三氟苯嘧啶在扬稻6 号和南粳46 水稻上的内吸与传导能力比较
2023-06-20徐王瑾杨春梅缪心忆范添乐孟志远陈小军
徐王瑾, 杨春梅, 缪心忆, 范添乐, 孟志远, 陈小军
(扬州大学 植物保护学院/农业与农产品安全国际合作联合实验室 (扬州大学),江苏 扬州 225009)
三氟苯嘧啶是杜邦公司 (现科迪华) 研制的新型介离子嘧啶酮类杀虫剂,为烟碱乙酰胆碱受体抑制剂,现已在亚太地区广泛登记[1]。三氟苯嘧啶主要用于水稻害虫防治,也可用于棉花、玉米和大豆等作物。田间试验证实,三氟苯嘧啶能高效防治稻飞虱和叶蝉等害虫,速效性好,且持效期长[2]。在生产上,作用于烟碱型乙酰胆碱受体的杀虫剂如呋虫胺、吡虫啉和啶虫脒等在田间防治稻飞虱方面发挥了关键作用[3-4],然而,由于这些杀虫剂的滥用,其对稻飞虱的防治效果迅速下降[5]。新烟碱类杀虫剂作用于昆虫烟碱乙酰胆碱受体(nAChR),能引起急性兴奋性症状,而三氟苯嘧啶是对nAChR 有抑制作用的杀虫剂,与新烟碱类乙酰胆碱受体激活剂靶点相同,但作用相反。三氟苯嘧啶可导致稻飞虱中毒而昏睡,有较高的选择性,对包括传粉媒介在内的非目标生物体的影响也很小[6]。
不同水稻品种因生态和生理上存在差异,从而会导致其对外源物质的吸收和转运有所不同,同时也会影响农药到达作用靶标的时间[7],因此,不同品种的水稻对农药的吸收和利用存在较大差异。此外,不同品种的水稻对氮的利用率也不同。在低氮胁迫下,氮高效利用品种仍能够获得较高的有效穗数和千粒重,从而达到高产[8]。关于不同品种水稻对农药的利用率已有一些研究报道,如陈志廷等研究了华南地区51 个水稻品种对呋虫胺的利用率,找到了对呋虫胺具有高效吸收利用和定向积累于病虫害为害部位的主推水稻品种,有利于实现农药的减施增效[9]。农药在植物体内的内吸传导特性与其施用方法和施用效果密切相关[10]。内吸性农药可通过植物的根系和叶片吸收,在植物体内达到一定剂量后才能有效地防治有害生物[11]。扬稻6 号主要适宜生长在长江中下游及南方一季中籼稻地区,目前作为我国两系杂籼应用面积最大的恢复系[12],而南粳46 是江苏省育成的优质高产晚粳品系[13],这两种水稻在生产上都有重要的应用价值。然而,不同水稻品种之间理化性质差异较大,可能会影响其对农药的内吸传导特性。合理利用和改善农药在植物体的内吸传导特性,可大幅提高农药在靶标部位的积累并减少农药对环境的污染[14]。
目前,关于三氟苯嘧啶的研究主要集中在水稻生态系统的残留及消解规律方面[15-16],鲜有关于不同水稻品种间对三氟苯嘧啶内吸传导差异的研究报道。鉴于此,本研究以扬稻6 号和南粳46 作为供试水稻,探究了三氟苯嘧啶在不同品种水稻中内吸、传导能力的差异,以期为提高农药利用率提供数据支持和研究思路。
1 材料与方法
1.1 仪器与试剂
日立LC-2000 高效液相色谱仪 (日本Hitachi公司);SB25-12DTN 超声波清洗机 (宁波新芝生物科技公司);AUY120 电子天平 (日本岛津公司);RXZ 型智能人工气候箱 (宁波江南仪器厂);H/T-16MM 台式高速离心机 (湖南赫西仪器装备公司);Milli-Q 超纯水器 (美国Millipore 公司);HY-2 漩涡混匀仪 (上海仪电科学仪器公司);DCY-12S 水浴式氮吹仪 (青岛海科仪器公司);CX21 生物显微镜 (日本奥林巴斯株式会社);接触角测量仪 (美国KINO 公司)。
97%三氟苯嘧啶 (triflumezopyrim) 原药,由扬州大学农药实验室研制并提纯;乙腈 (色谱纯,美国Tedia 公司);其余试剂为国产分析纯。
1.2 试验材料
供试水稻品种:扬稻6 号 (YD 6),籼型常规水稻,购自江苏里下河地区农业科学研究所;南粳46 (NJ 46),粳型常规水稻,购自江苏省农业科学院粮食作物研究所。挑选粒大饱满均一的上述两种水稻种子,用无菌水清洗后置于无菌水中浸泡24 h,用浸湿的纱布包裹后置于恒温培养箱中(30 ℃黑暗恒温) 催芽,保持纱布湿润。待水稻种子根芽分化明显后,挑选长势一致的水稻苗移栽至96 孔板中,置于黑盒中水培培养。
1.3 试验方法
1.3.1 施药剂量及方式 根据大田施药量配制三氟苯嘧啶药液[17],供试质量浓度为2.5 和5 mg/L,并在药液中加入体积分数为0.1%的吐温-80。将水稻幼苗置于水培营养液中培养,其中水培营养液参照Hoagland 营养液配方[18]配制。水稻幼苗在光/暗周期为16 h/8 h、昼/夜温度为30 ℃/28 ℃和昼/夜湿度为60%/80%的人工气候箱中生长。在水稻3 叶期时采用以下两种施药方式处理。所有样品在分析前均置于 -20 ℃冰箱中保存。
1) 水培法:待水稻长至3 叶期,将96 孔板从黑盒中取出,用纯水冲洗水稻根部至无水滴后,将其浸于2.5 mg/L 的三氟苯嘧啶药液中继续培养,随后分别于2、24、48 和72 h 时在每组中随机取出水稻30 株和培养液10 mL,依次用甲醇和纯水冲洗水稻根部各3 次。将各组水稻植株分为根、茎和叶3 部分并称重。每处理重复3 次。
2) 浸叶法:待水稻长至3 叶期,将上部同一高度的叶片浸渍于5 mg/L 的三氟苯嘧啶药液中20 s,分别于叶片取出后2、24、48 和72 h 时在每组中随机取出水稻30 株和培养液10 mL,依次用甲醇和纯水冲洗水稻根部各3 次。将水稻植株分为处理叶、茎叶和根3 部分并称重,每处理重复3 次。
1.3.2 样品前处理 水样的前处理采用液液萃取法。取10 mL 过滤后的水样,用5 mL 乙酸乙酯振荡提取,加入2 g 氯化钠和1 g 无水硫酸钠,在7500g下离心5 min。取1 mL 上清液过0.22 μm滤膜过滤,待高效液相色谱 (HPLC) 检测分析。
水稻样品的前处理采用QuECHERS 法[19]。取剪碎的水稻样品1 g,用10 mL 乙腈超声提取后,加入1 g 氯化钠和2 g 无水硫酸镁,振荡后在7500g下离心5 min。取上清液4 mL,加入100 mg吸附剂m(PSA) :m(C18) = 1 : 1,在7500g下离心5 min。取2 mL 上清液在温和的氮气流下蒸发至干燥,残留物用色谱纯乙腈定容至1 mL,用0.22 μm 滤膜过滤,待HPLC 检测分析。
1.3.3 HPLC 检测条件 Supersil ODS-B 色谱柱(4.6 mm × 250 mm,5 μm);流速1.0 mL/min;流动相为V(乙腈) :V(水) = 35 : 65 混合溶液;波长233 nm;进样量10 μL;柱温25 ℃。三氟苯嘧啶保留时间为9.61 min。
1.3.4 标准曲线绘制、添加回收试验及基质效应测定 将三氟苯嘧啶原药溶于色谱纯乙腈,配制成1000 mg/L 的母液,于4 ℃保存,备用。使用时将母液用色谱纯乙腈稀释,得到质量浓度分别为0.05、0.78、1.56、3.12 和12.5 mg/L 的三氟苯嘧啶标准溶液,按照1.3.3 节色谱条件测定。以三氟苯嘧啶质量浓度为横坐标,对应的峰面积为纵坐标,绘制标准曲线。在进行三氟苯嘧啶的添加回收试验时,设置3 个水平的三氟苯嘧啶标准溶液分别添加在两种水稻中,每个水平重复3 次,按照上述方法处理和测定样品,计算平均回收率及相对标准偏差 (RSD)。分别对不同浓度的混合标准溶液和基质混合标准溶液进行检测,按 (1) 式计算基质效应 (ME):
式中,km为基质标准曲线斜率,ks为纯溶剂标准曲线斜率。
当|ME| < 20%时为弱基质效应,可忽略而无需采取补偿措施;当20% ≤ |ME| ≤ 50%时为中等程度基质效应;当|ME| > 50% 时为强基质效应,须采取措施补偿基质效应[20]。
1.3.5 三氟苯嘧啶在两种水稻中内吸、传导差异机制研究
1) 根系吸附试验:将2 g 水稻根用去离子水反复冲洗后切碎,取1/2 样品在105 ℃下加热40 min 后冷却至室温,另1/2 样品不做处理,分别放入装有10 mL 0.01 mol/L CaCl2溶液的锥形瓶中,其中含有100 mg/L NaN3和不同质量浓度(1、2、3、4 和5 mg/L) 的三氟苯嘧啶溶液。将锥形瓶在25 ℃下以150 r/min 的速度摇动24 h,同时,设置对照溶液 (无水稻根) 以校正由药剂降解或摇动引起的损失。每处理重复3 次。吸附等温线根据 (2) 式进行拟合[21]。
其中cΔ是水稻根部对三氟苯嘧啶的吸附质量浓度(mg/L),cw是处理溶液中实际测量的三氟苯嘧啶的质量浓度(mg/L),kpw是从相应吸附等温线的斜率获得的植物-水分配系数。
2) 根系活力的测定:参照萧浪涛等[22]的方法,采用α-萘胺法测定。
3) 蒸腾速率的测定:利用电子天平迅速称量30 株水稻叶片的鲜重,重复3 次,10 min 后再称量1 次,其差值即为蒸腾水量。通过叶面积测量仪测定水稻叶片面积,根据 (3) 式计算蒸腾速率 (T)。
式中:m为蒸腾水量,g;S为水稻叶片面积,m2;t为蒸腾时间,h。
4) 含水量的测定:每个处理中随机选取30 株水稻,分为根、茎和叶3 部分并称其质量 (m1),然后放入烘箱60 ℃下烘至恒重后称其质量 (m2)。每处理重复3 次。按 (4) 式计算含水量 (W)。
5) 水稻根茎组织的观察:取新鲜三叶期水稻根和茎,徒手横切制片,并用0.1%结晶紫水溶液染色,在生物显微镜下观察各组织。
6) 接触角测定:通过接触角测量仪测量1.3.1节中用于浸叶处理的药液在两种水稻叶片正、反面的动态接触角,采用Young-Laplace 方程拟合法计算接触角。每处理重复3 次。
1.4 数据处理
利用IBM SPSS Statistics 25 对试验数据进行单因素方差分析,结合t检验比较处理间差异(P< 0.05)。采用Origin 2019 软件作图。转移因子(translocation factor,TF) 按(5) ~ (6)式计算得到。TF 值越大,表示三氟苯嘧啶在植物中具有更强的向顶或向下传导的能力[23]。
其中croot、cstem和cleaf分别表示水稻的根、茎和叶中三氟苯嘧啶的含量 (mg/kg);TFs/r和TFs/l分别表示三氟苯嘧啶从水稻根到茎秆以及从叶到茎秆的传导能力。
2 结果与分析
2.1 检测方法的线性范围、正确度与精密度
测定结果表明,在0.05~12.5 mg/L 范围内,三氟苯嘧啶的质量浓度与其峰面积间有良好的线性关系,线性方程为y= 45197x+ 1390,决定系数R2为0.9999。在此检测条件下,三氟苯嘧啶的检出限为0.006 mg/kg,定量限为0.02 mg/kg。添加回收试验结果表明:在0.05~2.5 mg/kg 添加水平下,三氟苯嘧啶的回收率为84%~103%,RSD为1.1%~9.2% (表1)。

表1 三氟苯嘧啶在水稻和水中的添加回收率及相对标准偏差 (n = 3)Table 1 Recoveries and relative standard deviations of triflumezopyrim in paddy rice and water (n = 3)
基质效应用于可视化样品基质对色谱中出现的数据信号的影响。在0.05~12.5 mg/L 范围内,对水稻各部分基质的标准曲线进行了线性评价。结果 (表2) 表明:基质对三氟苯嘧啶表现出轻微的抑制作用,|ME| 在1.3~5.4 之间,表明在所有基质中没有明显的基质效应。以上试验表明,本研究所建立的检测方法适用于检测水稻样品中的三氟苯嘧啶。

表2 三氟苯嘧啶在不同基质中的标准曲线比较Table 2 Comparison of standard curves of triflumezopyrim in different matrices
2.2 三氟苯嘧啶在两种水稻不同部位的吸收与分布
水培处理结果表明:在两种水稻的根、茎和叶中均能检测到三氟苯嘧啶。药剂处理后2 h,在两种水稻根部便均可检测出三氟苯嘧啶,同时在南粳46 的茎、叶中也检测出了三氟苯嘧啶,但在扬稻6 号中未检出,表明三氟苯嘧啶能快速被两种供试水稻的根系吸收,但南粳46 能将其快速向顶传导。在72 h 内,南粳46 茎中三氟苯嘧啶的含量均显著高于扬稻6 号,叶中的含量也高于扬稻6 号,但仅在2 和72 h 时差异显著。三氟苯嘧啶在两种水稻根中的情况与茎、叶中的相反,并且仅在72 h时差异显著 (图1)。

图1 水培条件下三氟苯嘧啶在两种水稻不同部位的含量Fig.1 Content of triflumezopyrim in different parts of two kinds of paddy rice under hydroponic culture
浸叶处理结果表明:仅能在水稻的处理叶和茎叶中检测到三氟苯嘧啶,并且两者含量差异不大,但南粳46 对三氟苯嘧啶的吸收能力在48 h 内显著高于扬稻6 号 (图2)。

图2 浸叶处理条件下三氟苯嘧啶在两种水稻不同部位的含量Fig.2 Content of triflumezopyrim in different parts of two kinds of paddy rice in the leaf soaking treatment
2.3 三氟苯嘧啶在两种水稻中的传导能力
不同品种水稻的根系和叶片对三氟苯嘧啶的吸收和传导存在较大差异,从水培处理的结果可以 (图3) 看出,扬稻6 号的根系对三氟苯嘧啶的吸收能力更强,但三氟苯嘧啶在南粳46 中向顶传导的速度更快。计算两种水稻TFs/r值 (表3) 得出,南粳46 茎部和叶部的TF 值明显大于扬稻6 号的,由此可知,三氟苯嘧啶在南粳46 的根部向顶传导能力要显著优于扬稻6 号。

图3 水培条件下三氟苯嘧啶在两种水稻中的分布Fig.3 Distribution of triflumezopyrim in two kinds of paddy rice under hydroponic culture

表3 水培条件下三氟苯嘧啶在两种水稻中的转移因子 (n = 3)Table 3 Translocation factor of triflumezopyrim in two kinds of paddy rice under hydroponic culture (n = 3)
浸叶处理结果 (图4) 表明,三氟苯嘧啶在两种水稻根中未检出,说明三氟苯嘧啶向下传导能力较弱。通过比较两种水稻的TFs/l值 (表4) 发现,两种水稻茎部的TF 值没有显著差异。

图4 浸叶处理中三氟苯嘧啶在两种水稻中的分布Fig.4 Distribution of triflumezopyrim in two kinds of paddy rice in the leaf soaking treatment
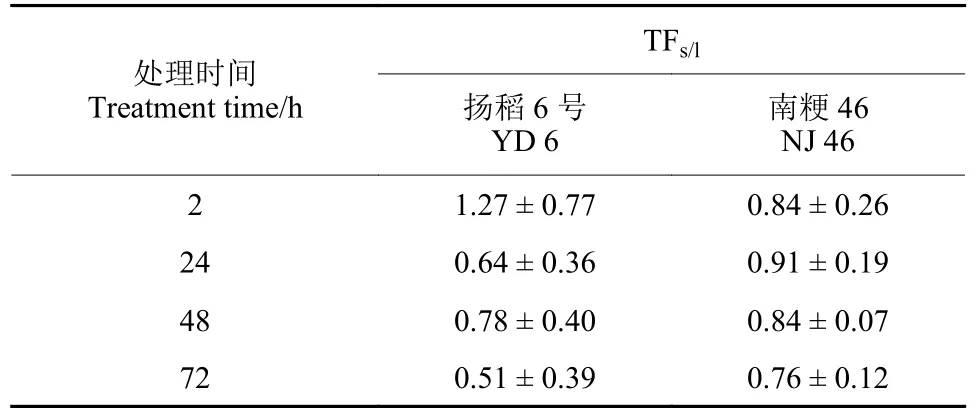
表4 浸叶处理中三氟苯嘧啶在两种水稻中的转移因子 (n = 3)Table 4 Translocation factor of triflumezopyrim in two kinds of paddy rice in the leaf soaking treatment (n = 3)
2.4 三氟苯嘧啶在两种水稻中内吸、传导差异机制
2.4.1 水稻根系和叶面吸收三氟苯嘧啶的途径
根系吸附研究中,水稻根部的三氟苯嘧啶含量随着水培溶液浓度的增加而增加,与吸附等温线高度拟合 (图5)。虽然加热增强了细胞膜的通透性,但并没有抑制细胞内的水分扩散[24]。同时,细胞中的主动转运受到抑制,导致共质通路减少。因此,死根主要通过质外体途径运输三氟苯嘧啶。本研究结果表明:两种水稻鲜根和死根中的吸收斜率相似,所以三氟苯嘧啶在这两种水稻的根系吸收是通过质外体和共质体途径进行的。对于浸叶处理,三氟苯嘧啶进入水稻体内需要克服叶片表面的角质层等屏障,同时扬稻6 号叶片上茸毛较多,南粳46 叶片较光滑[25],而这种水稻品种间叶片表面结构的差异,将导致叶片吸收农药含量不一致,也将导致其向下传导存在差异[26]。
图5 水稻根中三氟苯嘧啶的吸附等温线Fig.5 Adsorption isotherms of triflumezopyrim in paddy rice roots
2.4.2 三氟苯嘧啶在两种水稻根系中的吸收和传导差异机制α-萘胺法测定发现,南粳46 的根系活力显著高于扬稻6 号 (图6)。因此,南粳46 可以通过根系吸收更多的三氟苯嘧啶。通过根吸附试验发现,三氟苯嘧啶可以通过质外体途径进入植物体内,以植物体内的水为载体通过蒸腾作用不断从根中向上传导。测定两种水稻叶片蒸腾速率,发现南粳46 的蒸腾速率显著高于扬稻6 号,并且南粳46 茎中含水量要高于扬稻6 号 (图6),最终可能会导致叶片中三氟苯嘧啶含量的差异。此外,对比两种水稻根茎的切片 (图7),发现扬稻6 号根中的初生木质部明显多于南粳46,初生韧皮部也较发达,而在两种水稻茎中的情况则相反。木质部和韧皮部在植物体内主要起输导作用,农药通过质外体途径较经共质体途径向上传导更为容易,因为质外体途径能通过木质部的蒸腾作用将农药迅速向上传导[27]。这能很好地解释水培处理时三氟苯嘧啶在南粳46 中转移能力更强这一现象。

图6 两种水稻理化性质的差异Fig.6 Differences in physical and chemical properties of two kinds of paddy rice
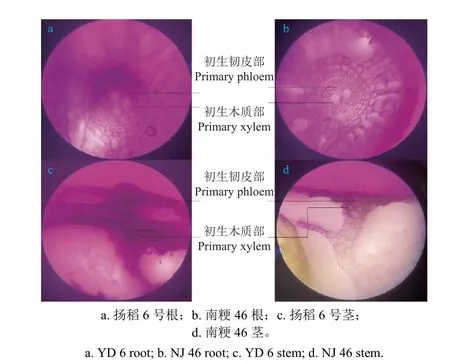
图7 水稻幼苗根茎的横切面生物显微镜图(放大倍数10 × 10)Fig.7 Biomicrograph image of the cross-section of the rhizome of paddy rice seedlings (magnification 10 × 10)
2.4.3 三氟苯嘧啶在两种水稻叶片中的吸收和传导差异机制 通过测定10 min 内液滴在两种水稻叶片上的动态接触角,发现南粳46 叶片正面上的接触角下降了6.5°,反面下降了3.5°。扬稻6 号叶片正面上的接触角下降了3.3°,反面下降了2.2°(图8)。液滴在南粳46 上的叶面接触角下降幅度明显高于扬稻6 号,表明南粳46 叶片吸收三氟苯嘧啶的能力要强于扬稻6 号。此外,扬稻6 号叶片含水量显著高于南粳46 (图6) ,且叶片较大,蒸腾作用较强,也会导致三氟苯嘧啶更难向下传导。

图8 三氟苯嘧啶溶液在两种水稻叶片正、反面的动态接触角Fig.8 Dynamic contact angles of triflumezopyrim solution on the front and back of two paddy rice leaves
3 结论与讨论
本研究结果表明:两种供试水稻经三氟苯嘧啶药液水培处理后,扬稻6 号的根对三氟苯嘧啶吸收多于南粳46,但南粳46 茎、叶中三氟苯嘧啶的含量更高,表明南粳46 具有更强的向顶传导能力;而水稻经浸叶处理后,南粳46 茎、叶中三氟苯嘧啶的含量明显高于扬稻6 号,但三氟苯嘧啶在南粳46 中向下的传导能力与扬稻6 号没有显著差异,三氟苯嘧啶在两种水稻中都表现出较弱的向下传导能力。
研究发现,由木质部介导的砷、镉从根系向地上部的转运是决定水稻、小麦茎秆和籽粒中砷、镉含量的一个关键过程[28]。此外,有研究表明,高氯酸盐可在植物体内或植物-水体系统中发生转移,并主要经木质部转移至地上部。受蒸腾作用影响,不同植物不同部位高氯酸盐含量差异巨大,并且叶片是高氯酸盐的主要累积部位。通过亚细胞试验发现,高氯酸盐主要分布在细胞可溶性组分中并且具有较高的移动性[29]。水溶性化合物多分布在植物组织的可溶性组分中,而三氟苯嘧啶和高氯酸盐均属于水溶性化合物[30]。
水稻吸收三氟苯嘧啶的能力与其理化性质有很大关系,根系活力和蒸腾速率等可反映其对农药的吸收和传导能力。Chiou 等[31]指出,对于辛醇-水分配系数 (Kow) 小于10 的有机物,水稻根部水分对于药剂的吸收贡献较大 (85%以上)。三氟苯嘧啶属于亲水性农药,Kow为1.24,因此,水稻根部含水量的差异会导致水稻对三氟苯嘧啶的吸收产生影响。此外,植物根系可以通过质外体和共质体途径吸收有机化合物,并通过木质部向上运输[32]。本研究中,南粳46 的根系活力显著高于扬稻6 号,茎中的初生木质部也要多于扬稻6 号,而大多数亲水化合物主要转移到木质部中[33],并且南粳46 茎部含水量更高,通过较强的蒸腾作用可以向上传导更多的三氟苯嘧啶[34]。因此,根系活力、蒸腾速率以及木质部的差异是三氟苯嘧啶在南粳46 中向顶传导能力优于扬稻6 号的主要原因。浸叶处理中,农药通过叶片吸收主要靠共质体途径,即从吸收部位通过胞间连丝或细胞间的渗透进入维管束组织,向上到达芽、叶等部位,向下进入根部,但有时因为环境条件、农药分子特性、植物体内物质等因素并不能由施药叶片向上或向下传递[35],蒸腾作用和叶片性状等差异是否会导致叶片吸收三氟苯嘧啶的含量不同以及影响向下传导的能力,需要进一步探究。此外,由于取样的时间较短,未能探究三氟苯嘧啶代谢方面的内容,值得深入研究以明确其在水稻生产中的安全性问题。虽然本研究仅采用了两种水稻品种,但扬稻6 号和南粳46 分别是扬州地区籼稻和粳稻的典型品种,由此可推测,三氟苯嘧啶能穿透水稻的根与叶表面以及快速的被植物吸收,并且能够双向传导,但具有更强的向上传导能力。稻飞虱主要刺吸水稻茎秆汁液,建议在进行田间防治时对准根茎处施药或者采用种子和土壤处理,在粳稻田中可以适当减少用量,从而提高农药的利用率。
