步明确发酵液中微生物是否具有降镉功能,选取第3 d 的发酵液进行了微生物菌种的分离鉴定。结果显示,从发酵液(D3)中共分离出58 株可培养细菌,经16S rRNA 分子鉴定,主要分布在4 个不同的细菌属,9 个细菌种。其中乳杆菌属Lactobacillus(63.79%)所占比例最高,与上文推测一致,其次是片球菌属Pediococcus(25.86%)和微球菌属Micrococcus(8.62%),仅分离出一株芽 孢 杆 菌 属Bacillus菌 株:L8(Bacillus subtilis)。初步排重以及排除一个不易培养的菌种粘膜乳杆菌(L. mucosae)后,选取以下8 株细菌进行后续试验:发酵乳杆菌(L. fermentum)L4、德氏乳杆菌(L. delbrueckiisubsp.)M30、乳酸片球菌(P.acidilactici)P1、藤黄微球菌(M. luteus)L19、云南微球菌(M. yunnanensis)L20、芦荟微球菌(M.aloeνerae)L24、微球菌(Micrococcussp.)L25、枯草芽孢杆菌(Bacillus subtilis)L8(表1)。所选菌株的系统发育关系如图3 所示。

表1 代表菌株的基本信息Table 1 Related information of the representative isolated strains
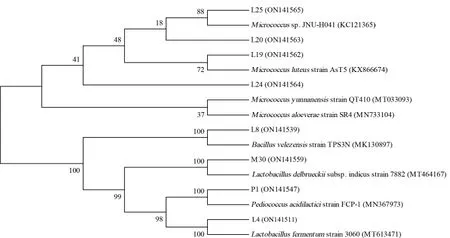
图3 基于代表菌株的16S rRNA 序列构建的NJ 系统发育进化树Fig. 3 Neighbor-joining phylogenetic (original) tree based on 16S rRNA gene sequence of representative isolated strains
2.4 分离菌株的降镐效果
2.4.1 单菌降镉效果 通过分离菌株对重度镉超标稻米的降镉试验发现,与不接种对照相比,发酵乳杆菌、德氏乳杆菌、乳酸片球菌、芽孢杆菌均表现出不同程度的降镉能力(图4)。其中,乳酸片球菌P1 菌株的降镉能力最高,发酵48 h 的降镉量达到0.55 mg/kg。发酵乳杆菌L4 的降镉能力与P1 相近,为0.54 mg/kg,其次是芽孢杆菌L8 和德氏乳杆菌M30,其降镉量分别为0.34 和0.33 mg/kg。相比之下,其他菌株的48 h 降镉能力均在0.15 mg/kg 以下,且与对照组相比无显著差异。由此可见,来源于发酵液中的乳酸菌确实有较强的降镉作用,但因本试验使用的稻米为重度超标的晚籼米(1.05 mg/kg),导致其降镉效率分别仅为51.75%、51.08%、31.83%和30.90%,其超高的镉含量和致密的种质结构可能是影响降镉效率的重要因素。
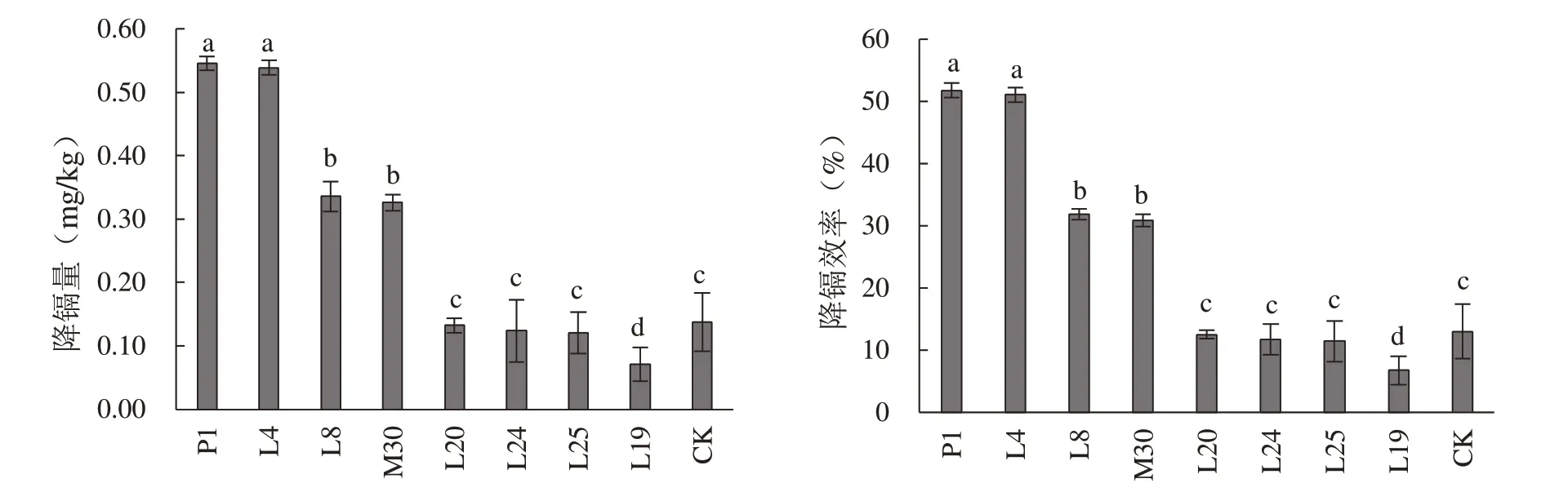
图4 8 株代表菌株的降镐量和降镐效率Fig. 4 The cadmium removal capacity and effi ciency of the representative strains
2.4.2 复合菌株降镉效果 由于以往研究显示,多种功能互补菌群进行组合有助于提升发酵品质,如德氏乳杆菌和枯草芽孢杆菌具有多种互补益生性能,被广泛应用在多种发酵工业和食品行业[27-28]。故本研究选取乳酸片球菌P1、发酵乳杆菌L4、德氏乳杆菌M30、芽孢杆菌L8 四株高效降镉菌株进行组合发酵试验。
结果显示(图5),与单菌株发酵效果相比,在总接种量一致的条件下,双菌组合发酵的降镉效率并未显著提升。其中L4+L8 的降镉效率显著高于其他双菌组合,48 h 内可去除晚籼米中0.46 mg/kg的镉,降镉效率达到43.59%,其次为L4+P1 组合,其降镉量和降镉效率分别为0.42 mg/kg 和39.45%,但其降镉能力均低于L4 或P1 单菌株。L8+M30 的降镉能力最差,48 h 镉去除量仅为0.19 mg/kg,降镉效率为17.82%。
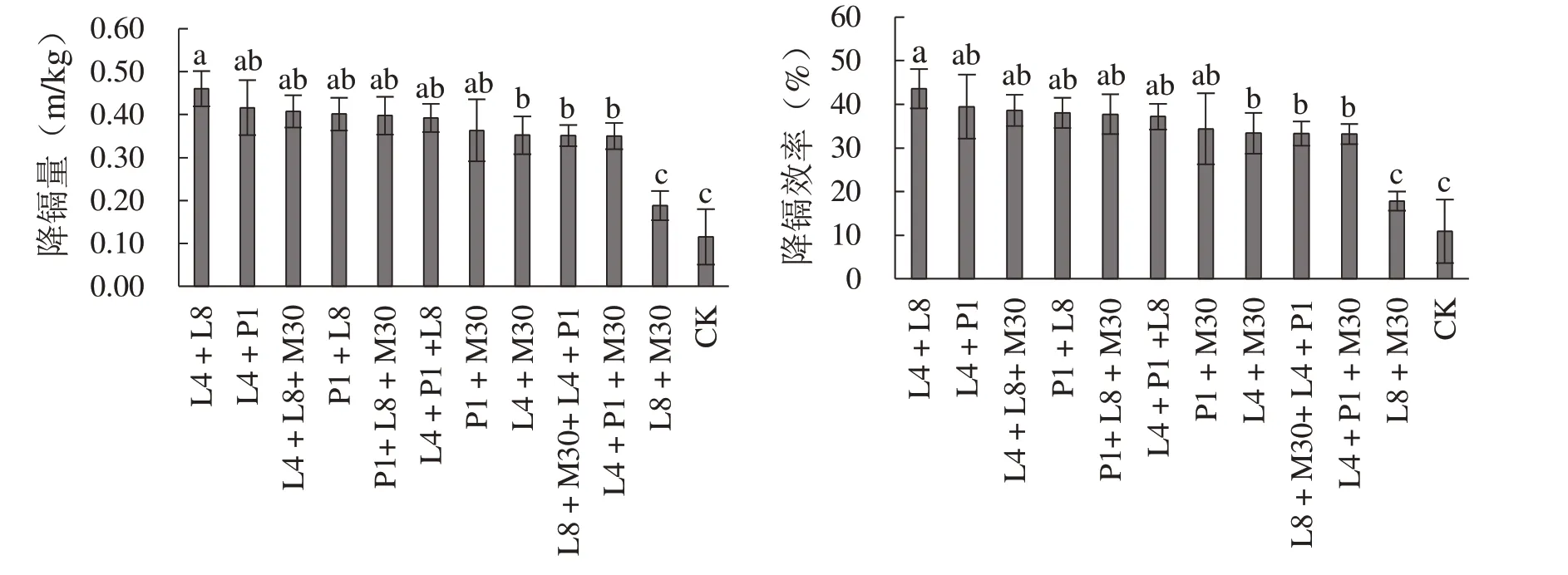
图5 不同菌种组合的降镐量和降镐效率Fig. 5 The cadmium removal capacity and effi ciency of the diff erent strains’ combinations
在双菌组合的基础上进行三菌或四菌组合发酵,并未提升发酵体系的降镉效率,甚至有所降低(图5)。例如,与L4 和L8 双菌组合相比,L4+L8+M30 的降镉效果显著降低(P<0.05),镉去除量为0.41 mg/kg,降镉效率为38.64%。L4+P1双菌组合改为L4+P1+M30 三菌组合或四菌组合后,其降镉能力也发生显著下降(P<0.05)。但将L8+M30 组合转变为L8+M30+P1 组合后,其降镉能力从0.19 mg/kg 显著提高到0.40 mg/kg,涨幅达到1.12 倍,说明在低活性发酵体系中添加功能活性较强的菌株可有效提高发酵体系降镉功能。
其余三菌或四菌组合与双菌组合的降镉效率较为接近,降镉量均在0.35~0.40 mg/kg 之间,无显著差异(P>0.05)。由此可见,在接种总量一定的发酵体系中,降镉功能主要以高活性菌株为主导,当以相对低活性菌株替换部分高活性菌株生态位时,降镉能力也会有所损失。因此,在后续混菌组合发酵试验过程中应考虑菌种比例合理性,在保障高活性菌种接种量的基础上添加无拮抗作用的功能辅助菌。
2.4.3 米粉发酵过程降镉机制探索 由于已有的研究表明,微生物发酵过程中有机酸和蛋白酶的产生可能在蛋白结合态金属消解过程中发挥了重要作用。因此,本研究选取对照组与试验组(L4 单菌株、L4+L8 组合),探究其发酵结束时发酵液的pH 值与乳酸含量差异,结果表明(表2),发酵结束时试验组与对照组均为酸性环境,但与对照组相比,L4 和L4+L8 组合处理发酵液pH 值分别显著降低了0.60和0.53 个单位(P<0.05)。试验组L4 和L4+L8 组合发酵液中的乳酸含量也显著增加,分别达到对照组的1.53 倍和1.68 倍(P<0.05),这与其发酵液中低pH 值相符。由此我们可以推测,大米发酵液中微生物产生的有机酸等物质促进了镉结合蛋白质的溶出,从而降低稻米中的镉含量。

表2 不同处理发酵结束时酵液pH 值和乳酸含量及降镐效率和降镐量Table 2 The pH and lactic acid content of the rice fermentation broth of the diff erent treatments andthe cadmium removal capacity and effi ciency
在本研究的稻米模拟发酵实验中,微生物菌种添加并未将重度镉污染的晚籼米(镉含量约为1.05 mg/kg)的镉含量降至国家标准以下,推测原因可能是其超高的镉含量和致密的种质结构影响了降镉效率。为进一步证实这一推测,本研究采用大米粉(粉碎的镉超标大米)为试验材料,对比分析了对照组与试验组(L4 单菌株、L4+L8 组合)的降镉能力、发酵液pH 和乳酸含量。结果发现,与整米处理相比,采用大米粉为原料可大幅提高降镉效率,其中L4 和L4+L8 处理的降镉量分别提高到0.64 和0.56 mg/kg,降镉效率分别达到67.14%和59.10%,显著高于稻米处理(P<0.05)。与此同时,其发酵液的pH 下降幅度更大,分别比对照下降了1.52 和1.54个单位(P<0.05),而乳酸含量分别达到了对照组的6.55 倍和7.11 倍。由此可见,在镉超标大米加工利用过程中,粉碎处理可进一步促进稻米中镉的溶出。
3 讨论
3.1 大米自然发酵液中的降镐微生物及其降镐能力
本文通过对大米发酵液样本中的细菌进行高通量测序以及微生物分离鉴定,结果表明,乳杆菌为发酵液中的优势细菌种。这与前期稻米等粮食作物发酵过程微生物菌群组成的研究发现基本一致[29-30],认为植物乳杆菌为大米发酵液的主要细菌物种,其中的大多数菌种具有消化淀粉、糖、蛋白质或脂质的能力[29]。由于镉元素在大米的各个部位都有分布,因此这些菌种可能也具有一定的降镉能力。在本研究的降镉试验中,乳酸片球菌P1、发酵乳杆菌L4、德氏乳杆菌M30、芽孢杆菌L8 均表现出不同程度的降镉效果。乳酸菌具有较好的降镉效果,这与前期初步研究结果一致[21-24]。而在其他领域的降镉过程中,芽孢杆菌也是一种常用的降镉微生物。例如,在重金属污染的废水中,芽孢杆菌与生物炭共同作用对镉的去除有良好的效果[17,31];亚马逊本地的芽孢杆菌可降低可可植株中的镉积累[32],同理,在其他地区植株的根际也可能存在可以降低植株镉积累的菌株。在本研究中,也分离到有降镉作用的芽孢杆菌。
通过分析各菌株的降镉能力发现,单菌株在48 h内的降镉量高达0.55 mg/kg,但因本试验使用的稻米为重度超标的晚籼米(1.05 mg/kg),导致其降镉效率在60%以下,未能将其镉含量降到国家安全标准以下。尽管如此,由于我国镉超标大米中镉含量范围普遍在0.33~0.69 mg/kg,大部分属轻度或中度镉超标,因此该系列菌株在我国“镉大米”安全利用中仍具有巨大的应用前景。鉴于以往研究表明,多菌株混合发酵有助于提升发酵效率[33],因此本研究进一步分析了多菌株发酵体系对稻米镉含量的影响。但结果发现,多菌种等比例组合发酵似乎没有显示出菌种多样性的优势,其降镉效率甚至低于单菌或双菌发酵,这可能与功能菌接种比例和总量有关。本研究所有处理接种总量均为1%,当发酵体系中部分高活性的菌株(如L4 和P1)被替换为相对低活性的菌株(如L8 和M30)时,其发酵体系的降镉能力明显下降。在同一发酵体系中,不同菌种对资源空间的竞争和拮抗作用可能是影响降镉效率的重要原因[34]。李芸[26]的研究也发现混菌培养时两种菌均未达到单菌培养时的最大生物量。在混合菌发酵体系中菌种间互作机理及其对发酵体系产生的影响机制还有待进一步探究。基于上述结果,在后续发酵试验中,可考虑在优先保障高效菌株接种量的基础上添加功能互补型菌株,以进一步强化发酵体系的降镉功能,提高组合发酵的降镉效率从而应用于实际生产。
3.2 米粉发酵过程降镐机制探索
大量研究表明,部分镉元素在稻米中的存在形式是蛋白结合态[13,33,35],发酵乳杆菌、德氏乳杆菌、乳酸片球菌等均会分泌大量乳酸[28,36-37],乳酸可以通过其ɑ-羟基结构的羧基与肽链上的基团形成氢键促进蛋白质的溶出[21-22,38],从而表现出降镉作用。而枯草芽孢杆菌往往具有产蛋白酶性能[27,39],其分泌的蛋白酶直接作用于稻米蛋白,使得枯草芽孢杆菌也表现出降镉作用。本研究通过对比分析对照组与试验组(L4 单菌株、L4+L8 组合)发酵液的pH值与乳酸含量差异发现,L4 和L4+L8 组合处理均显著降低了发酵液pH 值,同时显著提高了其乳酸含量,进一步证实了大米发酵液中微生物产生的有机酸和蛋白酶等使发酵体系处于较低pH,促进了镉结合蛋白质的溶出,使米粉生产发酵过程表现出降镉效应。这为高效降镉菌株的开发提供了一种思路,可筛选一些性能优良的工业微生物如高产酸菌、高产蛋白酶菌等运用到食品降镉行业,从而进一步丰富降镉功能微生物资源。
此外,稻米的质地形态可能也是影响其降镉效率的重要因素,本研究通过对比分析整米和大米粉(经过粉碎的大米)在微生物发酵体系中的降镉效应发现,采用粉碎后的稻米为原料可将发酵体系降镉效率提升50%以上。这可能是因为粉碎后的稻米为微生物提供了更多的营养物质支持微生物生长,同时也增加了微生物与镉结合态蛋白的作用面积,促进了镉元素的游离或溶出。研究结果中大米粉发酵体系的pH 下降幅度更大,乳酸含量也显著增加的数据可进一步证实大米粉发酵体系中降镉微生物活性更高。由此可见,在镉超标大米加工利用过程中,粉碎可进一步提高功能微生物的发酵活性,加速稻米中镉的溶出。
4 结论
本研究以鲜湿米粉加工过程中的发酵液为试验材料,通过对发酵液微生物高通量测序、功能微生物的分离培养及降镉能力试验等,研究了米粉生产过程中的降镉作用及其微生物学机理,结论如下:
1)鲜湿米粉加工工艺中的发酵过程可有效降低稻米镉含量,而发酵液中的微生物起关键作用。第1~3 d 的发酵液中细菌群落结构相似,均以乳杆菌属为优势菌属,第4 d 的发酵液中出现一些致病菌或腐生菌,因此实际生产中稻米最佳发酵时间以三天为宜。
2)通过对稻米发酵液中微生物的分离培养和降镉能力试验,发现发酵液中分离的发酵乳杆菌、德氏乳杆菌、乳酸片球菌和枯草芽孢杆菌等菌株均表现出较好的降镉效果,48 h 内的最高降镉量可达0.55 mg/kg,在我国“镉大米”安全利用中有巨大的潜力。其中发酵乳杆菌和乳酸片球菌的降镉效率显著高于德氏乳杆菌和枯草芽孢杆菌。菌种组合试验表明,除L4+L8(发酵乳杆菌+芽孢杆菌)组合外,其他组合均未表现出比单菌更高的降镉效率,在后续发酵试验中可优化菌种配比,保障高效功能菌株接种量基础上添加适量功能辅助菌,以进一步强化发酵体系的降镉功能。
3)菌种添加使发酵液pH 显著降低、乳酸含量升高,进而促进镉结合态蛋白分解,可能是米粉加工过程中稻米镉含量降低的主要作用机制。稻米粉碎可提高功能微生物活性并增加作用面积,大幅提升发酵体系降镉效率。该项研究对镉超标大米安全利用具有重要的实践意义,并提供了有效的降镉微生物菌种资源。
