ANG及其突变体对膀胱癌血管生成的影响及分子机制研究
2023-06-19刘玉林黄晓楠陈俊霞
刘玉林,黄晓楠,向 玉,陈俊霞
1.四川省遂宁市中心医院检验科,四川遂宁 629000;2.重庆医科大学细胞生物学与遗传学教研室,重庆 400016
膀胱癌是泌尿系统常见的恶性肿瘤之一,其发病率居我国泌尿系统肿瘤第一位[1]。相关数据显示,2020年全球新增膀胱癌确诊病例约5.73万例,占全部恶性肿瘤新发病例的3.0%,死亡约2.13万例,占全部恶性肿瘤死亡病例的2.1%[2]。血管生成素(ANG)是非常重要的血管生成因子,由147个氨基酸组成,包含一段信号肽序列,具有弱RNA酶活性[3]。ANG在多种实体瘤中表达升高,研究表明其与血管生成及肿瘤发生发展密切相关[4],新生血管形成可促进肿瘤的发生发展。ANG可以通过调节黏附因子表达[5]及免疫细胞[6-7]参与免疫反应,也可通过调节肿瘤血管变化[8]间接参与免疫反应。本文主要探索ANG及其突变体对膀胱癌血管生成的影响和作用机制,从血管生成角度探索膀胱癌的发生发展。
1 材料与方法
1.1材料 膀胱癌BIU87细胞(南京凯基生物有限公司);RPMI 1640培养基(美国Gibico公司);脂质体Lipofectamine 2000(美国Invitrogen公司);Endothelial Tube Formation Assay(体外血管,美国Cell Biolabs公司)、基质胶(美国BD公司);兔抗人磷脂酰肌醇3激酶(PI3K)、磷酸酯酶与张力蛋白同源物(PTEN)、磷酸化PI3K(p-PI3K)、蛋白激酶B(AKT)、磷酸化AKT(p-AKT)、磷酸化PTEN(p-PTEN)、糖原合成酶(GSK3)、磷酸化GSK3(p-GSK3)及二抗(美国Immunoway公司);ANG单克隆抗体(美国Abcam公司);核糖核酸酶抑制因子(RI)多克隆抗体(重庆医科大学细胞生物学与遗传学教研室保存);G418(中国碧云天生物技术有限公司);Dual-Glo®Luciferase Assay试剂盒和PRL-TK质粒(美国Promega公司);白皮受精鸡蛋(重庆市种禽育种公司)。
1.2实验方法
1.2.1构建突变体质粒
1.2.1.1pCMV-3×flag-ANGH13R和pCMV-3×flag-ANGH114R突变体合成 pCMV-3×flag-ANGH13R引物:上游引物为5′-CACACTTCCTGACCCAGCGCTATGATGCCAAACCACAG-3′,下游引物为5′-CTGTGGTTTGGCATCATAGCGCTGGGT CAGGAAGTGTG-3′;pCMV-3×flag-ANGH114R引物:上游引物为5′-GAAAATGGCTTACCTGTCCGCTTGGATCAGTCAATTTTC-3′,下游引物为5′-GAAAATTGACTGATCCAAGCGGACAGGTAAG CCATTTTC-3′。
1.2.1.2突变反应体系 模板DNA 2.5 μL,突变体引物2.0 μL,dNTPs 4.0 μL,Pyrobest聚合酶0.4 μL,pyrobest buffer 5.0 μL,ddH2O 36 μL,总体积50 μL。突变条件:94 ℃,30 s;68 ℃,30 s;72 ℃,10 min;12个循环。1%琼脂糖凝胶电泳鉴定突变产物。
1.2.1.3质粒转化 感受态菌中加入突变反应产物,4 ℃静置30 min。42 ℃孵育2 min,4 ℃刺激2 min。加入900 μL LB培养基(不含抗菌药物)混匀,37 ℃恒温摇床振荡培养1 h。取100 μL转化后的混合液接种到LB平板培养皿中(含抗菌药物)。37 ℃孵箱中正放培养30 min后,倒置培养18 h。挑选单克隆接种于LB培养基中(含抗菌药物),37 ℃,300 r/min振荡培养18 h。
1.2.1.4pCMV-3×flag-ANGH13R和pCMV-3×flag-ANGH114R酶切体系 Hind Ⅲ 2.5 μL,BamHⅠ2.5 μL,10×K Buffer 5 μL,pCMV-3×flag-ANG突变体质粒25 μL,ddH2O 15 μL,总体积50 μL。37 ℃恒温金属浴反应5 h,1%琼脂糖凝胶电泳鉴定,并送生工生物工程(上海)股份有限公司测序。
1.2.2建立稳定转染细胞系 实验组(pEGFP-C1-ANGH13R、pEGFP-C1-ANGH114R、pEGFP-C1-ANG)和对照组(pEGFP-C1)质粒分别用脂质体Lipofectamine 2000转染BIU87细胞。24 h后移植到6孔板,细胞培养箱中继续培养,48 h后换10%血清培养基(含600 μg/mL G418)培养。两周后G418浓度减半,用300 μg/mL G418继续培养两周,最后将幸存下来的细胞转移到96孔板中扩大培养,稳定转染细胞系构建成功。
1.2.3蛋白质印迹法(Western blot)检测PI3K/AKT/哺乳动物雷帕霉素靶蛋白(mTOR)通路靶蛋白 提取细胞总蛋白,使用BCA蛋白浓度试剂盒测定蛋白浓度。非磷酸化蛋白AKT、PI3K、PTEN、GSK3(α/β)、mTOR和磷酸化蛋白p-GSK3(α/β)、p-AKT、p-PTEN、p-PI3K、p-mTOR抗体稀释比例均为1∶500。
1.2.4鸡胚尿囊绒毛膜(CAM)血管生成实验 白皮受精鸡蛋清洗消毒,分为实验组和对照组,每组随机10只鸡蛋。37.5 ℃、湿度60%,孵育5 d,75%乙醇消毒气室端蛋壳,用尖嘴镊形成0.5 cm×0.5 cm小孔,形成假气室。75%乙醇消毒,滴2~3滴生理盐水将气室膜和CAM分离开,去除气室膜,暴露CAM。用基质胶将各组细胞制成悬液,分别加入CAM,命名为实验组(BIU87-ANGH114R、BIU87-ANGH13R、BIU87-ANG)和对照组(BIU87-C1)。继续孵育72 h。小孔中加入甲醇和丙醇等体积混合液3 mL,室温固定15 min,将固定后的CAM剪下,风干并拍照。
1.2.5小管形成实验 取预冷的96孔板,每孔加入解冻的ECM胶50 μL,37 ℃孵育30 min,使ECM溶液形成凝胶。每孔加入150 μL细胞悬浮液(1.5~3.0)×104个/毫升到固化的ECM凝胶上,37 ℃孵育10 h。高倍视野下用光学显微镜检查内皮管。
1.2.6过表达RI对ANG及其突变体促核糖体RNA(rRNA)转录活性的影响
1.2.6.1构建荧光素酶报告基因载体 以基因组DNA为模板,扩增rRNA启动子序列,上游引物:5′-CGGGGTACCGCGGTCCCTCTGCCGCGA-3′;下游引物:5′-CCCAAGCTTAGCGCGACCTCTCGGGC C-3′。PCR条件:94 ℃,2 min;94 ℃,30 s,63 ℃,30 s,72 ℃,100 s,30个循环;72 ℃延伸5 min。KpnⅠ和HindⅢ双酶切鉴定。
1.2.6.2双荧光素酶报告基因系统检测rRNA转录活性 96孔板中加入与孔中培养基相等体积的Dual-Glo®试剂,等待10 min,在发光仪中测量萤火虫荧光素酶的发光;向每个孔中加入与原培养基体积相等的Dual-Glo®Stop&Glo®试剂并混合,等待10 min,然后检测海肾荧光素酶发光;海肾荧光素酶发光应按与萤火虫荧光素酶发光相同的板序读取。计算实验报告发光与对照报告发光的比值。
1.3统计学处理 采用SPSS22.0软件进行数据处理和分析。多组间比较采用方差分析,组内比较采用LSD-t检验,以P<0.05为差异有统计学意义。
2 结 果
2.1ANG突变质粒(pCMV-3×flag-ANGH13R、pCMV-3×flag-ANGH114R)构建成功 琼脂糖凝胶电泳结果显示,pCMV-3×flag质粒大小约为6 400 bp,ANG突变体大小约为500 bp,与PCR结果吻合(图1)。测序结果分析也表明,在特定位置发生突变,突变体构建成功(图2)。

注:1为Marker;2为pCMV-3×flag-ANGH13R;3为pCMV-3×flag-ANGH114R;4为pCMV-3×flag-ANG;5为pCMV-3×flag;6为Marker。图1 pCMV-3×flag-ANGH13R、pCMV-3×flag-ANGH114R突变体质粒的酶切鉴定
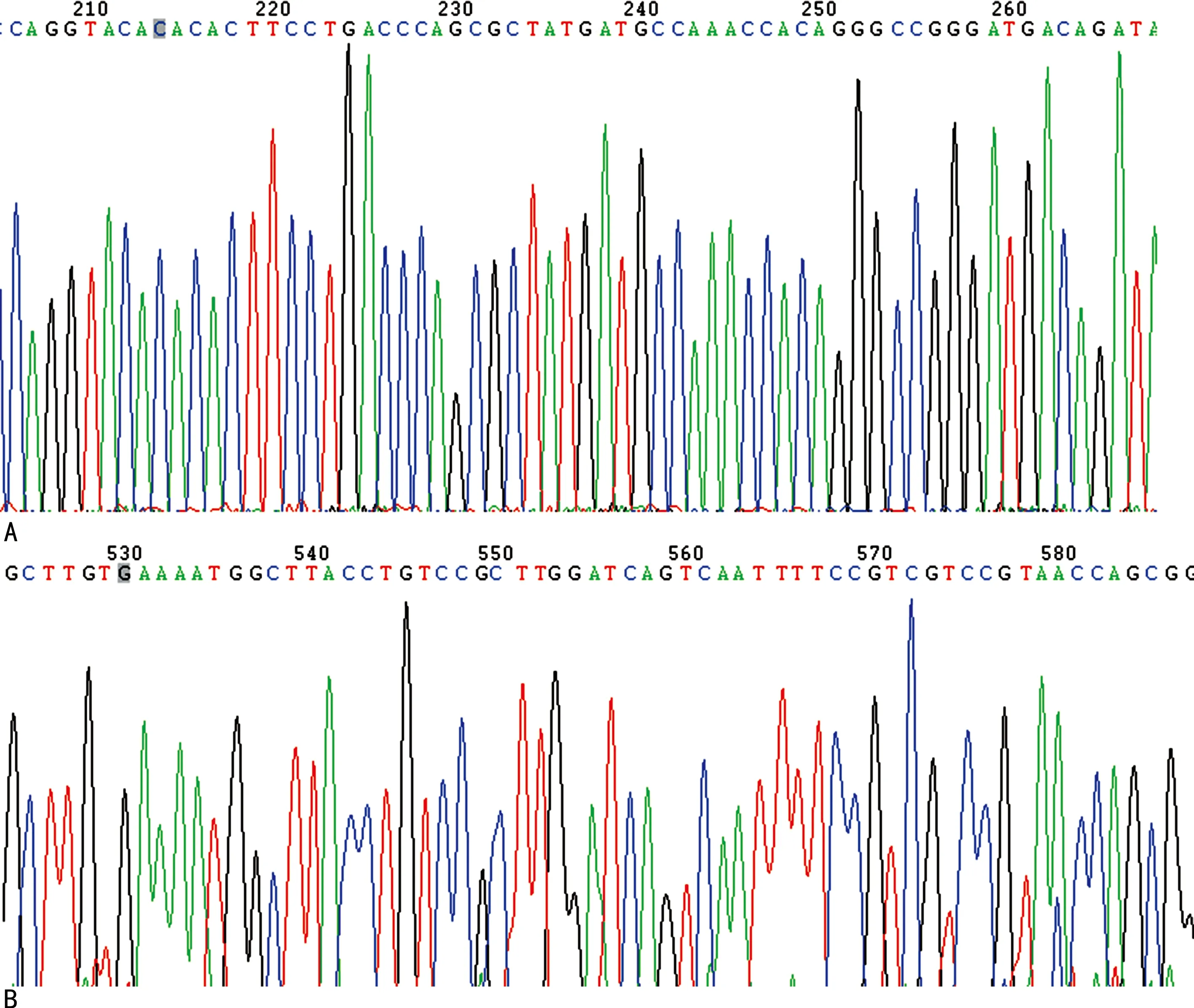
注:A为pCMV-3×flag-ANGH13R测序鉴定图;B为pCMV-3×flag-ANGH114R测序鉴定图。图2 pCMV-3×flag-ANGH13R、pCMV-3×flag-ANGH114R测序鉴定
2.2稳定转染ANG及其突变体的膀胱癌BIU87细胞系构建成功 各组稳定转染细胞系都属于pEGFP-C1 系列带有绿色荧光,可直接用荧光显微镜观察细胞系表达情况,表明各组稳定转染细胞系构建成功。见图3。
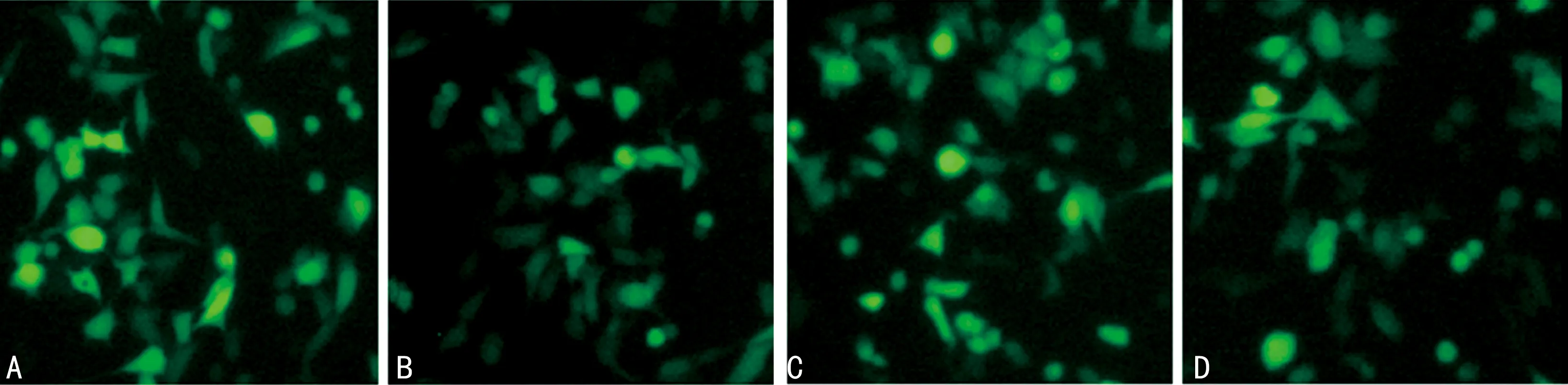
注:A为BIU87-ANGH13R;B为BIU87-ANGH114R;C为BIU87-ANG;D为BIU87-C1。图3 稳定转染ANG及其突变体的膀胱癌BIU87细胞系
2.3Western blot检测信号通路中的靶蛋白 结果显示,与对照组比较,实验组p-PI3K、p-AKT、p-GSK3(α/β)、p-mTOR、p-PTEN的表达增加,差异有统计学意义(P<0.05)。见图4。
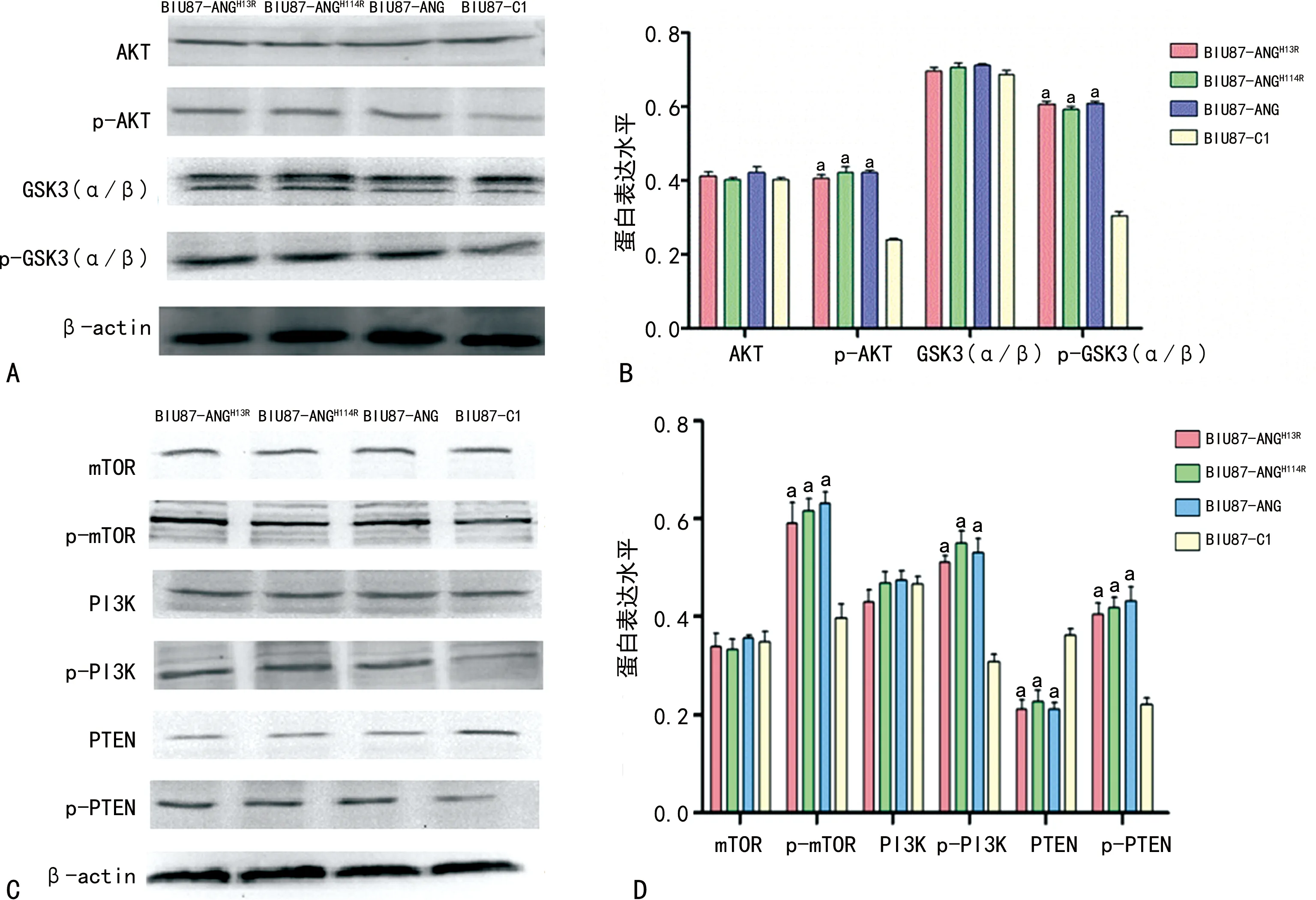
注:A、C为各蛋白的Western blot结果;B、D为各蛋白的统计分析结果;与BIU87-C1比较,aP<0.05。图4 过表达RI调控ANG促PI3K/AKT/mTOR信号通路磷酸化水平
2.4ANG及突变体促进CAM血管生成和小管形成 CAM血管生成实验和小管形成实验结果均显示,实验组(BIU87-ANGH13R、BIU87-ANGH114R、BIU87-ANG)CAM血管生成及小管形成均明显多于对照组(BIU87-C1),表明实验组ANG及其突变体可促进肿瘤血管生成。见图5。
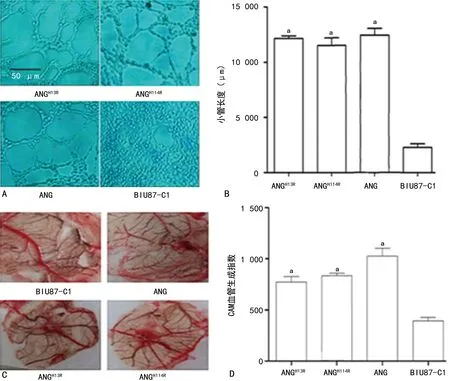
注:A为实验组和对照组小管形成实验结果;B为实验组和对照组小管形成实验统计分析结果;C为实验组和对照组CAM血管形成实验结果;D为实验组和对照组CAM血管形成实验统计分析结果;与BIU87-C1比较,aP<0.05。图5 ANG及其突变体对膀胱癌肿瘤小管形成及CAM血管生成的影响
2.5RI抑制ANG及其突变体促核糖体RNA的转录活性 运用双荧光素酶报告基因检测系统对rRNA转录活性检测结果表明,RI抑制ANG及其突变体促核糖体RNA的转录活性,差异有统计学意义(P<0.05)。见图6。
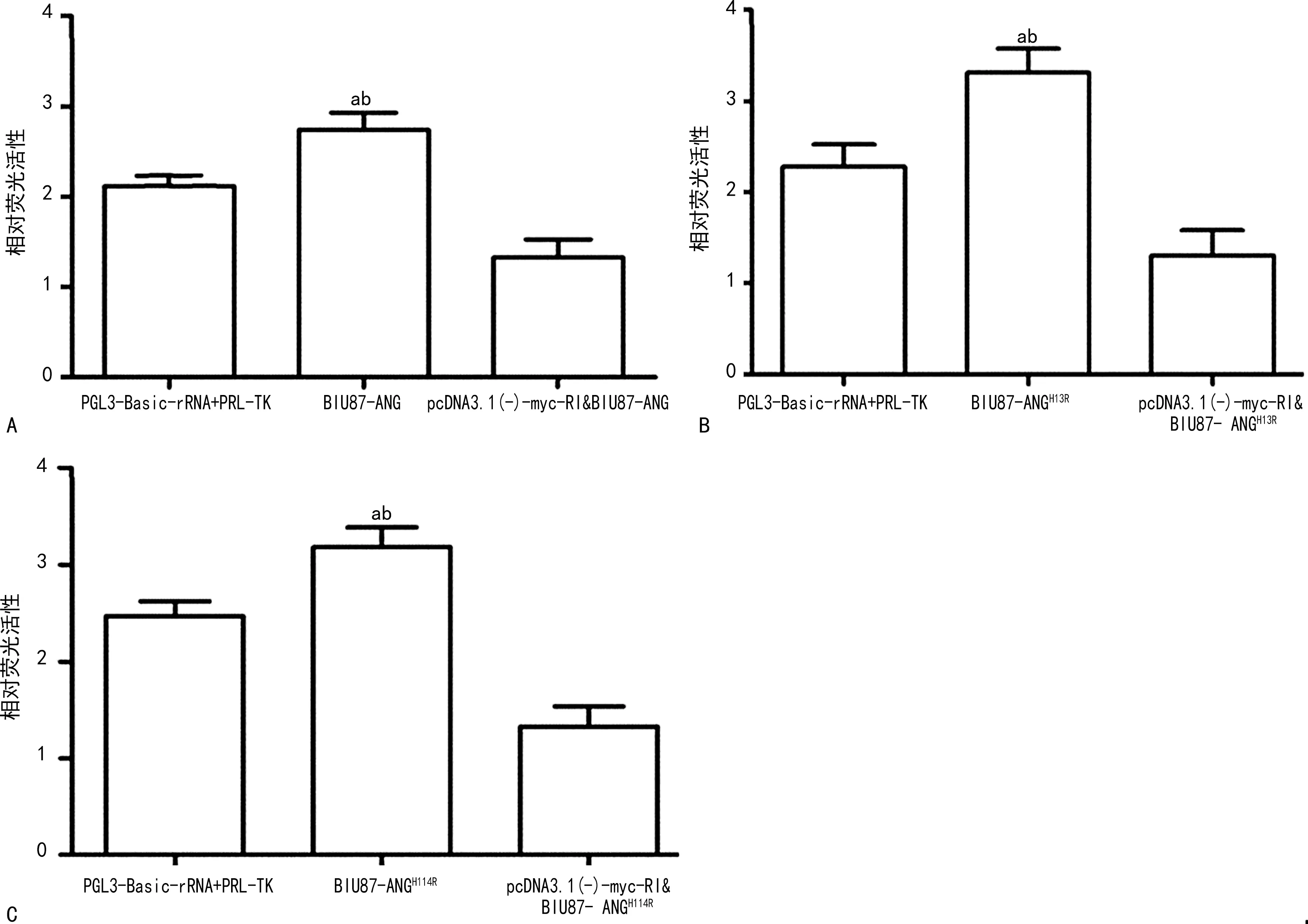
注:A为BIU87-ANG组双荧光素酶相对荧光活性;B为BIU87-ANGH13R组双荧光素酶相对荧光活性;C为BIU87-ANGH114R组双荧光素酶相对荧光活性;与PGL3-Basic-rRNA+PRL-TK比较,aP<0.05;与pcDNA3.1(-)-myc-RI&BIU87-ANG比较,bP<0.05。图6 RI抑制ANG及其突变体促核糖体RNA的转录活性
3 讨 论
ANG是一种重要的促进血管生成的作用因子,在淋巴管和血管生长成熟中起关键作用,由纤维蛋白原样结构、蛋白信号肽、螺旋结构域三部分组成。目前发现4个亚型[9],分别是ANG-1、ANG-2、ANG-3、ANG-4。ANG-1是 Tie2 的激动剂,与Tie2结合后能激活并诱导其磷酸化,促进血管的内皮细胞形成管状结构,增加结构稳定性;吸引血管周围细胞支持,包围内皮细胞,促进血管重新塑造,以维持血管完整性[10]。ANG-2约有60%与ANG-1相同,机体胚胎发育时其主要在肝血管壁和大血管平滑肌中表达,而处于成体时其主要在胎盘、子宫和卵巢中表达,通过抑制ANG-1活化的程度,开始调节内分泌环路,维持血管的生长及退化的动态平衡,在缺氧情况下可促进其表达[11]。目前,关于ANG-3、ANG-4的研究较少。在ANG的123个残基蛋白质中包含许多重要的生物活性位点,比如核转位序列、核受体结合位点、RNase活性中心等,此类位点可能与ANG的未知生物功能相关[12]。本研究针对核转位序列将ANG基因13位和114位的氨基酸由组氨酸突变为精氨酸,目的是初步探索ANG的生物特性,为进一步了解ANG促进膀胱癌肿瘤血管生成和肿瘤细胞增殖的研究做准备。
研究表明,血管生成及新血管的形成对实体瘤的生长转移起非常重要的作用,新生成的血管可为实体肿瘤的生长和转移提供丰富的氧气和营养物质,为肿瘤细胞的转移和免疫细胞的渗透提供通道,并且能带走肿瘤细胞产生的废物[13]。在没有新生血管形成时,实体肿瘤长到1~2 mm3时,肿瘤减小消退,甚至消失。在肿瘤的早期阶段,新生血管形成较少且缓慢,肿瘤处于休眠状态,当促血管生成因子占据优势时血管形成活跃,诱导新生血管形成,是肿瘤血管化的启动标志[14]。外源性试验抑制血管的生成可以明显地抑制肿瘤的生长,甚至可以使肿瘤消退,所以新生血管对于肿瘤的生长和转移发挥巨大作用。所有的实体肿瘤生长和转移都依赖于新生血管的形成,这个特征为实体肿瘤的诊断和治疗提供了一个新的方向。
本研究构建了pCMV-3×flag-ANGH13R、pCMV-3×flag-ANGH114R两个突变体质粒用于初步探索ANG的生物学功能,研究结果表明,ANG及上述两个突变体能促进小管的形成,在CAM血管形成实验中促进血管生成,实验组(BIU87-ANGH13R、BIU87-ANGH114R、BIU87-ANG)CAM血管生成及小管形成均明显多于对照组(BIU87-C1),表明ANG及其突变体可促进肿瘤血管生成。相关报道表明,ANG可通过PI3K/AKT/mTOR信号通路刺激rRNA转录和核糖体生物合成,其双重作用为肿瘤的治疗提供了方向[15]。PI3K/AKT/mTOR是一个经典的信号通路,影响细胞凋亡和肿瘤细胞生存,调节细胞增殖及分化[16]。ANG和PI3K/AKT/mTOR信号通路中的AKT交互应答,ANG激活AKT,激活的AKT促进ANG的核转位[16]。mTOR作为PI3K/AKT/mTOR信号通路下游的一个效应器,AKT能直接激活mTOR的磷酸化反应。ANG诱导AKT的磷酸化,可促进伤口愈合和CAM血管的生成。ANG不仅能促进PI3K/AKT磷酸化并激活AKT传导通路,还能使GSK-3β、mTOR蛋白磷酸化,磷酸化可抑制其生物学作用。ANG可通过PI3K/AKT/mTOR信号通路参与肿瘤血管的发生发展。前期研究证实,RI与ANG相互作用抑制BALB/C裸鼠人膀胱癌移植瘤的生长转移,同时抑制PI3K/AKT/mTOR信号通路中磷酸化蛋白的水平[17]。也有研究显示,Ang/Tie2 信号通路与结直肠癌[18]、黑色素瘤[19]、乳腺癌[20]等肿瘤的发生发展存在密切联系,这也为下一步研究拓展了方向。
本研究寻找到ANG中H13R和H114R两个可能的突变位点,它们通过PI3K/AKT/mTOR通路影响其促血管生成特性,为探索ANG在膀胱癌中的分子机制提供理论依据。目前研究结果表明,本研究ANG突变体基本达到预期的效果,笔者将在后续实验中继续探索ANG的生物学功能。
