9个欧洲李品种自交不亲和S-RNase基因的克隆及序列分析
2023-06-20王亚铜玉米提玉素甫耿文娟章世奎孙召展王绍鹏王尚栋樊国全
王亚铜,玉米提·玉素甫,耿文娟,章世奎,孙召展,王绍鹏,王尚栋,樊国全
(1.新疆农业科学院轮台果树资源圃,新疆轮台 841600;2新疆农业大学园艺学院,乌鲁木齐 830052)
0 引 言
【研究意义】自交不亲和性(self-incompatibility,SI)是植物花粉与雌蕊之间的相互识别,防止其近亲繁殖的一种生殖机制。欧洲李(PrunusdomesticaL.)属于蔷薇科果树,该系列优良品种在新疆喀什、阿克苏等地区均有栽培,其中喀什地区栽培面积最大,2021年为4.25×104hm2[1]。法兰西是该系列品种中主栽品种之一,女神、斯泰勒等品种是继法兰西之后的栽培品种。欧洲李系列品种大部分自花不能结实或自交结实率较低[2-3],属于典型的配子体自交不亲和性果树(self-incompatibility,GSI),在生产中需要配置适宜的授粉树才能保证一定的经济效益。GSI由一个具有复等位基因的S位点(S-locus)控制,当花粉与雌蕊中的其中一个S等位基因相同时,花粉无法识别或花粉管生长受到抑制,从而表现为自交不亲和。鉴定欧洲李品种的S基因型并预测授粉品种之间的亲和性,对生产果园合理配置授粉树及杂交育种亲本的选择具有重要意义。【前人研究进展】早期通过田间授粉、IEF-PAGE和SDS-PAGE方法鉴定品种的S基因型。随着S基因PCR鉴定技术的发展,并以田间授粉予以辅助验证,S基因型鉴定已在部分蔷薇科核果类果树中取得了较大进展,如在杏(Prunusarmeniaca)[4-6]、甜樱桃(Prunusavium)[7-8]、扁桃(Prunusdulicis)[9-10]等树种上都成功鉴定出了S-RNase基因,并以鉴定的86份仁用杏S基因型为基础,构建了杂交组合数据库。李S基因相关的报道主要集中在中国李和杏李中,张树军等[11]从15个槜李等品种中鉴定出了9个新的S-RNase基因,分别为S15~S22和S27。朱福荣等[12]结合田间授粉验证,对桐乡槜李及其4个授粉品种进行了S-RNase基因鉴定,结果表明4个授粉品种不适宜作为槜李的授粉树。李芳东等[13-14]先后确定了味王、味厚等7个杏李S-RNase基因型和花粉SFB基因型。李洪果[15]利用RT-PCR技术鉴定了45个李和杏李品种的S基因型,并根据花期、单果重和内在品质等多个性状指标筛选出了66对可供进行授粉杂交的组合。目前,已有近70个李和杏李品种的S基因型被相继鉴定。欧阳丽婷等[16]认为野生欧洲李属于异交为主的混合交配系统,具有一定的自交可能。王斐[2]、冯贝贝等[3]利用田间授粉验证了栽培欧洲李品种属于自交不亲和果树,并通过不同品种间的坐果率确定了授粉树。【本研究切入点】目前国内主要对欧洲李的研究集中在资源调查、引种栽培、部分生物学特性及组织培养等方面[17-20],但关于S基因方面的研究还未见报道。研究鉴定欧洲李品种自交不亲和S-RNase基因型,有助于合理配置授粉品种、高效育种及提高生产效率。【拟解决的关键问题】利用PCR和DNA测序技术鉴定9个欧洲李品种的S基因型,以丰富欧洲李品种S基因型信息,为杂交育种亲本的选择和授粉树的配置提供科学依据。
1 材料与方法
1.1 材 料
9个试验品种采自新疆农业科学院轮台果树资源圃2号种质保存圃(84°22′E、41°78′N,海拔906 m),分别是法兰西、女神、斯泰勒、理查德早生、塔城酸梅、乌孜干1号、总统、Silvia及Decbrowice。该保存圃地势平坦,园相整齐,水肥管理良好。试验树均于2013年定植,株行距为2 m×4 m,树体生长良好。2021年4月下旬采集各品种幼嫩的叶片用硅胶保存备用。
1.2 方 法
1.2.1 叶片DNA提取
参照新型植物基因组DNAsecure试剂盒(天根科技生化有限公司)提取欧洲李叶片DNA,用超微量紫外可见分光光度仪(型号ND5000)在260和280 nm处测定吸收值确定DNA的质量,1.2%琼脂糖凝胶电泳确定抽提DNA的完整性。
1.2.2 引物
选取2对蔷薇科通用引物组合,分别为Pru-C2/PruCE-R、EM-PC2consFD/EM-PC3consRD。引物由宝生物工程(大连)有限公司合成。表1

表1 选取的引物组合及其序列
1.2.3 S基因的特异性PCR扩增
利用2对引物组合对试验品种进行PCR扩增,扩增体系和程序参考Ortega等[23]方法,并在此基础上略作调正,PCR反应为30 μL:DNA模板2 μL,3 μL 10×Buffer,1 μL dNTP(浓度50 ng/mL),1 μL正反向引物(浓度10 mmol/L),1 μLTaqDNA聚合酶,21 μL ddH2O。PCR反应程序:95℃预变性10 min,94℃变性30 s,60℃退火30 s,72℃延伸30 s,40个循环后72℃延伸5 min。
1.2.4 S基因的克隆测序
用琼脂糖凝胶电泳分离扩增产物,回收并纯化目的基因条带,连接至pMD18-T克隆载体,转化XL10-Gold感受态细胞,蓝白斑筛选后挑取单菌落于LB培养基中摇菌,酶切法鉴定阳性菌落,挑取质粒送青岛鹏翔生物科技有限公司测序,3次重复测序。
1.3 数据处理
测序结果用生物信息学软件进行校正分析,将校正后的测序结果在NCBI中Blast搜索比对同源性并分析序列,确定供试品种S基因型。
2 结果与分析
2.1 欧洲李品种 S-RNase基因的PCR扩增
研究表明,9个欧洲李品种均扩增出2条带,检测出13个大小不同的片段,片段长度在308~830 bp,其中引物Pru-C2+PruCE-R扩增出6个大小不同的片段,包括:308 bp(理查德早生等)、309 bp(Silvia等)、473 bp(女神)、653 bp(斯泰勒)、659 bp(乌孜干1号)、662 bp(乌孜干1号),引物EM-PC2consFD+EM-PC3consRD扩增出6个大小不同的片段,包括330 bp(法兰西等)331 bp(理查德早生等)、482 bp(总统)、678 bp(塔城酸梅)、736 bp(Decbrowice)、828 bp(Silvia)、830 bp(法兰西等)。图1,图2

注:M为2 000 bp的Marker,1.理查德早生;2.乌孜干1号;3.Silvia;4.法兰西;5.总统;6.Decbrowice;7.斯泰勒;8.女神;9.塔城酸梅
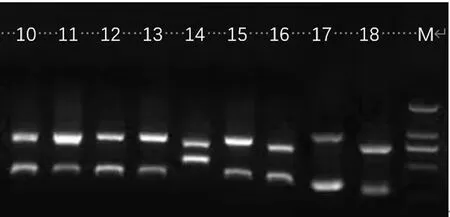
注:M为2 000 bp的Marker,10.理查德早生;11.乌孜干1号;12.Silvia;13.法兰西;14.总统;15.Decbrowice;16.斯泰勒;17.女神;18.塔城酸梅
2.2 欧洲李S-RNase基因型鉴定
研究表明,仅有1条S-RNase基因序列与已登录的欧洲李S-RNase基因序列同源性仅为96%,其它序列均高度同源。
不同引物扩增出的308、331 bp两个大小不同的基因片段与S1-RNase基因同源性为100%,属于同一种S-RNase基因。同一种引物扩增出的序列长度相近的片段也有可能属于同一种基因,如330 bp与331 bp、659 bp与662 bp、828 bp与830 bp。根据13个基因片段在GenBank中的同源性比对结果,确定了7条不同的欧洲李S-RNase等位基因,鉴定出了9个参试品种中的8个欧洲李品种的S-RNase基因型,其中塔城酸梅仅鉴定出具有S1-RNase等位基因。理查德早生和斯泰勒基因型相同为S1S9;法兰西和Silvia基因型相同为S1S5;乌孜干1号、总统、Decbrowice和女神4个品种的S基因型分别为S1S11、S6SSJ、S1SSJ和SSAS11。表2

表2 13个基因片段在GenBank上的比对
2.3 欧洲李S基因频率
研究表明,理查德早生、斯泰勒等7个品种共有1条S1-RNase基因,法兰西和Silvia、理查德早生和斯泰勒分别具有S5和S9基因,总统和Decbrowice、女神和乌孜干1号分别具有SSJ和S11基因,各个S基因出现的频率具有较大差异,其中S1基因出现的频率最高为66.7%;S6、SSA基因出现的频率最低为11.1%。
2.4 供试品种在蔷薇科李亚科S-RNase基因遗传距离
研究表明,从NCBI数据库中选取20条蔷薇科李亚科已公布的S-RNase基因序列:欧洲李、杏、甜樱桃、扁桃、梅(Prunusmume)、黑刺李(Prunusspinosa)、中国李(Prunussalicina)、野樱桃(Prunusvirginiana)、野杏(Prunuswebbii)、樱桃李(Prunuscerasifera)、桂樱(Prunuslaurocerasus)、欧李(Prunushumilis),再与试验获得的7条不同的S-RNase基因序列共同利用MEGA6软件的Neighbor-Joining法构建系统进化树。该发育树聚成3个大类,第一大类主要为试验检测出的5个欧洲李的S-RNase基因和其它李亚科树种的S-RNase基因的聚类;第二大类将野樱桃、黑刺李、中国李和欧洲李S1-RNase基因聚为一类;第三大类为甜樱桃、扁桃、野杏、中国李和试验欧洲李中检测出的S11-RNase基因的聚类。获得的7个S-RNase基因在3个分支上均有分布,其中欧洲李SSA、S6、S9、SSJ和S5的亲缘关系较近。李亚科的杏、中国李、黑刺李等树种的S-RNase基因均不能单独聚为一类,而是彼此交错分布在各个分支中。图3
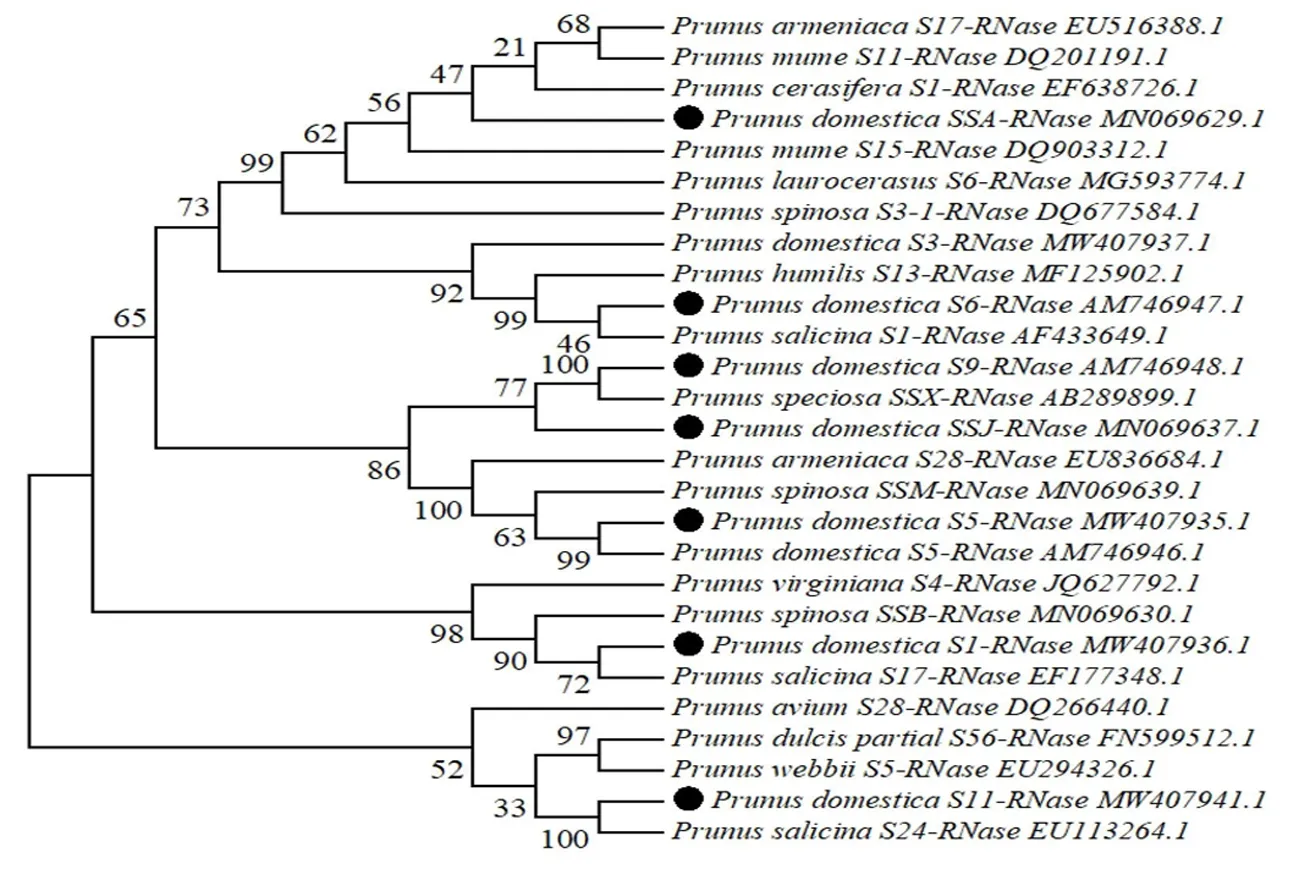
注:●表示研究中获得的基因;SSA:女神;S6:总统;S9:理查德早生,斯泰勒;SSJ:总统,Decbrowice;S5:法兰西,Silvia;S1:理查德早生,斯泰勒,法兰西,Silvia,乌孜干1号,Decbrowice,塔城酸梅;S11:乌孜干1号,女神
3 讨 论
关于S-RNase等位基因扩增时引物的选择,目前大多数选用蔷薇科通用引物直接进行PCR扩增[5-6],也有根据S等位基因的特异序列设计引物再结合蔷薇科通用引物两种方法进行PCR扩增[24],能扩增出理想条带。试验选用了2对蔷薇科通用引物对9个供试欧洲李品种均扩增出了2个条带,共检测出13个大小不同的基因片段。也发现利用单对蔷薇科通用引物进行PCR扩增,并不能检测出所有供试欧洲李品种的S-RNase基因片段,姜新等[5]也发现过此类现象,并且选用了3对蔷薇科通用引物成功鉴定出了供试新疆杏品种的S基因型。
鉴定9个供试欧洲李品种的S基因型发现,塔城酸梅品种的尚未鉴定出S基因型,仅鉴定出单条S1-RNase基因,而另1条S基因与NCBI中同物种相似度最高的S16基因相比,同源性仅为96%,该条S基因可能为新S基因。同时也发现法兰西和Silvia品种的S基因型相同为S1S5,理查德和斯泰勒品种的S基因型也相同为S1S9,通过比较两组疑似品种表型性状发现,各组之间除果实大小和果皮色泽存在差异外,其它形态特征和部分生物学性状相似度较高,很可能是它们的亲缘关系很近或者属于同物异名现象,后续需借助DNA分子标记可进行鉴定。此类现象在柚类资源[25]中也有过报道。梁梅[26]研究发现,S-RNase基因型相同的不同品种不能互作授粉树,试验鉴定出的女神与法兰西、理查德早生、塔城酸梅、总统和Decbrowice品种基因型不同,可以互作授粉树。王斐等[2]对法兰西和女神授粉可以验证S基因型鉴定的结果。法兰西、理查德早生、乌孜干1号和塔城酸梅品种的S基因型中都共有1条S1-RNase基因,理论上它们之间不宜互作授粉树。刘梦培[4]结合田间授粉对具有1条相同S-RNase基因的多个仁用杏品种进行了验证。但也有例外,在个别品种如南杂和北杂两个仁用杏中都共有1条S9-RNase基因,理论上应为杂交半亲和,实际田间授粉表明二者杂交亲和,但正反交的坐果率差异较大,杏品种间存在杂交不亲和现象,并有单方不亲和与相互不亲和之分[27],是否在欧洲李中也有此现象,后续有待进一步研究。
植物S基因的遗传特性对生殖方式是自交还是异交起着决定性作用[28]。研究发现,鉴定出的7条不同的S-RNase基因在供试欧洲李品种中分布频率不均衡,其中S1-RNase等位基因出现的频率最高,可能是该基因在品种间授粉时有较强的选择优势,也有可能是该基因与其它基因相互连锁,可产生有利的经济性状,从而表现出有作为亲本的选择优势[29-30]。总统和女神两个栽培品种中的S6-RNase和SSA-RNase基因出现频率最低,可能是因为供试欧洲李品种克隆出的S-RNase基因群体数量较少,后续需要克隆更多的S-RNase基因可进一步验证。
在构建的系统进化树中,欧洲李、中国李、杏等李亚科树种的S-RNase基因并没有单独聚成1个亚类,而是彼此交错、随机分布在李亚科各树种中,树种之间亲缘关系较近。蔷薇科S-RNase基因的种间同源性往往大于种内同源性[6,31-32]。研究的系统进化分析结果也验证了此观点,从而说明李亚科植物S基因的分化早于各个种形成之前,而S基因种内的进化发生在各自单独的系统中。
4 结 论
供试欧洲李品种的S基因型是由2种不同的S等位基因组成,检测出了7个不同的S-RNase等位基因,确定了9个供试样品中的8个欧洲李品种的S基因型,其中理查德早生和斯泰勒的S基因型相同为S1S9,法兰西和Silvia的S基因型相同为S1S5,乌孜干1号、总统、Decbrowice 和女神的S基因型分别为S1S11、S6SSJ、S1SSJ和SSAS11,塔城酸梅仅确定具有1条S1-RNase基因,未能鉴定出其S基因型。S基因频率分析显示S1-RNase基因频率最高,S6-RNase和SSA-RNase基因频率最低。李亚科植物S基因的分化早于各个种形成之前,而S基因种内的进化发生在各自单独的系统中。鉴定欧洲李品种自交不亲和S-RNase基因型,可以丰富欧洲李品种S基因型信息。
