彩色花椰菜叶绿体全基因组特征分析
2023-06-19朱惠霞陶兴林刘明霞
朱惠霞, 陶兴林, 刘明霞
(甘肃省农业科学院蔬菜研究所, 兰州 730070)
花椰菜属十字花科芸薹属甘蓝种的变种,原产于地中海沿岸,由野生甘蓝演化而来,花球颜色以白色为主,在长期的进化过程中逐渐出现了紫色、橙色、淡绿色等,统称为彩色花椰菜。彩色花椰菜中含有一些特殊的物质,如紫色花椰菜中的花青素、橙色花椰菜中的类胡萝卜素、绿色花椰菜中的叶绿素等,具有保护视力、缓解过敏、保护心血管、抗衰老、抗炎和抗癌作用,越来越受到人们的关注[1-5]。在十字花科作物中,通过对油菜、芥菜、萝卜、芝麻菜、芜菁、海甘蓝、甘蓝、西兰花等叶绿体全基因组的研究[6-16],揭示了部分十字花科叶绿体基因组结构特征,并利用叶绿体基因组在大小、种类以及排列顺序的差异[17-19],结合系统发育分析探讨不同十字花科作物属种间的关系。本研究分析白色、紫色、橙色、淡绿色花椰菜的叶绿体全基因组特征,并进行系统发育分析,以期阐明花椰菜与十字花科物种之间亲缘关系,为彩色花椰菜遗传关系研究提供基本信息。
1 材料与方法
1.1 试验材料
试验材料分别为白色、紫色、橙色、淡绿色花椰菜(BrassicaoleraceaL. var.botrytis),于2019 年3月种植于甘肃省农业科学院蔬菜研究所试验地(36.0999°N,103.6900°E),试验品种均由甘肃省农业科学院蔬菜研究所提供,4~5片叶片时采集材料嫩叶,干冰保存,送样进行测试分析。凭证标本存放于甘肃省农业科学院蔬菜研究所实验室超低温冰箱。
1.2 试验方法
1.2.1DNA的提取与测序
提取花椰菜样品DNA,检测DNA质量做片段化处理,纯化并进行末端修复,再选择片段扩增,建成测序文库。质检合格后用Illumina HiSeq平台进行测序。
1.2.2叶绿体基因组组装与注释
对Raw Data软件进行数据过滤,按参考物种的叶绿体基因组序列进行组装和基因结构注释,得到完整的花椰菜叶绿体基因组序列。利用BLAST软件对比NCBI数据库(National Center for Biotechnology Information,美国国家生物技术信息中心)叶绿体基因组cds序列,手工校正后获得最终注释结果,使用hmmer软件比对NCBI数据库里叶绿体基因组rRNA序列,使用aragorn软件进行tRNA的预测,分别获得rRNA和tRNA的注释信息,利用 OGDRAW 在线软件绘制花椰菜叶绿体基因组图谱。
1.2.3SSR位点与密码子偏好性分析
使用MISA (MIcroSAtellite identification tool)软件进行叶绿体基因组SSR分析,利用Codon W 1.4.2 软件检测花椰菜叶绿体基因组中大于200 bp 蛋白编码基因的密码子,然后分析相对密码子的使用频率(RSCU)。
1.2.4系统发育分析
从NCBI数据库下载部分十字花科类植物的叶绿体全基因组序列,以紫罗兰作为外类群,与4种颜色的花椰菜叶绿体基因组进行比对,RAxML软件,选用GTRGAMMA模型,进行进化树构建。
2 结果与分析
2.1 叶绿体基因组特征
获得4种颜色花椰菜叶绿体基因组信息并上传NCBI数据库,登录号分别为:橙色花椰菜:MT 499334;白色花椰菜:MT 499335;淡绿色花椰菜:MT 499336;紫色花椰菜:MT 499337。花椰菜叶绿体基因组共由4个区域组成环状结构,包括1个88 136~88 138 bp的长单拷贝(LSC)、1个17 834~17 835 bp的短单拷贝区(SSC)以及2个26 196~26 197 bp的反向重复 (IRa/IRb),淡绿色花椰菜叶绿体基因组全长为153 366 bp,其余3种全长均为153 364 bp,平均GC含量均为36.36%,LSC区GC含量34.15%,SSC区GC含量29.10%,IR区GC含量42.35%(表1)。4种颜色花椰菜均注释出132个基因,其中,8个为rRNA基因、37个为tRNA基因、87个为蛋白编码基因,基因数量与排列顺序完全相同,在IR区重复2次的有19个基因,包括4个rRNA基因、7个tRNA基因和8个蛋白编码基因(表2)。
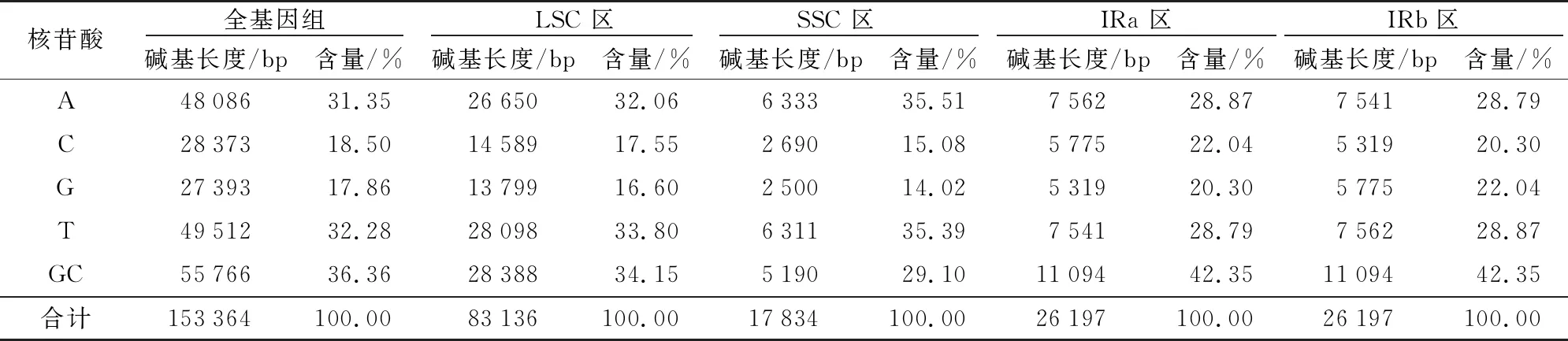
表1 花椰菜叶绿体基因组长度和核苷酸含量

表2 花椰菜叶绿体基因组注释基因
花椰菜叶绿体组中有22个基因具有内含子(表3),其中有12个分布在LSC区,9个分布在IR区,1个分布在SSR区,其中,ycf3、rps12、clpP、rps12分别含有3个外显子,ycf3和clpP含有2个内含子。分析外显子和内含子的长度发现,ndhB的外显子Ⅰ和外显子Ⅱ最长,分别为777 bp和762 bp,petB的外显子Ⅰ最短,仅为6 bp,trnK-UUU内含子最长为2 557 bp,trnL-UAA内含子最短为311 bp。

图1 花椰菜叶绿体基因图谱
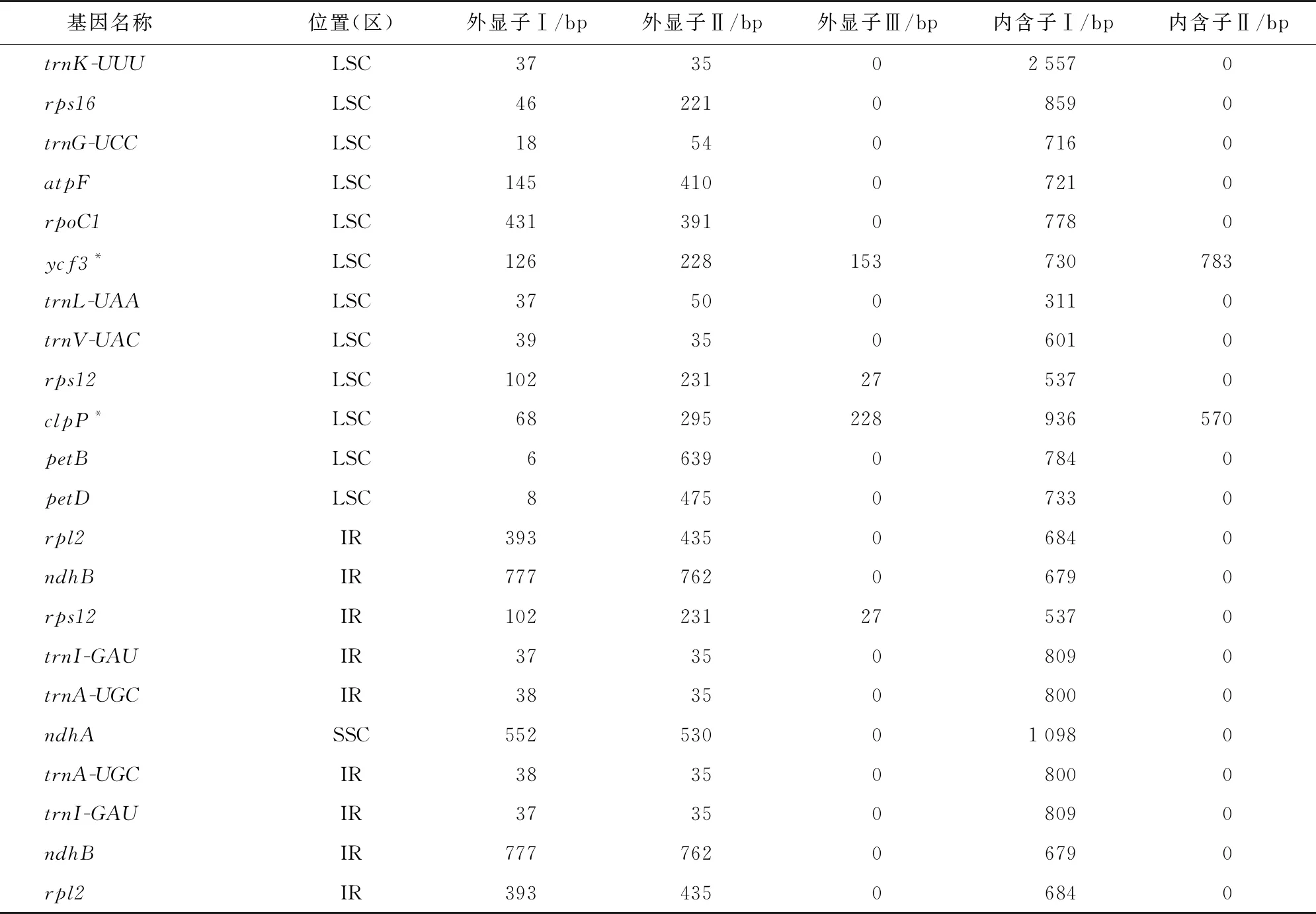
表3 花椰菜叶绿体基因组中具有内含子基因长度
2.2 密码子偏好性分析
在花椰菜叶绿体基因组的67个编码序列中(表4),有32个密码子的RSCU值大于1.00,为高频密码子,其中,以A结尾的13个,以U结尾的16个,其余3个以G结尾,以A或U结尾的同义密码子占90.6%;34个RSCU值小于1.00的密码子中,以G结尾的有13个,以C结尾的有17个,以A结尾的有4个,以G或C结尾的密码子占88.2%;有1个密码子RSCU值等于1。RSCU值最大和最小的密码子均编码甲硫氨酸(Met),其中密码子AUG的RSCU值最大,为3.97,密码子AUC的RSCU值最小,仅为0.01。亮氨酸(Leu)为编码率最高的氨基酸(图2),由CUA、CUC、CUG、CUU、UUA、UUG编码,共有2 828个密码子参与编码。半胱氨酸(Cys)为编码率最低的氨基酸,由UGC、UGU编码,仅有325个密码子参与编码。
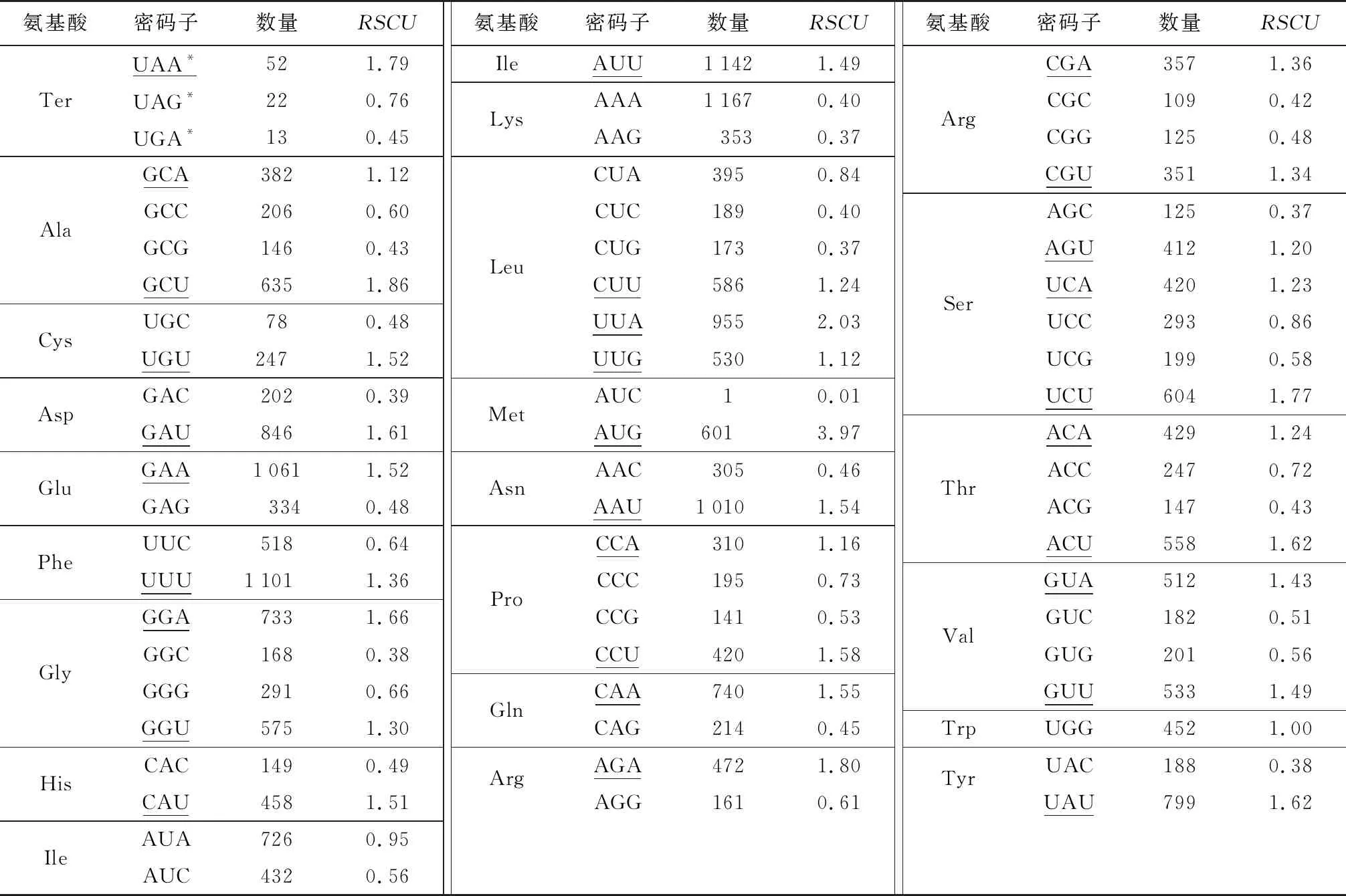
表4 花椰菜同义密码子(RSCU)使用情况分析
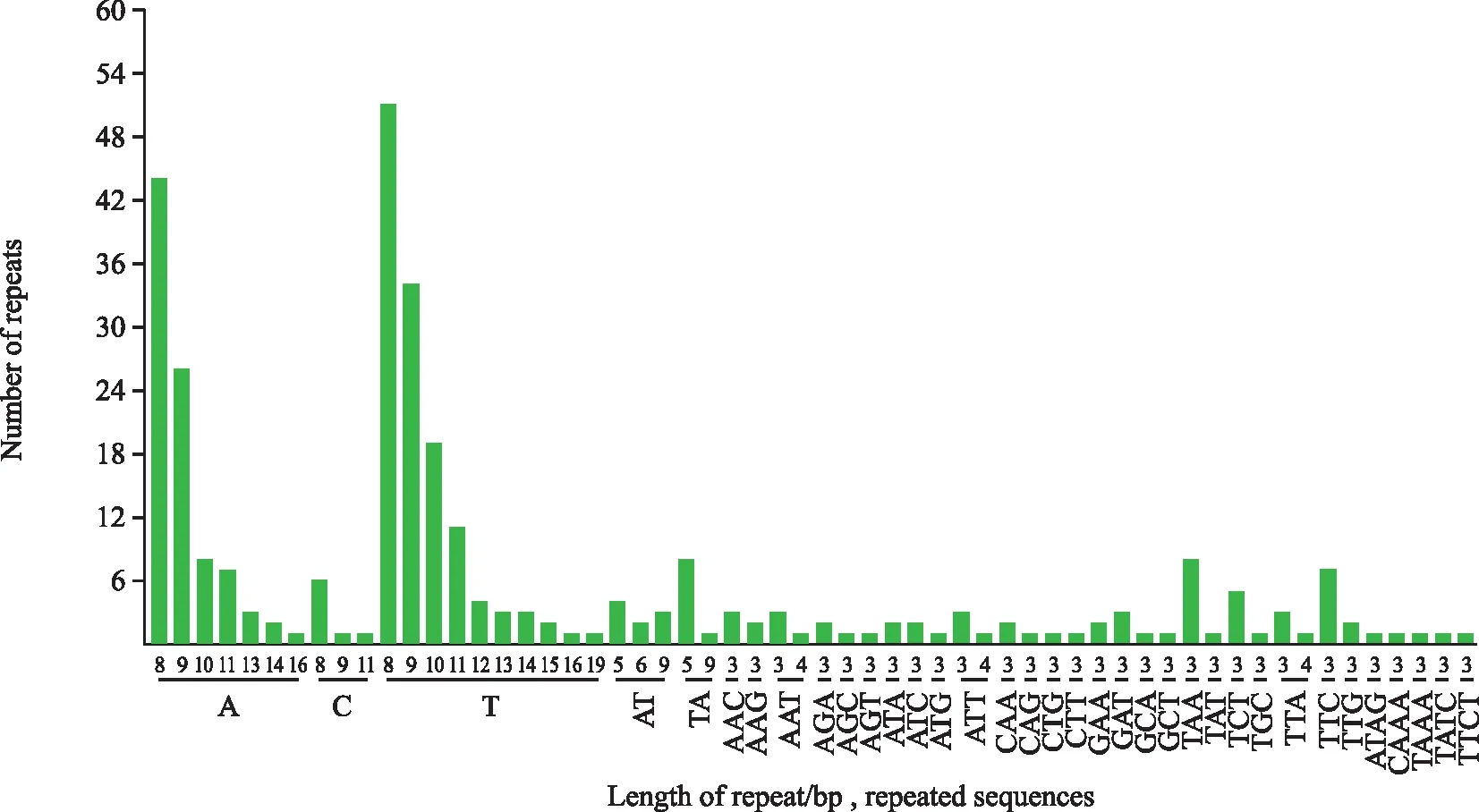
图2 花椰菜叶绿体基因组SSR重复类型与次数
2.3 SSR分析
花椰菜叶绿体基因组共检测到290个SSR位点,其中LSC区188个,占64.8%;SSC区60个,占20.7%;IR区42个,占14.5%。单核苷酸重复有A、C和T共3种类型,占总碱基数的72.84%,在单碱基中,A/T类型最多,占96.49%,重复8次和9次的碱基最多;二核苷酸重复有AT、TA共2种类型,重复次数集中在5~7次;三核苷酸重复有AAC、AAG、AAT、AGA、AGC等25种类型,重复数大多为3次,仅有AAT、ATT、TTA重复了4次。四核苷酸重复有ATAG、CAAA、TAAA、GATT、TATC和TTCT共5种类型,重复次数均为3次(图2)。
2.4 IR边界分析
IR区扩张和收缩分析(图3)表明,4种颜色花椰菜的LSC/IRb边界均位于rps19,且rps19有116~118 bp位于LSC区,112~115 bp位于IRb区; SSC/IRb边界位于ycf1和ndhF,且ndhF向IRb区扩张了34~38 bp,ycf1向SSC区扩张了1~5 bp;但在SSC/IRa边界,白色、橘色、浅绿色花椰菜边界位于ycf1,有1 030 bp位于IRa区,4 358 bp位于SSC区,与其中9种植物相似,但紫色花椰菜的ycf1在边界处被分为两段,位于SSC区4 248 bp的ycf1向SSC区收缩了111 bp,位于LSC区1 030 bp的ycf1向SSC区扩张了14 bp,且缺失了trnN;在IRa/LSC边界,4种花椰菜均为rpl2向LSC区收缩167 bp,trnH向IRa区收缩3 bp。

图3 花椰菜叶绿体基因组 IR/SC 边界分析
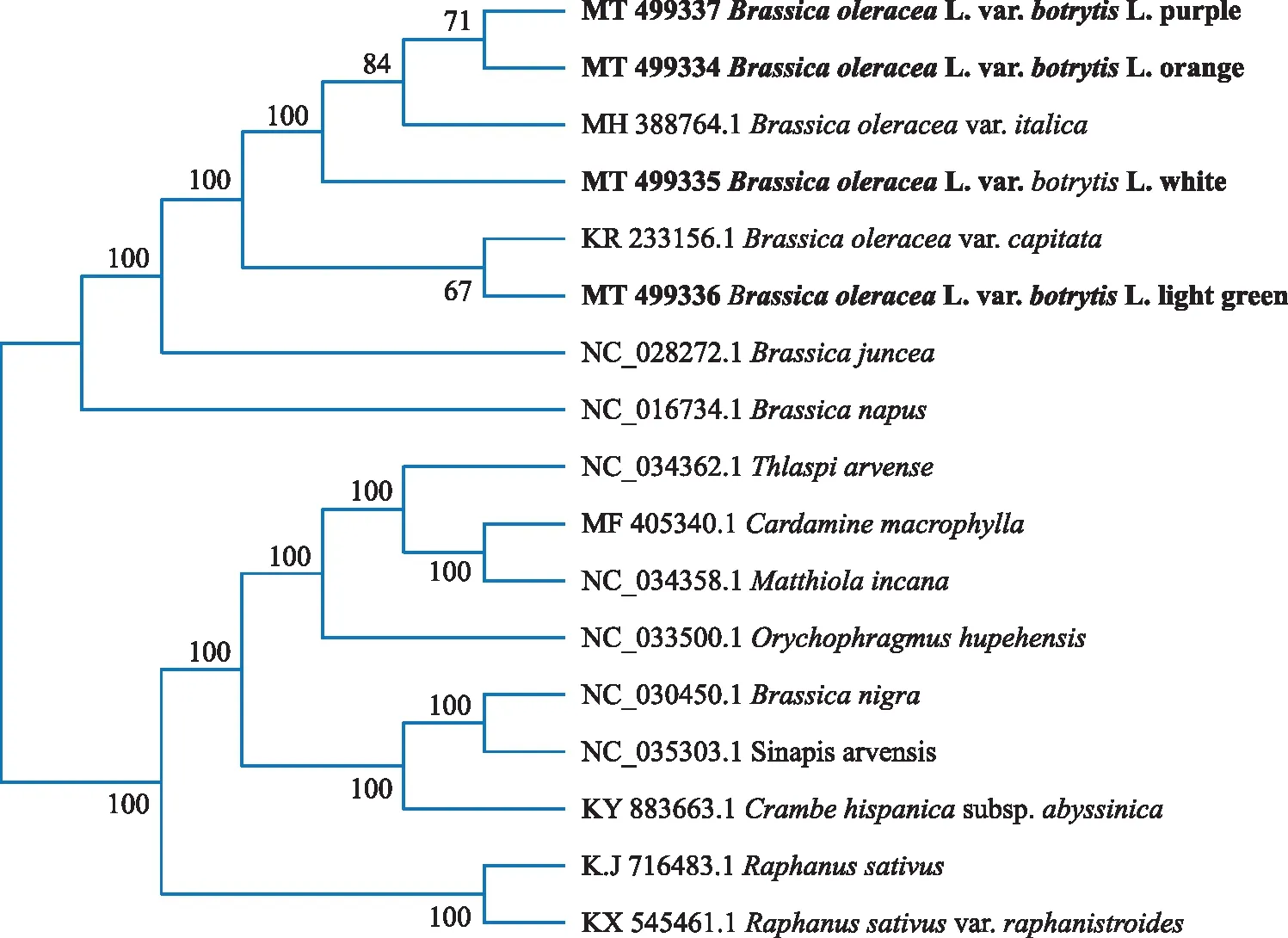
图4 基于叶绿体基因组序列构建的系统发育树
2.5 系统发育分析
将4个花椰菜和十字花科13个物种的叶绿体全基因组序列进行比对后构建系统进化树。结果(图4)显示,4种不同颜色花椰菜中,亲缘关系最近的是紫色与橙色,它们聚为一支后与甘蓝(MH 388764.1)聚在一起,白色花椰菜独立为一支,而淡绿色花椰菜与甘蓝(KR 233156.1)亲缘关系最近,它们聚为一支,之后才与花椰菜和甘蓝聚在一起,最后与独立分支的欧洲油菜、芥菜聚为一个大类,成为十字花科芸薹属的分支,而芸薹属的黑芥和野芥则和其他属的聚在一起。
3 讨 论
芸薹属甘蓝种的叶绿体基因组长度几乎相同,4种颜色的花椰菜叶绿体基因组长度基本一致,淡绿色花椰菜叶绿体基因组全长为153 366 bp,与甘蓝(Brassicaoleracea)相同,白色、紫色、橘色花椰菜全长为153 364 bp,与青花菜(BrassicaoleraceaL. var.italica)相同,总GC含量均为36.36%[19]。芸薹属的芥菜(BrassicajunceaL.)和欧洲油菜(BrassicaNapusL.)的叶绿体基因组全长分别为153 483 bp和152 860 bp[15-20],同一属内的叶绿体基因长度差别最大为623 bp,且受LSC长度的影响较大,所有芸薹属植物具有几乎相同的GC含量,均为36%左右,注释基因111~113种,说明芸薹属植物的叶绿体基因组高度保守,这与其他植物[21]上的研究相似。
从花椰菜叶绿体基因组中检测到228个单核苷酸,在A、C和T 3种类型中,其中,A/T类型占96.49%,18个二核苷重复全部为AT/TA类型,62个三核苷酸重复中全部由A、T碱基构成的占比为37.10%,这与张明英等[21]研究的美丽芍药基因组中三核苷酸全部由A、T碱基组合构成的结果不一致。
IR边界分析发现,在SSC/IRa边界,紫色花椰菜与其他不同,4种花椰菜有明显的不同,ycf1向SSC区收缩了111 bp,向SSC区扩张了14 bp,与 2种甘蓝(Brasrsicaoleraceavar.capitata;Brasrsicaoleraceavar.italica)和芥菜(Brasrsicanapus)相同,而在LSC/IR和SSC/IR边界区有相同的基因种类和排列顺序,存在不同程度的扩张和收缩,且与大多数十字花科植物相似。野芥(Sinapisarvensis)和海甘蓝(Crambehispanicasubsp.abyssinica)SSC/IRb边界则与其他种稍有区别,海甘蓝ycf1扩张程度与其他种相同,而IRb区则注释为72 bp的trnN向内收缩1 356 bp,野芥则表现为ycf1向SSC区扩张了44 bp;在IRa/LSC边界,有3种植物有明显不同,蓝花子的ycf1向LSC扩张了511 bp,trnM向LSC收缩了599 bp,甘蓝的trnA向内收缩3 bp,野芥有完全不同的基因P31pgt向LSC收缩了2 bp,其他植物的IRa/LSC边界均处于rpl2和trnH之间,其中,rpl2向LSC区收缩167 bp,trnH向IRa区收缩3 bp,在此边界,4种花椰菜均无区别。
本研究中,十字花科叶绿体全基因组序列构建的系统发育树,分成为两大分支,其中一支包括甘蓝种、芥菜种、欧洲油菜种,全部为芸薹属;而第二支,芸薹属的黑芥和野芥亲缘关系最近,且它们与海甘蓝亲缘关系更密切,这与芝麻菜[16]、海甘蓝[18]叶绿体基因组上的研究结果相似,碎米芥属和紫罗兰属亲缘关系最近,它们与诸葛菜属、菥蓂属(遏蓝菜)聚为一类,之后与黑芥、野芥和海甘蓝分支聚在一起,最后与萝卜属聚为一支。本研究分析了4种花椰菜与部分十字花科部分植物的亲缘关系,将为研究十字花科植物的系统进化关系提供一定参考。
