新疆野生桦褐孔菌生物学特性及化学成分分析
2023-06-20华兰兰时红玲李金玉霍向东
华兰兰,林 青,时红玲,王 娜,娄 恺,李金玉,霍向东
(1.新疆师范大学生命科学学院,乌鲁木齐 830054;2.新疆农业科学院微生物应用研究所/新疆特殊环境微生物实验室,乌鲁木齐 830091;3.新疆大学生命科学与技术学院,乌鲁木齐 830052)
0 引 言
【研究意义】桦褐孔菌(Inonotusobliquus)是一种白腐真菌,属于担子菌纲锈革孔菌科,主要生长于桦树的树干上[1],可耐受 -40℃的低温,主要分布于N 45~50°的寒冷地带,包括北美、芬兰、波兰、俄罗斯(西西伯利亚、东部、堪察加半岛),以及中国东北部(黑龙江省的小兴安岭和吉林省的长白山)和日本(北海道)[2-3]。作为一种药用真菌,桦褐孔菌含有多种生物活性成分,其提取物具有降血糖[2-4]、抗病毒[5]、抑菌[6]、抗氧化[7-8]、抗炎[9-10]、保肝[11-12]等作用。【前人研究进展】当前,关于桦褐孔菌菌核、菌丝体和发酵液活性物质的成分报道虽多,如桦褐孔菌多糖、桦褐孔菌素、桦褐孔菌醇、栓菌酸等[3],但对桦褐孔菌总体化合物种类的研究甚少。真菌所含有的大量有机化合物,不仅有药用价值,对真菌自身也起着重要的生态和生理作用[13]。液体发酵是一种快速、可行的替代方法,可以获得质量一致的桦褐孔菌菌丝体[14]。【本研究切入点】桦褐孔菌资源稀少,药用价值高,为加强该菌株的资源利用及相关产品的开发,需对桦褐孔菌的生物学特性及化学成分进行分析。【拟解决的关键问题】对新疆阿勒泰野生桦褐孔菌菌核进行培养物分离、纯化及鉴定,使用Biolog FF微孔板对菌丝体进行碳源利用分析,经液体培养,获得桦褐孔菌菌丝体和发酵液,采用气相色谱-质谱(GC-MS)法确定桦褐孔菌菌核、发酵菌丝体和发酵液的化学成分组成。
1 材料与方法
1.1 材 料
1.1.1 材 料
桦褐孔菌菌核采自新疆阿勒泰小东沟林场(47°56′N, 88°07′E) ,实验室分离获得纯菌株,菌株编号为HS819;植物凝胶和吐温40均购自索莱宝(Solarbio)公司,其余试剂均为国药分析纯试剂。
1.1.2 仪器与设备
HVA-85高压灭菌锅(日本);SW-CJ-2FD净化工作台(上海博讯实业有限公司);ZWY-2102C恒温培养振荡器(上海智城分析仪器制造有限公司);Thermo Presco17台式高速离心机(美国);DS-Ri2 Nikon显微镜(日本);GP306C-120-500电动组织研磨器(生工生物工程股份有限公司);Biostack Ready浊度仪 (美国);Microplate Reader读数仪(美国);SONICS VC130超声波破碎仪(美国);SHIMADZU SH-Rtx-Wax色谱柱(30 m*0.25 mm*0.25 μm),TQ8040NX三重四极杆型气相色谱-质谱联用仪(日本岛津)。
1.1.3 培养基
分离培养基(BA):桦树皮粉末15 g/L,琼脂20 g/L,水1 000 mL,121℃ 高压灭菌20 min,使用前加入氨苄青霉素0.1 g;斜面培养基:PDA培养基;PDA加富培养基:马铃薯200.0 g/L、葡萄糖25.0 g/L、蛋白胨5.0 g/L、酵母粉3.0 g/L、KH2PO41.0 g/L、琼脂25.0 g/L;种子培养液:葡萄糖30.0 g/L,蛋白胨5.0 g/L,MgSO40.5 g/L,KH2PO42 g/L,pH 自然;发酵培养液:葡萄糖20.0 g/L,蛋白胨4.0 g/L,MgSO41.0 g/L,KH2PO41.0 g/L,pH 自然;FF-IF 接种液:植物凝胶0.25%,吐温40 0.03%,121℃灭菌20 min。
1.2 方 法
1.2.1 分离纯化
选取新鲜的桦褐孔菌菌核,去除表面杂质,75%酒精清洗其外表面3次。采用组织分离法进行分离。在无菌条件下,切取5 mm × 5 mm的菌核组织块接入分离培养基, 置于28℃ 恒温箱中培养培养5 d;挑取菌落边缘,在PDA加富平板上进行纯化培养10 d,并将分离、纯化的菌株接种于PDA斜面,30℃遮光培养,待菌株长满斜面后,放置4℃保存。
1.2.2 液体发酵
生长于PDA加富平板上的菌落用打孔器打取直径1 cm的菌饼4块转置50 mL离心管中,研磨后,加入5 mL无菌水混匀,接入500 mL锥形瓶中(100 mL种子培养基),在30℃,150 r/min 条件下培养3.5 d后将种子液转置250 mL锥形瓶(100 mL发酵培养基),接种量为10%,150 r/min,28℃培养10 d。
1.2.3 形态学
观察菌株在固体平板上的生长特征,包括菌落形态、菌丝颜色、渗出物颜色。以30~45°斜插入无菌的盖玻片,28℃避光培养,待菌丝体蔓上载玻片后,取出载玻片用尼康显微镜对菌丝体进行形态学拍照。
1.2.4 基于ITS序列的桦褐孔菌进化
上海生工生物工程股份有限公司利用真菌ITS通用引物ITS1(5’-TCCGTAGGTGAACCTGCGG-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC -3’)扩增并测序。ITS rDNA 序列提交至GenBank数据库,登录号为 OK586158;BLAST在线比对分析,利用MEGA 11.0软件、以Neighbor-joining法(1000 bootstrap)构建系统进化树,分析同源关系。
1.2.5 碳源谱
制备Biolog FF板接种液[15],采用95种碳源Biolog FF微孔板分析桦褐孔菌菌株碳源利用情况。从PDA平板生长菌落刮取菌丝体,转置于含有500 μL FF-IF接种液的无菌EP管中,电动手持式研磨机破碎,收集的菌丝体10 000 r/min离心10 min,弃上清,再次加入FF-IF接种液,离心去上清,重复该步骤两次,将最终获取的菌丝体沉淀悬浮于装有4 mL接种液离心管中,静置去除菌丝团。用浊度仪将装有10 mL FF-IF接种液的Biolog FF标准比浊管的浊度调至100%后,加入菌丝体悬浮液将其浊度范围调至75%T左右,将此悬浮液倒入V型加样槽,迅速接种至Biolog FF板(每孔100 μL),28℃培养。
碳源利用活性:菌株利用碳源的能力,可用平均吸光值(AWCD)表示,通过计算和绘制菌株的AWCD值随时间的变化,得到菌株利用碳源的平均活性[16]。菌株代谢活性用590 nm下的吸光度值减去750 nm下的吸光度值表示,其中数值小于0时按0处理,计算方法:AWCD=Σ(C-R)/n,C为每个碳源孔的两波段吸光度差值,R为对照孔的吸光度值,n为培养基碳源种类数,研究中为95[17]。 碳源的平均吸光值(AWCD)能反映微生物代谢活性,是利用单一碳源的重要指标,能衡量菌株对于不同碳源的总体利用能力[18]。绘制AWCD随时间变化的曲线,并通过方差分析不同时间点样本间AWCD的差异。数据分析应用SPSS 24软件,绘图应用Origin 8.0软件。
1.2.6 GC-MS
样品前处理:取5 g样品,加入50 mL石油醚(沸点30~60℃),振荡混匀,超声处理15 min,8000 rpm离心3 min,收集上清液,减压浓缩至干,再加入1 mL石油醚溶解沉淀,经0.22 μm滤膜过滤后进行分析。
GC条件:分流进样,分流比10∶1;进样口温度为250℃,程序升温:初始温度为30℃,保持1 min,以5℃/min升至220℃,保持3 min,再以10℃/min升至250℃,保持5 min。
MS条件:EI电离源;载气为He;吹扫流量3 mL/min;电子能量70 eV;离子源温度200℃,接口温度250℃。全扫描模式,质量扫描范围为 50~600 amu;溶剂延迟2 min。
2 结果与分析
2.1 菌株HS819的形态学特征
研究表明,菌株HS819菌核无柄,表面有不规则沟痕,内部黄色至黄褐色;经组织分离,在BA板上菌丝呈白色,匍匐在培养基上;菌落初期白色,后逐渐变为浅黄至黄褐色,而边缘保持白色;菌丝在加富PDA平板上生长较缓慢,28℃培养15 d直径8.3 cm左右;背面有环横纹出现;在微分干涉学显微镜下(×400),菌丝粗壮,无可见孢子。图1
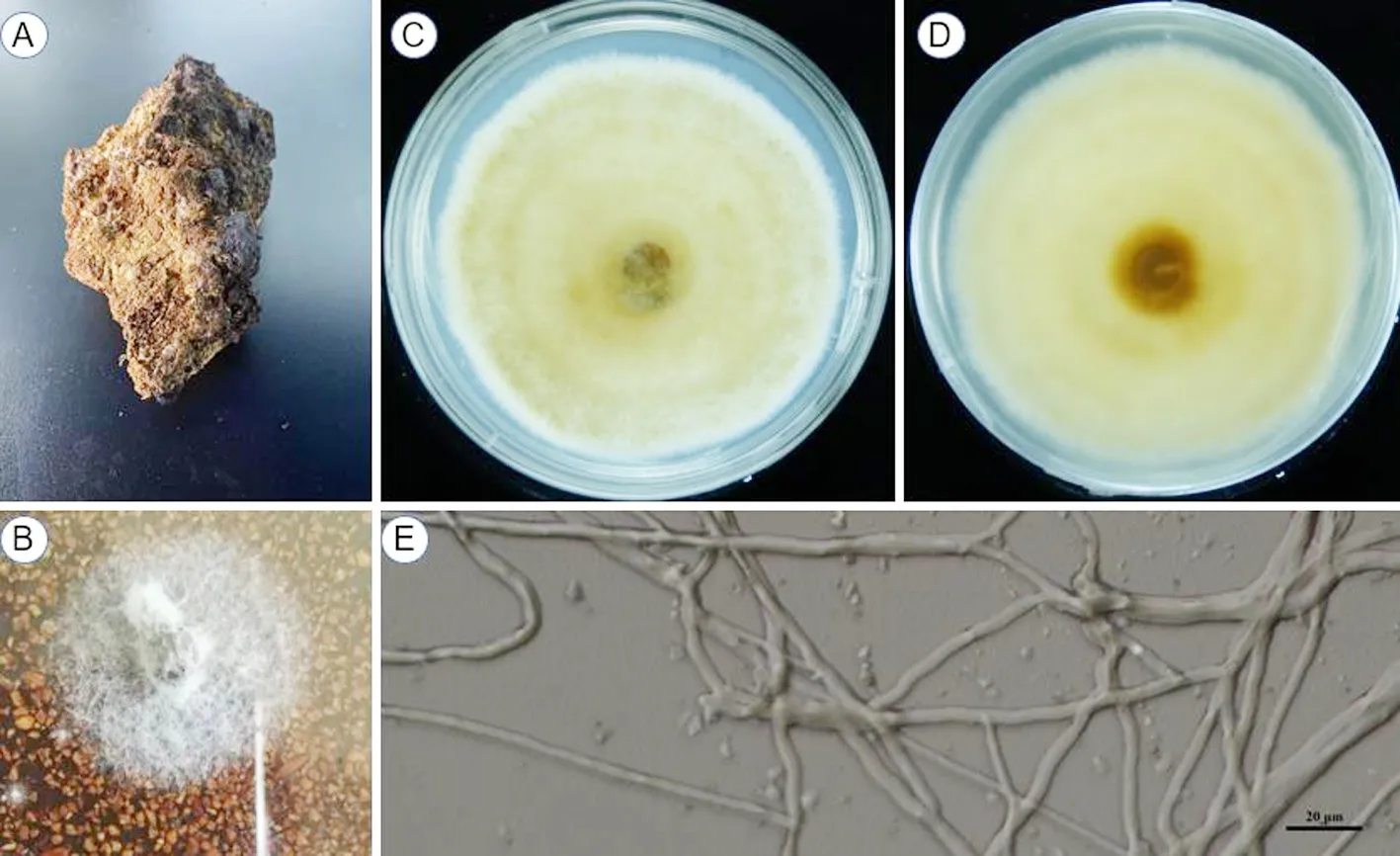
注:A:菌核;B:BA生长菌落;C:PDA加富平板生长菌落(正面);D:PDA加富平板生长菌落(背面);E:菌丝体(标尺=20 μm)
2.2 系统发育分析
研究表明,菌株HS819的测序 ITS rDNA长度为778 bp,在 NCBI 上BLAST的比对结果显示与GenBank 中OP019327、OP019325、KC312697、MZ484606等桦褐孔菌菌株的序列相似度达99% 以上,且系统发育树也表明该菌株与以上四株菌聚为一类。确认菌株HS819为桦褐孔菌。图2
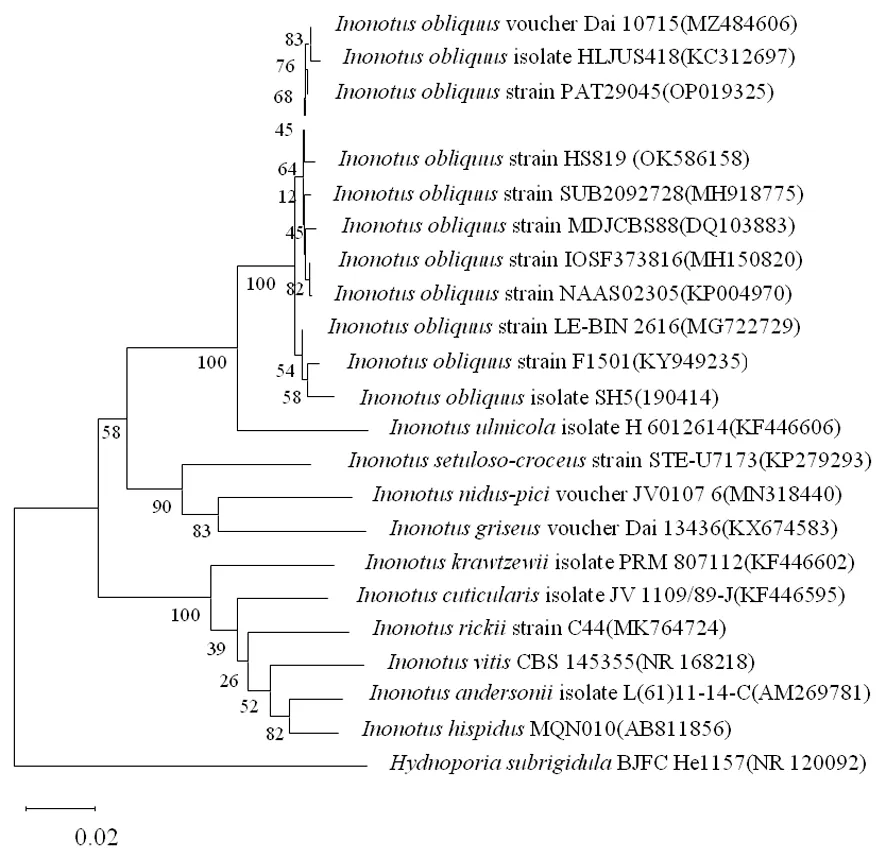
图2 基于ITS序列的系统发育
2.3 桦褐孔菌菌株HS819碳源谱特征
2.3.1 桦褐孔菌菌株HS819代谢碳源平均活性
研究表明,前7 d桦褐孔菌对碳源利用率低,随着培养时间延长,呈逐渐增加趋势,在第15 d利用率达到最高。图3

图3 桦褐孔菌菌株HS819利用 碳源平均吸光值变化
2.3.2 桦褐孔菌菌株HS819对碳源种类利用
研究表明,菌株HS819能代谢95种测试碳源中的33种,其中7种碳源能够被有效利用,17种碳源适度利用;其余碳源完全不能代谢,包括环式糊精(B1孔)、α-D-葡萄糖-1-磷酸(C1孔)和D-甘露醇(D1孔),赤藓糖醇(B4)等,孔内均无菌丝生长。表1

表1 桦褐孔菌菌株HS819在Biolog FF微孔板中的碳底物利用
2.4 野生桦褐孔菌菌核、菌株HS819发酵液及菌丝体化学成分(表2)
2.4.1 野生桦褐孔菌菌核中化学成分及分析
研究表明,桦褐孔菌菌核43种化学成分中含有酸类8种,烃类21种、酯类2种、醛类2种、醇类5种、其它5种。酸类化合物占整体化合物的比例最大,达到66.07%;其次是烃类占21.69%;醇类,醛类,酯类,分别占3.82%,2.03%,1.53%。酸类化合物主要有亚油酸(24.39%)、反油酸(21.15%)、棕榈酸(7.4%)等;角鲨烯(4.53%)是烃类的主要化合物;醇类化合物主要有二十七烷醇(1.55%)、叶绿醇(1.32%);醛类化合物仅有顺-7-十四烯醛和硬酯烷醛;脂类化合物主要为乙酸二十八脂(1.06%)和邻苯二甲酸异丁基环己基甲酯(0.47%)。其它化合物中二十烷基壬基醚(2.7%)、2,4-二叔丁基-苯酚(0.85%)等含量相对较高。
2.4.2 桦褐孔菌菌株HS819菌丝体中化学成分及分析
研究表明,发酵菌丝体39种化合物中含有酯类6类,烃类18种,酸类6种,醇类2种。酸类化合物占整体化合物的比例最大,达到60.03%,其次是酯类占30.72%,烃类、醇类分别占4.59%、1.04%。酸类中含量较高的是油酸(28.76%)、棕榈酸(18.51%)、反油酸(7.4%)等;脂类主要有 (Z)-十八碳-9-烯内酯(27.12%)、油酸乙酯(1.13%)、乙酸二十八烷基酯(1.06%)等;烃类主要有1-十九烯(0.42%)、α-衣兰油烯(0.2%)、正二十四烷(0.41%)、1,2-环氧十六烷(0.77%)等;醇类有二十七烷醇(0.72%)、1-二十三烷醇(0.32%);其它化合物中依兰酚(1.55%)的含量最高。
2.4.3 桦褐孔菌菌株HS819发酵液中化学成分及分析
研究表明,菌株HS819发酵液中38中化合物中含有烃类20种、醇类5种,酸类3种、酯类4种。烃类化合物占整体化合物的比例最大,达到54.01%;其次是醇类占8.24%,酯类与酸类分别占12.89%,7.72%。烃类中含量较高的是11-甲基二十烷(7.11%)、三十五烷(7.44%)、正三十六烷(6.58%)、正二十四烷(4.89%)等;醇类主要有二十七烷醇(3.05%)、叶绿醇(1.87%)、鲸蜡醇(0.78%)等;酯类有四十三烷基七氟丁酸酯(2.36%)、十八碳酸乙烯基酯(5.04%)、4-甲氧基乙酸十三烷基酯(4.43%);酸类主要有肉豆寇酸(3.06%)、棕榈酸(3.3%)、油酸(1.36%)等;其他化合物中依兰酚(8.39%)、2,4-二叔丁基-苯酚(2.16%)、十二烷基七聚乙二醇醚(2.57%)、肉豆蔻酰胺(2.1%)含量相对较高。
3 讨 论
桦褐孔菌菌株HS819的碳代谢指纹图谱表明,95种碳源中只有少量种类能被代谢,且这些碳源基本参与了生命体的三个重要的碳代谢途径。琥珀酸、琥珀酸甲基酯、α-酮戊二酸、L-天冬氨酸、L-谷氨酸参与三羧酸循环(TCA);戊糖磷酸途径(PPP)相关的一些碳源桦褐孔菌也可以使用,包括D-阿拉伯糖、L-阿拉伯糖、D-核糖和D-木糖;其余的碳源(D-葡萄糖醛酸、α-D-乳糖、麦芽糖醇、肝糖、D-果糖、龙胆二糖、D-纤维二糖、α-D-葡萄糖等)主要参与糖酵解途径。
碳源是微生物营养的基本组分,特定菌株同化某些碳源的能力可能与其在特定营养条件下的竞争力有关[17],例如与许多植物相关的真菌能分解并利用主要由纤维素和半纤维素组成的底物[19]。大型真菌的底物使用和代谢可表明其潜在的生态功能,了解菌株在自然界不同生态位条件下的营养适应性有助于选择培养基成分,以优化次级代谢产物的生产,例如为提高金针菇菌株的漆酶活性,利用 Biolog FF微孔板来优化培养条件[20]。桦褐孔菌可利用碳源中的α-D-葡萄糖、D-纤维二糖、D-木糖、D-半乳糖、L-阿拉伯糖、D-半乳糖醛酸、D-葡萄糖醛酸是纤维素或半纤维素的主要成分[21],这与桦褐孔菌主要生长于桦树上相一致。
研究发现,野生桦褐孔菌菌核、菌丝体及其发酵液在化学成分种类和含量上的差异明显,这可能是桦褐孔菌基因在自然环境下和液态发酵过程中差异性表达的结果[22]。例如1-碘-二十烷、1-碘癸烷及1,14-二溴十四烷仅存在于菌核中,其角鲨烯的含量是发酵液的1.1倍,而发酵液中的烃类相对含量远多于菌核和菌丝体,α-衣兰油烯则只在菌丝体中检测到;菌丝体中的酯质含量分别是菌核的20倍,发酵液的2.4倍,油酸乙酯、十六酸乙酯、乙酸三十烷酯、邻苯二甲酸二丁酯、(Z)-十八碳-9-烯内酯仅存在于菌丝体中,而十八碳酸乙烯基酯、四十三烷基七氟丁酸酯,4-甲氧基乙酸十三烷基酯则只存在于发酵液中;菌核中有机酸的种类多于菌丝体及发酵液,亚油酸和亚麻酸是菌核特有成分,油酸在菌丝体中的含量高达28.76%;顺-7-十四烯醛和硬脂烷醛则只在菌核中存在。
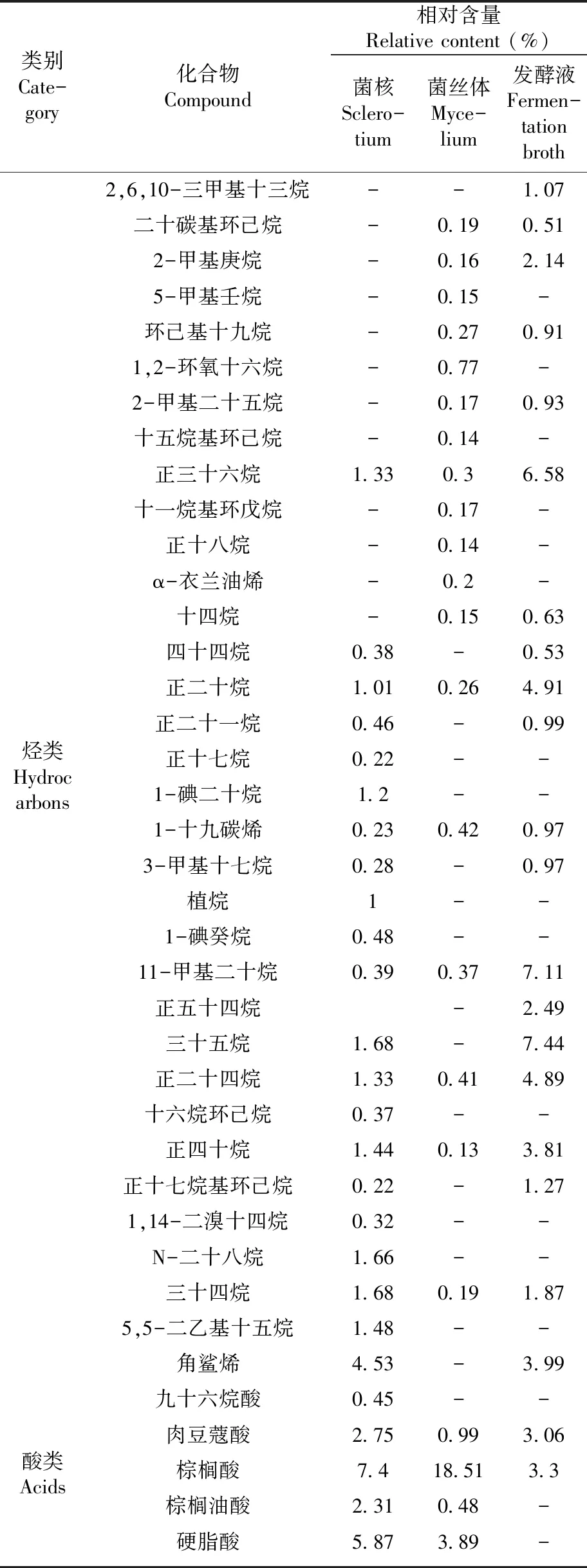
表2 野生桦褐孔菌菌核、菌株HS819 菌丝体、发酵液中化学成分的定性 和定量变化
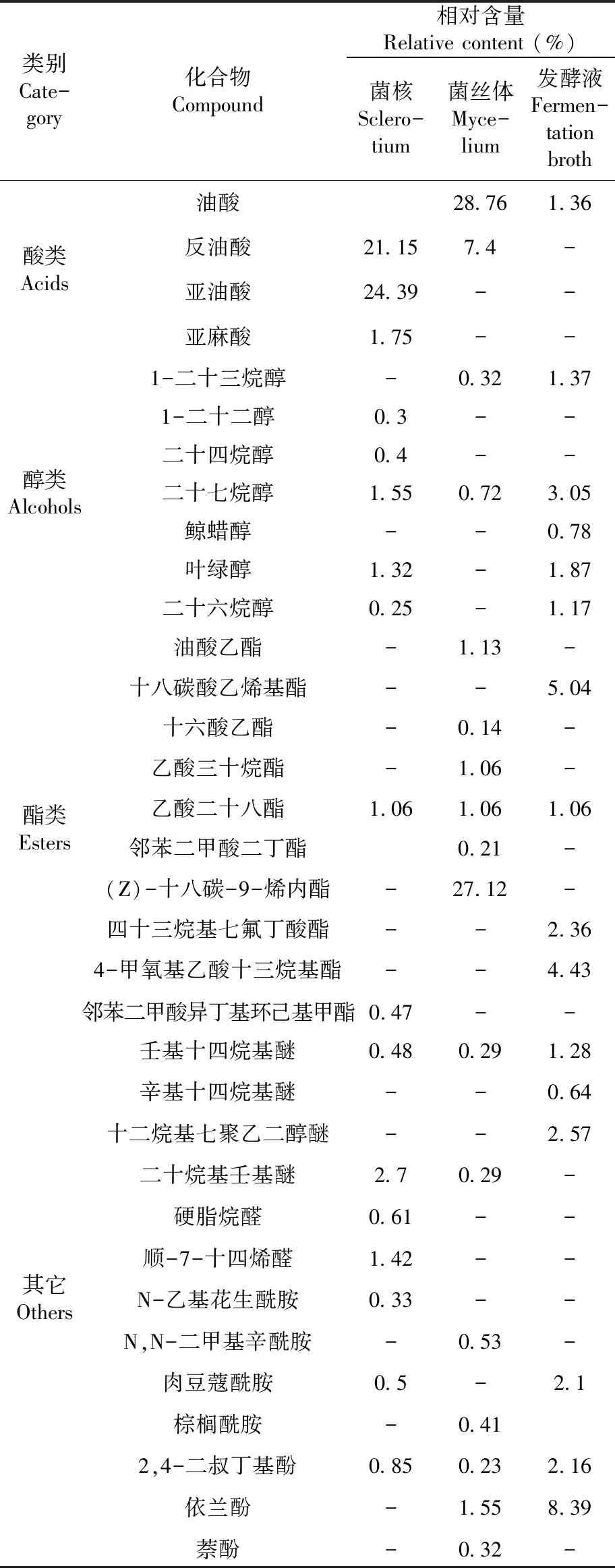
续表2 野生桦褐孔菌菌核、菌株HS819 菌丝体、发酵液中化学成分的定性 和定量变化
担子菌胞外酶酶解木质素所释放的低聚物是真菌和细菌生长的合适底物,为获取更多营养物质,真菌会抑制周围细菌的生长[23],该现象也可能在桦褐孔菌中存在。例如:菌核中所特有的有机卤素(1-碘二十烷、1,14-二溴十四烷及1-碘癸烷),有可能在木质素的降解中起到中介作用[24],也可能抑制相互竞争的真菌和细菌[23];菌核中所特有的醛类物质及其酸含量多于菌丝体,而这2类物质均有一定的抑菌活性[25],有研究表明高含量的油酸、亚油酸、棕榈酸还能够有效抑制腐生线虫[26]。在未来可尝试挖掘这些化合物的生态功能,为桦褐孔菌的生长和保护提供策略的同时也可开发新的化学植物保护剂[27]。
真菌在自然环境和液态发酵过程中基因的差异性表达可能与生物胁迫、生长环境等因素有关[28],如温度会影响真菌的脂肪酸组成[29-30]。本研究结果显示,野生菌核中不饱和脂肪的含量为49.6%,其中多不饱和脂肪酸的含量(亚麻酸、亚油酸)及单不饱和脂肪酸(油酸、棕榈油酸)的含量分别占总不饱和脂肪酸含量的52.7%,47.29%;菌丝体及其发酵液中不饱和脂肪酸均为单不饱和脂肪酸,含量分别为36.64%,1.36%。桦褐孔菌菌核生长在低温环境,菌丝发酵培养温度为28℃,这与随着培养温度的降低,脂肪酸的不饱和度增加的观点相一致[31-32],这些不饱和脂肪酸也可能通过脂质过氧化反应在降解木质素的过程中发挥作用[33]。此外,本研究菌核中反油酸的相对含量高达21.15%,是菌丝体的2.9倍,反式脂肪酸的增多与自然环境中菌株的营养缺乏有关[34]。在摇瓶培养过程中,菌丝体主要以菌块、菌球形态存在,易于传氧、传质,菌丝体为营养生长阶段,酶系活跃。菌核直接长于大气中,靠菌托传递营养,为生殖生长阶段,酶系相对单一[28]。
4 结 论
新疆桦褐孔菌菌株HS819对D-核糖、水杨苷、D-纤维二糖等7种碳源的代谢能力最强;酸类是野生菌核及菌丝体的主要成分,分别占总成分的66.07% 和60.03%,在发酵液中,烃类为主要成分,占54.01%。
