养分添加与干扰对荒漠草原群落组成及构建的影响
2023-06-28乔静娟左小安王国林王景圆王泽宙
乔静娟, 左小安, 岳 平, 王国林, 王景圆, 王泽宙
(1.中国科学院西北生态环境资源研究院乌拉特荒漠草原研究站,甘肃 兰州 730000;2.中国科学院大学,北京100049;3.甘肃省寒区旱区逆境生理与生态重点实验室,甘肃 兰州 730000;4.内蒙古乌拉特梭梭林蒙古野驴国家级自然保护区乌拉特后旗管理站,内蒙古 乌拉特 015500)
荒漠草原作为草原中最旱生的类型,在全球范围内分布广泛且易受到环境条件波动的影响。自工业革命以来,全球范围内的养分沉降和人类活动对荒漠草原群落物种组成、土壤理化性质和生态系统功能造成了严重影响[1-2],改变了群落内物种稳定共存及维持的构建机制[3-4],导致荒漠草原生态系统功能与服务的维持与保护面临巨大挑战。以往的研究表明,由于植物间资源利用效率的差异[5]和抗干扰能力的强弱[6],养分添加和干扰会影响荒漠草原局域群落内的物种组成(α多样性)和群落间物种组成的差异(β多样性)。由于多重养分添加会降低土壤生态位维度,增强环境过滤的定向选择作用,因此将导致物种多样性的丧失[7-8]。大量研究表明,干扰对生物多样性的影响符合驼峰模式(中度干扰理论),即在中等程度的干扰水平下,群落能维持较高的物种多样性,这是因为过低的干扰水平导致只有少数竞争力强的物种在群落中占据优势,而过高的干扰水平导致少数生长速度快、侵占能力强的物种才能生存下来[6,9]。然而,在物种丧失尚未发生时,群落的性状结构可能已经因栖息地环境破坏而改变,因此基于性状的分析预计更有助于提前了解生态系统应对环境变化的响应[3,10]。
功能性状表征植物体定植、存活、生长和死亡等的一系列属性,反映了植物对外部环境条件变化的响应和适应,并将环境与植物个体和生态系统联系起来[11-13]。经过长期的自然筛选和进化,不同功能型植物根据其功能需求,在叶片功能性状之间形成了在资源获取与分配中的权衡关系,即叶经济谱(leaf economics spectrum, LES)[14]。在LES 的一端,为“快速投资-收益”型物种,它们具有寿命短、叶氮含量高、叶磷含量高、光合能力强、呼吸速率快和比叶质量小的性状特点;而在LES 的另一端,则为“缓慢投资-收益”型物种,功能性状特点与“快速投资-收益”型物种相反[14-15]。高度反映了植物对光照资源利用的强弱,叶片厚度和叶干物质含量和植物的持水能力和抗逆性密切相关,比叶面积是叶经济谱中表征植物在不同生境下资源获取、利用以及适应特征的核心性状,研究这些性状如何响应养分添加和干扰对阐明群落功能结构变化和生态系统演替机制具有重要意义[15-16]。此外,由于生态系统功能在很大程度上由群落内的优势种决定[10,17],因此,综合考虑群落物种组成和功能性状特征,基于质量比假说(Mass Ratio Hypothesis,MRH)的群落加权性状均值(Community-weighted mean,CWM)作为表征功能多样性的关键指标,能够很好地反映群落整体如何响应与适应环境变化[3,17]。目前,尽管有一些研究聚焦于养分添加对荒漠草原植物功能性状的影响,但这些研究大多聚焦于单一的实验处理和功能性状,并不能全面地揭示荒漠草原植物群落对养分添加和干扰以及交互处理的响应和适应策略,因此,探究几种反映植物不同生态策略的功能性状以及CWM如何随着养分添加和人为干扰变化以及性状与局域土壤因子的联系,既可以使我们充分了解荒漠草原不同植物资源利用策略的特征,也能清晰地阐明环境波动造成的土壤因子的变化如何驱动当地生物多样性的变化,进一步为脆弱生态系统的保护提供理论依据。
群落内物种如何稳定共存以及多样性的维持机制一直是生态学研究的热点问题[18],研究表明,确定性过程和随机性过程的强弱受环境条件和生态系统类型等的影响[18-19]。确定性过程强调共存物种之间的生态位分化,包括环境过滤(主要由气候和土壤因子决定)和各物种间的相互作用(例如竞争、促进等),这些非生物和生物因素决定了物种的存在/缺失和相对丰度[20]。随机性过程基于中性理论,通常包括随机的出生-死亡事件、概率扩散和生态漂变,而这些变化并不是由环境决定的适应性结果,而是强调群落内共存物种组合的随机性和偶然性[21]。养分添加和干扰可能会改变荒漠草原确定和随机过程的相对重要性,从而改变群落构建的轨迹。例如,养分添加造成的土壤养分富集和酸化可能更有利于更强的资源竞争或者耐受低pH 的物种,而干扰引起的土壤环境条件的变化可能更有利于演替早期物种的定殖和繁衍,这将加强确定性过程的重要性[4]。另一方面,养分添加和干扰预计会造成群落生产力的提高,这将加强随机性过程的重要性[22]。运用零模型(Null Model)方法, 目前的大量研究基于物种的存在-缺失或者丰度数据来探索群落构建机制[23],但这些研究主要集中在其他生态系统上,在荒漠草原草本群落,养分添加和干扰引起的群落组成的变化如何影响构建机制仍然不清楚,并且很少有研究同时运用两种零模型方法来比较探究。
荒漠草原是我国北方重要的草原类型,面临着日益严重的全球气候变化和人为干扰作用下的退化威胁[1,24-25]。乌拉特荒漠草原作为内蒙古集中分布的天然草场之一,是国家生态安全战略格局中“东北森林屏障带”和“北方防沙带”的重要组成部分,因当地恶劣的自然环境条件,生态脆弱性强[24,26]。近年来,各种人为干扰叠加在全球气候变化之上,进一步加剧了当地荒漠草原生态系统的退化。目前,大部分研究主要聚焦于单独探究养分和干扰对荒漠草原植物群落物种多样性和功能性状的影响上,很少有研究同时从物种多样性、功能性状、土壤理化性质和群落构建机制的角度来全面地探究这两种因子及其交互作用对当地草本群落的影响。因此,本研究以乌拉特荒漠草原草本植物群落为研究对象,通过野外控制试验,探究养分添加和干扰处理对当地植物群落组成和群落构建机制的影响,旨在揭示荒漠草原植物群落响应与适应全球气候变化和人为干扰的机制,从而为国家生态安全和北方生态屏障的维持和保护提供理论依据和实践指导。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古巴彦淖尔市乌拉特荒漠草原研究站干扰与恢复试验场(107°E,41.4°N,海拔1650 m)。该地区地处中温带,属高原大陆性气候,气候特点为冬季寒冷干燥,夏季干燥少雨,日照强烈,蒸发量大。年平均气温约3.8 °C,平均降水量100 mm。7—8 月降水量占全年降水量的70%[27]。由于长期风蚀,土壤养分含量较低[27]。年无霜期130 d,年冻土深度2.3 m。根据中国土壤分类系统(国家土壤调查局,1998 年),将土壤类型分为棕壤和灰棕荒漠土,土壤pH 范围为7.5~9.5。研究区的建群种为沙生针茅,优势种主要为骆驼蓬和猪毛菜,此外,蒙古虫实、碱韭、蒙古韭、天门冬、鸦葱、兔唇花、冷蒿、糙隐子草、银灰旋花、砂蓝刺头和冰草也是该研究区的常见物种(表1)。

表1 荒漠草原主要优势物种生活型Tab.1 Life form of dominant species in desert steppe
1.2 试验设计
本研究样地为全球草原干扰和资源网络(Disturbance and Resources Across Global Grasslands Network,DRAGNet)(https://dragnetglobal.weebly.com)的一个试验点。为了尽可能减小环境异质性对试验结果的影响,于2019年4月在2013年围封的大样地内选取了环境条件较为均匀,地势较为平坦的代表性荒漠草地进行了试验处理。试验共包含4 个处理,分别为:对照(Control)、营养元素添加(NPKμ)、物理干扰(Dis)和养分添加+干扰(NPKμ+Dis),每个处理重复5 次,共20 个试验小区,每个小区的面积为5 m×5 m。为了尽可能避免相邻小区之间互相影响,相邻小区之间设置1 m的缓冲带。NPKμ处理同时添加大量和微量元素,大量元素采用与全球营养物研究网络(Nutrient Network, NutNet) 中“NPKμ”处理相同的养分添加方案,即N、P和K各10 g·m-2,从2019年开始在每年8月的中下旬添加,微量元素采用微肥复合肥,成分及比例为Ca: Mg: S: B: Cu:Fe: Mn: Mo: Zn = 6: 3: 12: 0.10: 1: 17: 2.50: 0.05: 1(%),只在处理的第一年即2019 年8 月的中下旬进行添加,具体添加量为100 g·m-2。采用物理干扰的方法模拟人为干扰对荒漠草原典型植物群落的影响,在每年8月中下旬进行试验处理,具体步骤为先移除所有植物地上部分并耕地15 cm 深,然后去除植物地下根系,探究干扰后植物群落物种组成和演替的过程。此外,为了探究群落如何响应与适应养分添加和干扰的相互作用,本试验同时设置NPKμ+Dis的交互处理,每年8月中下旬去除植物地上部分后,耕地和去除根系,然后进行养分添加,具体的养分添加方式和养分用量和单独的养分添加处理一致。
1.3 调查方法
在试验处理的第三年,即2021年生物量高峰期的7月下旬至8月上旬进行采样。采用样方法进行植被调查,每个试验区随机选取1 m×1 m样方,为了避免边缘效应,将样方选取在距离小区边缘至少0.5 m的位置。完成后,采用齐地面刈割法收获样方内的植物并带回试验室,在65°C 下干燥48 h 至恒定重量,测量其干重,获得每种植物的地上生物量。根据功能性状采样和测量的标准方法[28],在每个样方中对优势物种(主要为沙生针茅,骆驼蓬,猪毛菜和蒙古虫实)的叶片进行采样和测量。测定的性状指标包括:株高(Height, cm)、叶片厚度(Thickness,mm)、比叶面积(Specific Leaf Area,SLA,m2·kg-1)、叶干物质含量(Leaf Dry Matter Content,LDMC,g·kg-1)、叶片氮含量(Leaf Nitrogen Content,LNC,g·kg-1)和叶片碳含量(Leaf Carbon Content,LCC,g·kg-1)。选择每种植物的10~60个左右完全舒展且无病虫害的向阳叶,将叶片放在潮湿的滤纸之间,并迅速放入密封袋中,储存在便携式车载冰箱(内部温度<5°C)。在试验室去除叶片叶柄,将叶片在水中浸泡12 h,使叶片充分吸水并展开。取出叶片,擦干表面水分,用千分之一电子秤称量得到叶片鲜重,然后将叶片平铺在扫描仪上进行扫描,再用ImageJ(Version 1.52a)软件进行处理,得到叶片总面积。最后将叶片放入烤箱内,在65°C 烘干48 h,称量得到叶片干重。同时采集优势植株的叶片,先在105°C下烘干12 h,然后在65 °C 下烘干48 h,后将样品粉碎,用Costech 元素分析仪(Costech Ecs4010, Italy)测定各物种叶片碳氮含量。每个小区用直径3 cm 的土钻在0~10 cm 土层挖取3 个土样,然后将3 个土样混合,置于通风处晾干,用孔径2 mm的筛子筛分后,粉碎土壤,测量土壤全碳含量(TCC,g·kg-1)、全氮含量(TNC,g·kg-1)(Costech Ecs4010,意大利)、pH和土壤电导率(Electrical conductivity, EC)。使用SX823 pH-电导率仪测定pH 和电导率,土壤含水量(Soil Water Content,SWC)采用烘干法进行测量[29]。
SLA和LDMC的计算公式分别为:
群落水平的功能多样性用群落性状加权均值CWM(Community-weighted mean)表示,计算公式为:
式中:Pi表示物种i在群落内的相对生物量;traiti表示物种i的性状值;n表示群落内的物种数。
1.4 统计分析
数据的统计分析以及绘图均在RStudio(2022.02.1+461) 中完成。将4 个处理作为固定效应,5 个区组作为随机效应,使用“lme4”和“sjstats”包通过线性混合模型比较不同处理下物种多样性、土壤理化性质、功能性状和群落构建指数的差异。利用“vegan”包和“adespatial”包分别进行物种非度量多维标度(NMDS)分析和群落β多样性的计算,利用“FD”包计算6个性状的CWM,土壤和性状主成分分析利用“FactoMineR”和“factoextra”包完成。为了探究不同处理下土壤理化性质变化与CWM 的关系,采用混合线性模型进行回归,除厚度性状外,对CWM 进行ln 变换。采用Duncan 法进行多重比较。零模型采用基于物种存在-缺失数据的“NST”指数[30]和基于物种丰度数据的“Raup-Crick”指数[31]来探究不同处理下群落构建的确定性或随机性过程。
2 结果与分析
2.1 养分添加和干扰对群落物种组成和土壤理化性质的影响
NMDS 分析表明,不同处理下群落物种组成存在显著差异(Stress<0.05,图1)。干扰和交互处理下,物种组成较为单一,蒙古虫实(Corispermum mongolicum)和猪毛菜(Salsola collina)为两大优势种。对照和养分添加处理下,物种组成较为丰富,主要有沙生针茅(Stipa glareosa)、骆驼蓬(Peganum harmala)、隐子草(Cleistogenes squarrosa)、冰草(Agropyron cristatum)、碱韭(Allium polyrhizum)和兔唇花(Lagochilus ilicifolium)(图1)。
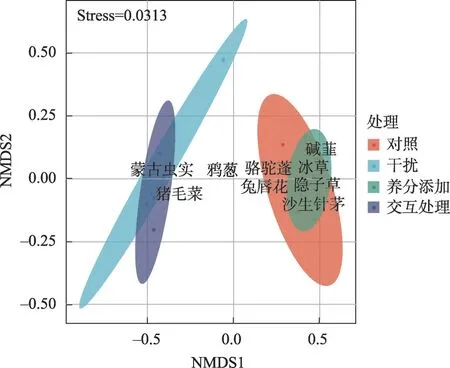
图1 养分添加、干扰及交互处理下植物群落主要物种组成Fig.1 Effects of nutrient addition,disturbance and interaction treatment on plant community main species composition
不同处理下物种丰富度和β多样性存在极显著差异。与对照相比,干扰和交互处理显著降低了物种丰富度和β多样性,养分添加处理下物种丰富度和β多样性无显著变化(P<0.01,图2)。
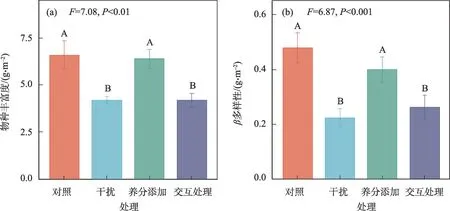
图2 养分添加、干扰及其交互处理对植物群落物种丰富度和β多样性的影响Fig.2 Effects of nutrient addition,disturbance and interaction treatment on plant community species richness and β diversity
与对照相比,干扰处理对土壤性质无显著影响,养分添加和交互处理对土壤SWC、TNC 和TCC无显著影响,但极显著降低了土壤pH,增加了EC(P<0.01,表2)。

表2 不同处理下土壤性质的变化Tab.2 Changes in soil properties under different treatments
2.2 养分添加和干扰对物种和群落功能性状的影响
不同处理对优势物种沙生针茅、骆驼蓬、猪毛菜和蒙古虫实的性状具有显著影响。与对照相比,养分添加处理下,沙生针茅的LDMC 显著降低(P<0.1,图3c),但SLA 和LNC 显著增加(P<0.05,图3d;P<0.1,图3e)。干扰处理下骆驼蓬的LDMC 显著增加(P<0.1,图4c),养分添加处理下,SLA极显著增加(P<0.01,图4d),LNC显著增加(P<0.05,图4e)。干扰和交互处理下猪毛菜株高极显著增加(P<0.001,图5a),养分添加、干扰和交互处理下SLA极显著增加(P<0.01,图5d),养分添加和交互处理下的LNC显著高于干扰处理下的LNC(P<0.05,图5e)。交互处理下,蒙古虫实的株高显著增加(P<0.05,图6a),干扰和交互处理下的LDMC显著高于养分添加处理下的LDMC(P<0.05,图6c),养分添加和交互处理下的SLA显著增加(P<0.05,图6d),交互处理下的LNC极显著增加(P<0.001,图6e),干扰处理下的LCC 显著高于交互处理下的LCC(P<0.05,图6f)。

图3 养分添加、干扰及其交互处理对沙生针茅性状的影响Fig.3 Effects of nutrient addition,disturbance and interaction treatment on functional traits of Stipa glareosa
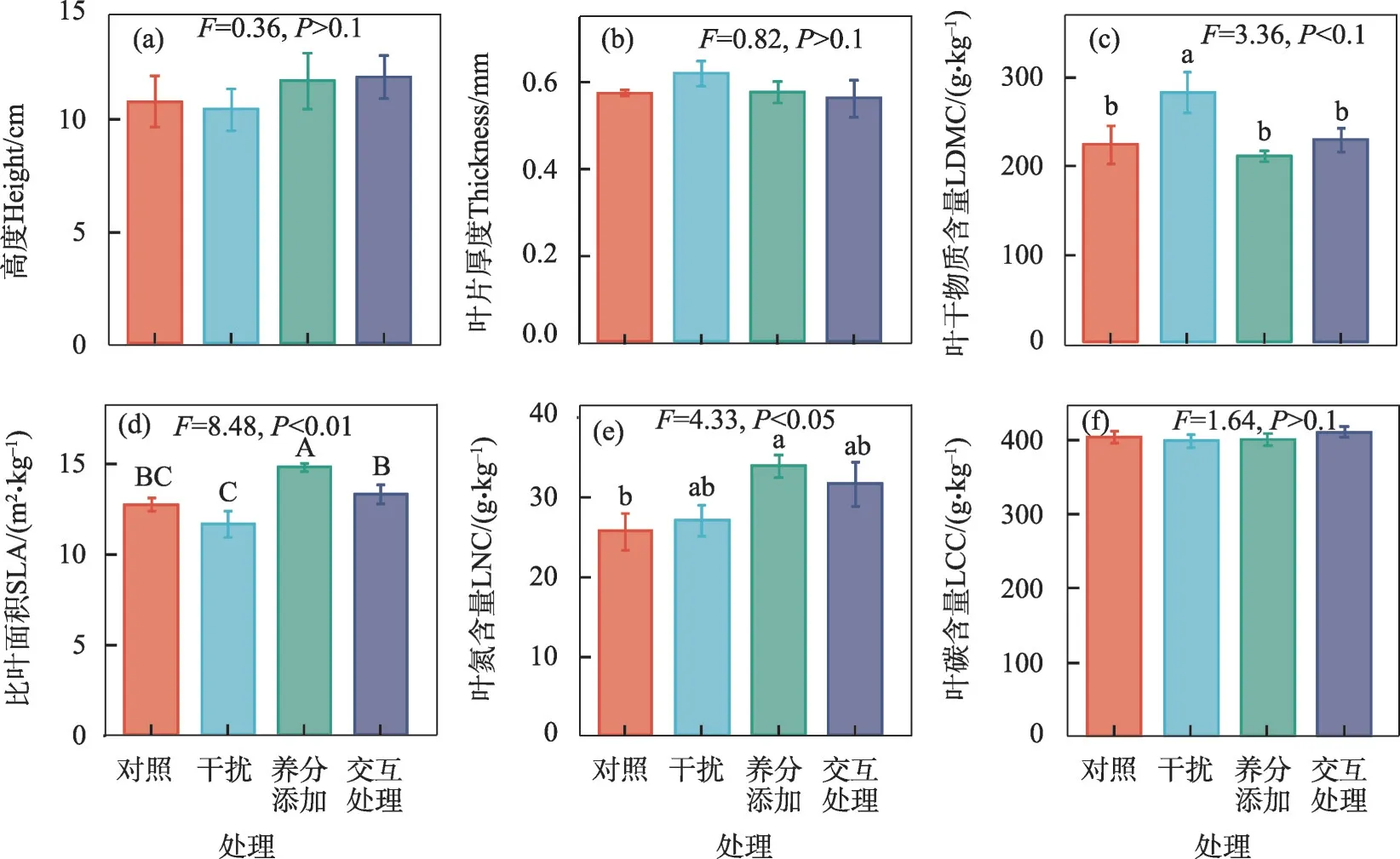
图4 养分添加、干扰及其交互处理对骆驼蓬功能性状的影响Fig.4 Effects of nutrient addition,disturbance and interaction treatment on functional traits of Peganum harmala

图5 养分添加、干扰及其交互处理对猪毛菜功能性状的影响Fig.5 Effects of nutrient addition,disturbance and interaction treatment on functional traits of Salsola collina

图6 养分添加、干扰及其交互处理对蒙古虫实功能性状的影响Fig.6 Effects of nutrient addition,disturbance and interaction treatment on functional traits of Corispermum mongolicum
不同处理对群落高度,群落叶干物质含量和群落比叶面积具有显著影响(P<0.05,图7),对群落厚度,群落叶氮含量和群落叶碳含量具有极显著影响(P<0.01,图3)。与对照相比,交互处理下CWMHeight显著增加(P<0.05,图7a),CWMThickness极显著增加(P<0.01,图7b)。干扰和交互处理下CWMLDMC显著降低(P<0.05,图7c);养分添加和交互处理下CWMSLA显著增加(P<0.05,图7d)。养分添加、干扰及其交互处理均显著增加了CWMLNC(P<0.001,图7e),干扰和交互处理下CWMLCC极显著降低(P<0.001,图7f)。
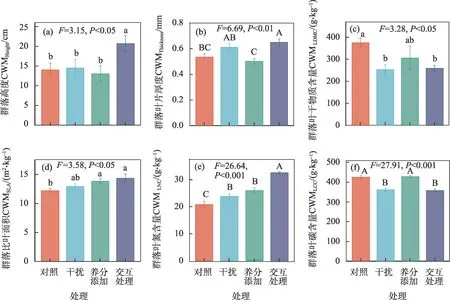
图7 养分添加、干扰及其交互处理对植物群落功能性状(CWM)的影响Fig.7 Effects of nutrient addition,disturbance and interaction treatment on plant community functional traits(CWM)
2.3 土壤性质和群落功能性状的主成分分析及其相关关系
土壤性质的主成分分析结果表明,pH和TNC在第一主成分上载荷较高,TCC和SWC在第二主成分上载荷较高,而EC 在第一和第二主成分上均具有较高的载荷。第一主成分的解释率为45%,第二主成分的解释率为27.1%,累计解释率为72.1%(图8a)。群落功能性状的主成分分析结果表明,CWMLCC,CWMThickness和CWMLNC在第一主成分上载荷较高,CWMHeight在第二主成分上载荷较高,而CWMLD-MC和CWMSLA在第一和第二主成分上均具有较高的载荷。第一主成分的解释率为50.1%,第二主成分的解释率为20.7%,累计解释率为70.8%(图8b)。回归分析表明,土壤PC1 和CWMLDMC之间具有极显著的正相关关系(P<0.01,图9c),与CWMSLA和CWMLNC之间具有极显著的负相关关系(P<0.01,图9d;图9e)。
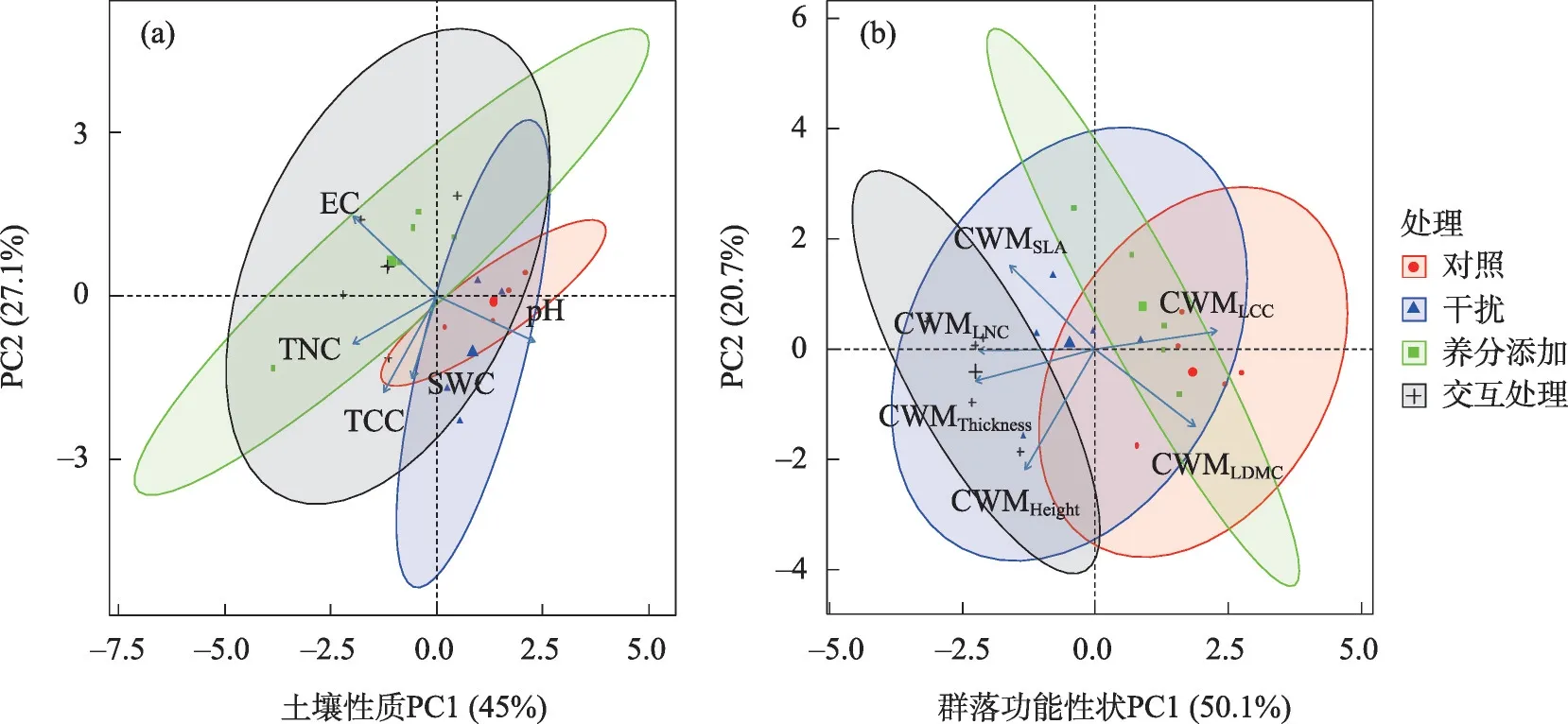
图8 不同处理下植物群落功能性状(CWM)的主成分分析(PCA)Fig.8 Principal component analysis(PCA)of plant community functional traits(CWM)under different treatments

图9 土壤性质与群落功能性状(CWM)的关系Fig.9 Relationship between soil properties and community functional traits(CWM)
2.4 养分添加和干扰对群落构建机制的影响
不同处理间NST 指数存在极显著差异(P<0.001,图10a)。干扰和交互处理下的NST指数小于0.5,而对照和养分添加处理下的NST 指数大于0.5。与对照相比,养分添加处理下的NST指数显著减小且显著大于干扰和交互处理下的NST 指数。这些结果表明随机性过程主导了对照和养分添加处理下的群落构建,而确定性过程主导了干扰和交互处理下的群落构建。不同处理间Raup-Crick相异指数存在极显著差异(P<0.001,图10b)。与对照相比,3种处理下的相异指数均小于0且显著减小。养分添加处理下的Raup-Crick指数显著减小且显著大于干扰和交互处理下的Raup-Crick指数。这些结果表明确定性过程对养分添加、干扰和交互处理下的群落构建有更大影响。

图10 养分添加、干扰及其交互处理下的群落构建过程Fig.10 Effects of nutrient addition,disturbance and interaction treatment on community assembly process
3 讨论
以往的研究表明,养分添加对物种丰富度有消极的影响,这与土壤酸化,光照限制以及低丰度物种的随机丧失有关[4,7,25]。在本研究中,养分添加对物种丰富度无显著影响,可能是由于本研究中试验处理的时间较短所致。荒漠草原地区土壤质地疏松,持水和保肥能力较差,贫瘠的土壤和较少的降水量使该区域植物受到养分和水分的双重限制[32]。当地植物在长期自然选择和进化过程中,已经适应了当地的生态条件,植物的生态幅即耐性范围在短时期内不会因养分添加而改变,因此短时期内新物种的建立和旧物种的丢失较少,物种丰富度的变化不明显。物种丰富度响应干扰的变化模式一般为驼峰曲线(中度干扰理论)[6],即轻度和重度干扰下物种丰富度均会降低,中度干扰下物种丰富度最高。本研究中,干扰和交互处理显著降低了物种丰富度,这与以往的研究结果相符[6,33],即重度干扰对物种丰富度会产生强烈的消极影响。β多样性揭示了群落之间物种组成的差异[4,33],受到局域环境条件特别是土壤因子的强烈驱动。以往的研究表明,养分添加会显著降低植物群落的β多样性[4,34],本研究中,养分添加处理下β多样性虽有所下降但无显著变化,表明短期养分添加虽然会导致群落物种组成更为相似,但影响程度较轻。β多样性在干扰和交互处理下显著下降,这与以往的研究结果相符[33],即重度干扰下群落物种组成更为相似。植被-土壤互馈表明,植物的生长发育与土壤性质的演变之间是互馈作用,土壤为植物生长提供水分和矿质营养,进一步决定植物群落类型及其物种组成,而植被又是土壤有机质最主要的来源[35]。在本研究中,养分添加和干扰处理通过影响土壤性质而进一步改变了群落物种组成和多样性。与之前大量研究结果一致[36-37],养分添加显著减小了土壤pH,显著增加了土壤EC,但土壤LNC 却无显著变化,这一方面可能是由于本研究中添加的养分量较少,另一方面可能是由于试验处理的时间较短;干扰处理在根本上改变了土壤表层特性,使土壤质地更加疏松,增加了土壤孔隙度,这种土壤条件有利于演替早期先锋种一年生植物猪毛菜和蒙古虫实的生长繁殖,因为其具有生长快、种子产量大、较高的扩散能力等特点;在交互处理下,土壤中养分含量增加,这将促进干扰后先锋植物更快速地生长。干扰相当于对局域群落内的物种施加了强烈的定向环境过滤器,只有那些适合干扰后土壤条件的物种才能在早期定殖并繁殖。
植物功能性状反映了植物对外界环境变化的响应和适应策略,体现了植物在资源获取效率和投资间的内在权衡关系[14,16]。例如,一年生植物将更多的资源用于叶面积的投入,因而用于结构建成的资源较少,而多年生植物则相反,因此SLA与LDMC之间存在负相关关系,之前的研究也证明了反映不同生态策略的各个叶功能性状之间存在显著的相关性[13]。本研究中,干扰和交互处理下,“快速投资-收益”(获取)型策略的一年生物种猪毛菜和蒙古虫实是优势种,叶功能性状的特点为:低的LDMC,大的SLA 和高的LNC,这种功能性状的组合方式允许它们能够快速利用环境中的有限资源并实现更强的碳同化,在短时期内实现生物量的快速增长。与此相反,对照和养分添加处理下,多年生优势种针茅和骆驼蓬的生态策略倾向于“缓慢投资-收益”(保守)型,叶功能性状的特点则和获取型策略的物种相反,这种功能性状组合使其对不良环境条件特别是养分和水分的亏缺具有更强的抗逆性[11,15]。养分富集的环境条件促使一年生物种将更多的资源用于叶面积投资而减小LDMC,而干扰环境下骆驼蓬和蒙古虫实LDMC的增大可能有助于增强其对不利环境的抵抗能力。植物高度通常反映了其对光资源竞争能力的强弱,叶片厚度与植物的保水能力相关,低C:N的特征有利于植物快速的营养生长[38]。本研究中,干扰和交互处理环境下获取型植物猪毛菜和蒙古虫实高度的显著增加,可能是因为物种组成单一、种间竞争减少所致,高度的增加也增强了它们对光能更强的获取能力,同时,更厚的叶厚有利于获取型物种在干旱的环境条件下储存水分。与以往的研究结果不同的是[39],与对照相比,养分添加处理下CWMLDMC虽有下降但变化不显著,由于荒漠草原地区受到养分和水分的双重限制,因此群落物种长期进化适应形成的抗逆机制,即倾向于将更多的资源用于结构性投资的特点可能不会在短期内发生变化。干扰和交互处理下更高的CWMHeight、CWMThickness、CWMLNC和CWMSLA表明,群落内优势物种对光照资源的利用更为充分,更厚的叶厚和更大的叶面积使获取型植物实现更强的光合作用,因此,猪毛菜和虫实作为先锋植物能在短期内大量生长繁殖。相比较而言,CWMLDMC和CWMLCC在对照下更大,揭示了当地植物长期适应养分和水分共同限制环境条件而形成的保守生态策略,并且这种策略不会因短期的养分添加而显著改变。此外,本研究的结果表明,土壤性质的改变在一定程度上可以指示CWMLD-MC、CWMLNC和CWMSLA的变化,揭示了基于MRH假说的群落功能多样性与土壤条件的密切联系[17,40]。
β多样性的变化可用来衡量群落构建确定性和随机性过程的强弱,并与生态系统功能相联系[41]。本研究中,物种是否在群落内发生以及相对丰度受到养分添加和干扰的影响。有研究表明,氮添加增强了定向的环境过滤作用,导致植物β多样性随氮的输入而线性下降,因此氮富集下的植物群落构建由确定性过程主导[4],这与本研究的结果一致。与养分添加处理相比,干扰处理下群落物种组成更为单一,环境筛选作用更强,因而植物β多样性更低,群落构建更倾向于确定性过程。此外,对照处理下群落构建倾向于随机性过程,这表明荒漠草原草本群落在自然状态下容易受到出生、死亡、迁移和扩散事件等随机性过程的影响。综上所述,干扰和短期养分添加均会影响当地植物群落原有的构建机制,且干扰的影响更为显著,这些结果表明,荒漠草原群落内物种的稳定共存以及维持受到全球气候变化和人类活动的强烈影响,进而改变生态系统结构和功能。
本文研究了荒漠草原群落物种组成、土壤理化性质、功能性状和群落构建机制对养分添加和干扰的响应,但遗憾的是,研究的时间尺度较短,今后的研究将持续关注不同处理下地上和地下群落的长期动态,并观测干扰后试验样地的恢复趋势,以求更加系统和清楚地阐明荒漠草原群落对养分富集和干扰的响应与适应机制,从而为荒漠草原生态系统的保护与恢复提供一定的理论依据。
4 结论
荒漠草原草本群落物种多样性、功能性状、土壤理化性质和群落构建过程对养分添加、干扰及其交互处理存在差异性响应,具体表现为:
(1)沙生针茅和骆驼蓬是对照和养分添加处理下的优势物种,猪毛菜和蒙古虫实是干扰和交互处理下的优势物种。干扰和交互处理显著降低了群落物种丰富度和β多样性,养分添加对群落也有类似的影响,但程度较轻。养分添加和交互处理显著降低了土壤pH,增加了EC。
(2)与对照相比,养分添加减小沙生针茅的LDMC,增加沙生针茅、骆驼蓬、猪毛菜和蒙古虫实的SLA 以及沙生针茅和骆驼蓬的LNC,干扰增加猪毛菜和蒙古虫实的Height,骆驼蓬和蒙古虫实的LDMC 以及猪毛菜和蒙古虫实的SLA。养分添加显著提高了CWMSLA和CWMLNC,干扰处理显著提高了CWMLNC,降低了CWMLDMC和CWMLCC,而交互处理显著提高了CWMHeight、CWMThickness、CWMSLA和CWMLNC,降低了CWMLDMC和CWMLCC。主成分和回归分析表明,土壤性质与CWMLDMC呈显著正相关关系,与CWMSLA和CWMLNC呈显著负相关关系。零模型分析结果表明,对照下的群落构建趋向于随机性过程,干扰和交互处理下的群落构建趋向于确定性过程,养分添加处理下的群落构建趋向于较弱的确定性过程。
(3)干扰和养分添加促使荒漠草原群落优势种由保守型策略的物种向获取型策略的物种转变,干扰的影响程度更强。针茅等多年生植物,具有保守型策略,在水分和养分双重限制的自然环境中具有更强的抗逆性,从而占据优势;猪毛菜等一年生植物,具有获取型策略,作为先锋植物,能快速抢占干扰后群落的生态位,在土质疏松的早期环境中定殖,并大量生长繁殖。不同处理影响局域环境条件特别是土壤pH 和EC,进一步改变了群落物种组成和功能性状。
(4)干扰和养分添加后环境特别是土壤条件的筛选决定了群落内物种的发生及其相对丰度,进而影响生态系统功能和群落构建机制。优势物种性状和群落性状之间的权衡反映了物种和群落整体在资源获取和分配中的权衡,与物种寿命和采取的生态策略密切相关。
