甘薯CLE 多肽家族的全基因组鉴定和IbCLE12 的克隆及抗旱性表达分析
2023-06-19周媛媛赵春玲侯夫云李爱贤秦桢王庆美董顺旭
周媛媛,赵春玲,侯夫云,李爱贤,秦桢,王庆美,董顺旭
(1. 山东省农业科学院作物研究所,山东济南 250100;2. 山东师范大学,山东济南 250014)
多肽类激素与脱落酸、茉莉酸、细胞分裂素、生长素、赤霉素、乙烯等传统植物激素一起,在植物抵御逆境胁迫过程中起重要作用,近年来多肽激素的研究进展较快[1,2]。 CLAVATA3/Embryo surrounding region-related (CLE)多肽家族是研究最多的一类多肽激素[3,4],其命名依据是拟南芥CLAVATA3(CLV3)和玉米胚胎周边区(embryo surrounding region, ERS);在结构上,一般编码50~200 个氨基酸的小蛋白,N-末端存在一个分泌信号肽,C-末端存在一个含14 个氨基酸的保守结构域,中间是可变区域[5];根据CLE motif 的首个氨基酸类型将其分为R(arginine,Arg)和H(histidine, His)两类,这两类之外的统称other类[6]。 相关研究表明,CLE 家族成员在根、茎尖分生组织的干细胞分裂和分化、维管(原)形成层、激素信号响应以及抗逆性响应中发挥着关键性作用[7-9]。 目前,在拟南芥中发现32 个CLE 成员[6],其中,CLE25 多肽在根部响应干旱胁迫,并通过BAM 受体将缺水信号传导至叶片,与ABA共同调控气孔的关闭,减少蒸腾作用,从而提高对干旱的抗性[9]。
甘薯[Ipomoea batatas(L.) Lam.]是世界主要农作物之一,可用作粮食、饲料、工业原料和新型能源,广泛种植于100 多个国家或地区,具有重要的经济价值[10,11]。 甘薯在生长过程中经常遭受干旱等逆境胁迫[12],所以研究CLE 家族成员及其功能对提高甘薯抗逆性具有重要意义。 目前已有57 个植物基因组中的1628 个CLE 多肽得到预测,但在甘薯中尚未见研究报道。 本研究利用野生型甘薯Ipomoea trifida(H.B.K.) G. Don.的基因组[13],系统挖掘甘薯CLE 家族成员,利用生物信息学方法进行分析,并进一步从甘薯栽培种中克隆得到IbCLE12基因,利用实时荧光定量法分析其在干旱胁迫下的表达,以期为探究甘薯CLE 多肽家族基因的功能奠定基础。
1 材料与方法
1.1 试验材料
甘薯近缘二倍体野生种Ipomoea trifida(H.B.K.) G. Don.,是栽培甘薯的祖先种之一,其基因组测序已经完成。 甘薯栽培种选用济薯25。
1.2 Ipomoea trifida CLE 家族基因的全基因组鉴定及分析
1.2.1 甘薯CLE 家族基因的筛选 基于甘薯组学资源网GT4SP 项目的基因注释文件(http:/ /sweetpotato.plantbiology.msu.edu/index.shtml)查找所有已注释的ItfCLE基因,并下载相关序列。
1.2.2ItfCLE基因结构分析 利用ProtParam 在线软件(https:/ /web.expasy.org/protparam)分析ItfCLE 蛋白理化性质。 利用TMHMM 2.0(https:/ /services.healthtech.dtu.dk/service.php? TMHMM-2.0)分析基因的跨膜螺旋位点,利用Soft-Berry ProtComp 9.0 预测蛋白亚细胞定位情况。利用MEME (http:/ /meme-suite.org/tools/meme)分析ItfCLE 氨基酸序列的保守结构域,参数设置为允许保守结构域重复出现、搜索得到的保守域上限为6 个、基序长度为10 ~100。 利用TBtools工具绘制ItfCLE基因结构图。
1.2.3 ItfCLE 家族的系统发育分析 采用MEGA 5.0 软件的NJ 法,基于保守CLE motif 氨基酸序列,构建ItfCLEs 和拟南芥AtCLEs 的系统发育进化树,Bootstrap 重复1000 次。
1.2.4ItfCLEs基因的组织特异性表达和不同逆境下的表达谱分析 利用甘薯基因组学资源网站中的RNA-seq 基因表达数据,用FPKM(fragments per kilobase per million mapped reads)值表示基因相对表达水平,利用TBtools 建立热图,进行不同基因间聚类。
1.3 IbCLE12 基因的克隆及表达分析
1.3.1IbCLE12基因的克隆 利用TRIzol 试剂盒(Invitrogen,上海,中国)提取总RNA,利用TaKa-Ra 反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)对样品进行反转录,获得济薯25 的cDNA,利用TaKaRaLA Taq试剂盒扩增IbCLE12基因的完整CDS 序列。 扩增引物为IbCLE12-F(5'-ATGGGTAGTAATTTGAGGAGTAGGA-3')和IbCLE12-R(5'-TCATGGCTTGTCAAGCTCTCC-3')。 扩增体系(50 μL):LA Taq0.5 μL,LA Buffer(Mg2+plus)8 μL,dNTP Mixture 5 μL,引物混合物2 μL,cDNA 2 μL,ddH2O 32.5 μL。 扩增程序:94℃3 min;94℃30 s,55℃30 s,72℃30 s,34 个循环;72℃10 min。
1.3.2 干旱胁迫和ABA 处理下IbCLE12基因的表达分析 用2021 年收获的济薯25 薯块,在温室中培养4 周,取长势一致、含5~6 个功能叶的茎段,分别用含20% PEG6000 和100 mmol/L ABA 的1/2 霍格兰溶液处理,分别于处理0、3、6、12、24、48 h取叶片,用液氮速冻,-80℃保存备用。
利用Invitrogen TRIzol 试剂盒提取总RNA,利用TaKaRa反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser 对每个样品进行反转录,利用SYBR PremixEx TaqⅡ定量试剂(Tli RNaseH Plus, TaKaRa, 大连, 中国) 在Roche LightCycler®480Ⅱ(Roche,英国)进行qRT-PCR分析。 扩增引物为ItfCLE-qPCR-F(5'-TATTGGTGGGAGTGGTTGGGT-3')和ItfCLE-qPCR-R(5'-CATGCTCATATGCCTCATGCT-3')。 扩增体系:SYBR Green Master Mix 5 μL,引物混合物0.4 μL,cDNA 1 μL,ddH2O 3.6 μL。 反应程序:95℃预变性2 min;95℃变性15 s,55℃退火15 s,72℃延伸15 s,40 个循环。
使用内参基因IbActin为校准,利用2-ΔΔCt计算基因的相对表达水平。 每个样品进行3 次重复试验,以未处理(0 h) 的样品为对照,采用Student’st-test 方法进行显著性分析。
2 结果与分析
2.1 ItfCLE 基因的筛选和蛋白特征分析
从I.trifida中筛选到12 个CLE 家族基因,根据其在染色体上的位置进行重新编号,分别命名为ItfCLE1~ItfCLE12(表1)。 这12 个ItfCLE 蛋白长度在81 ~154 个氨基酸之间,分子量在9.0 ~17.5 kDa 之间,等电点均大于7,属于碱性蛋白。只有ItfCLE2 和ItfCLE6 为稳定蛋白,其余均为不稳定蛋白,且ItfCLE3 不稳定系数最高。 除分子量最大的ItfCLE5 为疏水脂溶蛋白外,其余均为亲水性脂溶蛋白。 根据保守区的首个氨基酸类型分类,只有ItfCLE6 属于H 型,其余均为R 型。
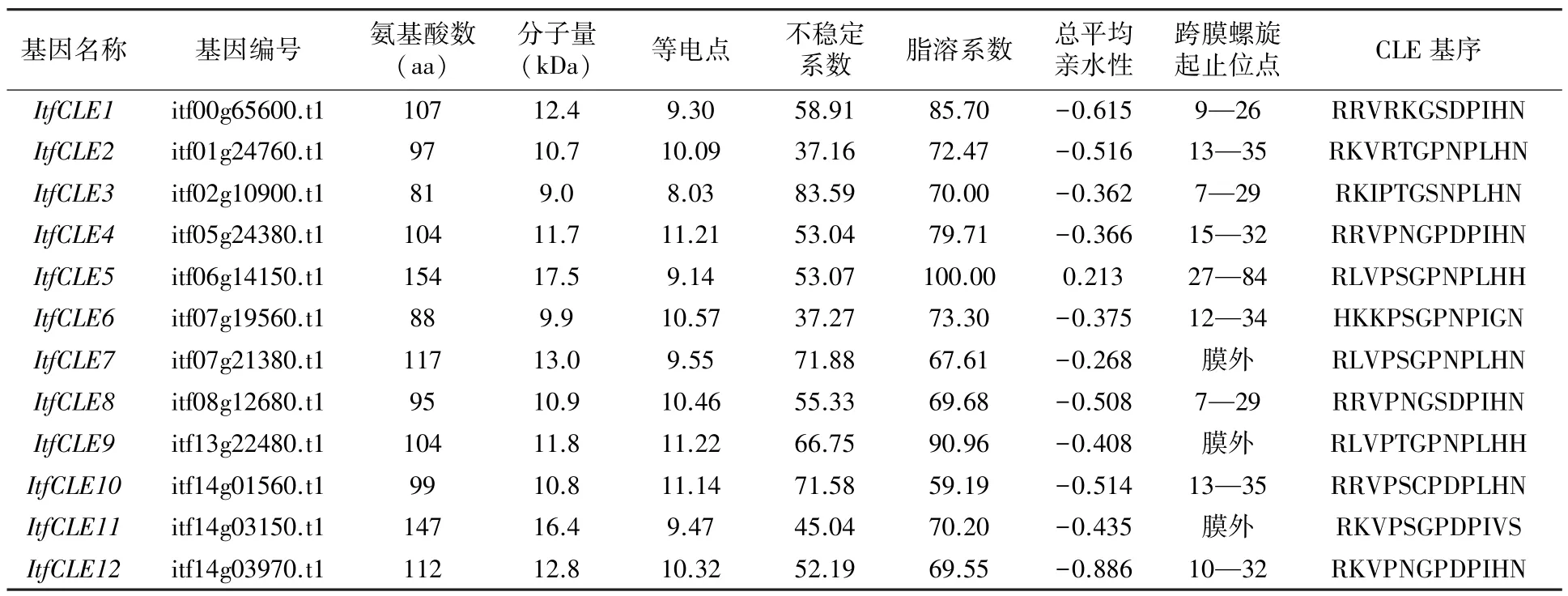
表1 ItfCLEs 蛋白理化性质
利用MEME 软件分析ItfCLE 蛋白的保守结构域,共获得6 个保守基序,分别命名为Motif1 ~Motif6。 12 个 ItfCLE 成员均含有 Motif1(VSKRKVPSGPBPJHN),所以,Motif1 是甘薯CLE的特征保守基序(图1)。
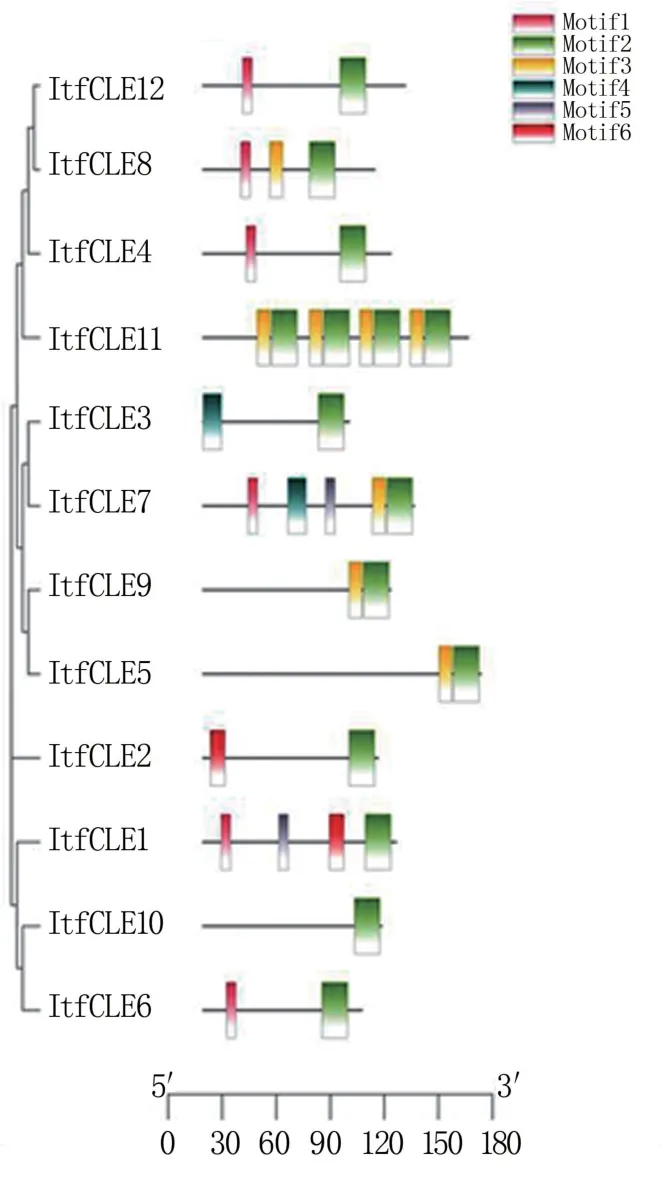
图1 ItfCLE 蛋白保守基序分析
根据蛋白定位预测结果(表2),12 个ItfCLE蛋白在胞外都有分布;仅ItfCLE9 在质膜中有分布,其余蛋白则在细胞质、过氧化物酶体以及叶绿体中有分布。 据此推测ItfCLE 蛋白可能主要存在于过氧化物酶体、叶绿体以及细胞质中,外排到胞外。
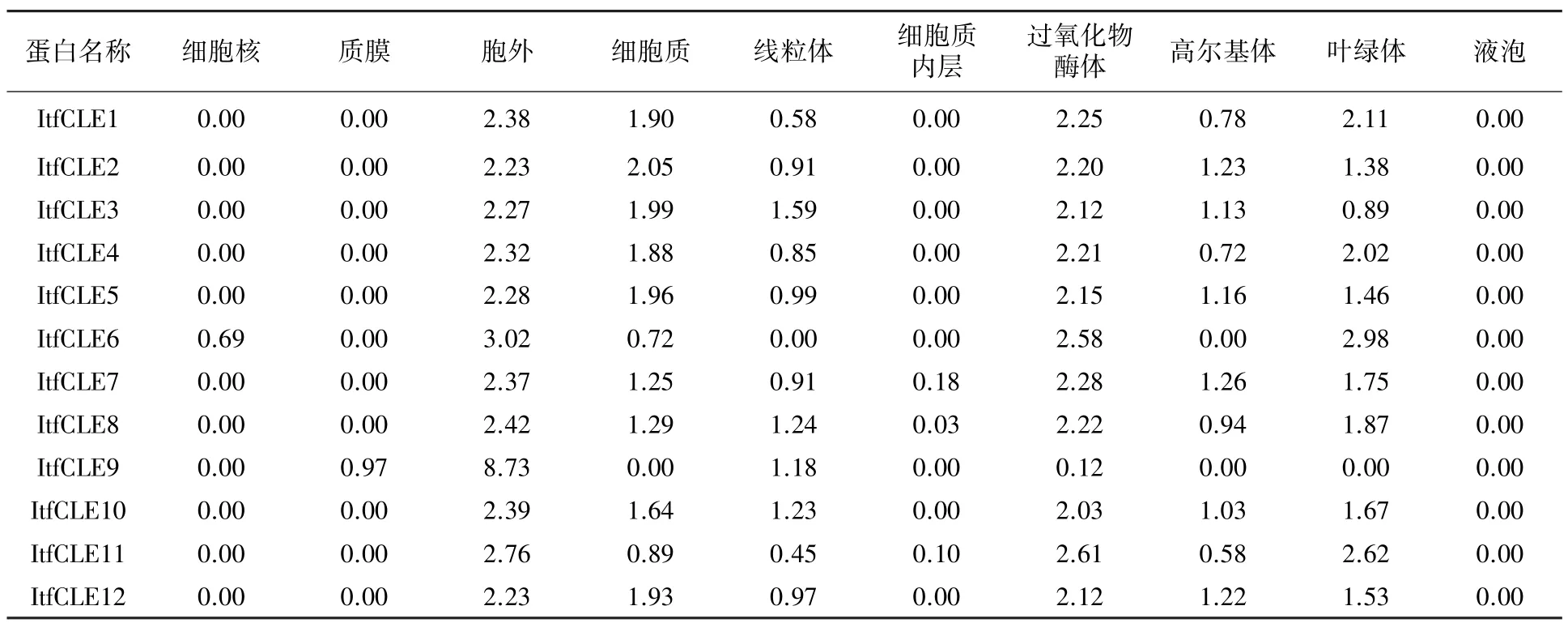
表2 ItfCLE 蛋白的定位分析
2.2 ItfCLEs 的进化分析
通过ItfCLEs 与AtCLEs 的进化关系树(图2)可知,甘薯的ItfCLEs 主要分为5 个亚族。 Itf-CLE1、 ItfCLE4、 ItfCLE6、 ItfCLE8、 ItfCLE11、 Itf-CLE12 亲缘关系较近,其中,ItfCLE1、ItfCLE8、Itf-CLE4 与拟南芥的AtCLE45 属于同一组,ItfCLE12与AtCLE25 属于同一组,ItfCLE6 与AtCLE46 属于同一组;ItfCLE10 与AtCLE27 亲缘关系较近;ItfCLE3 和ItfCLE2 属于同一亚族,并与AtCLE20 亲缘关系较近;ItfCLE7 与AtCLE9、AtCLE10 亲缘关系较近;ItfCLE5 和ItfCLE9 属于同一亚族,分别与AtCLE13 和AtCLE8 亲缘关系较近。
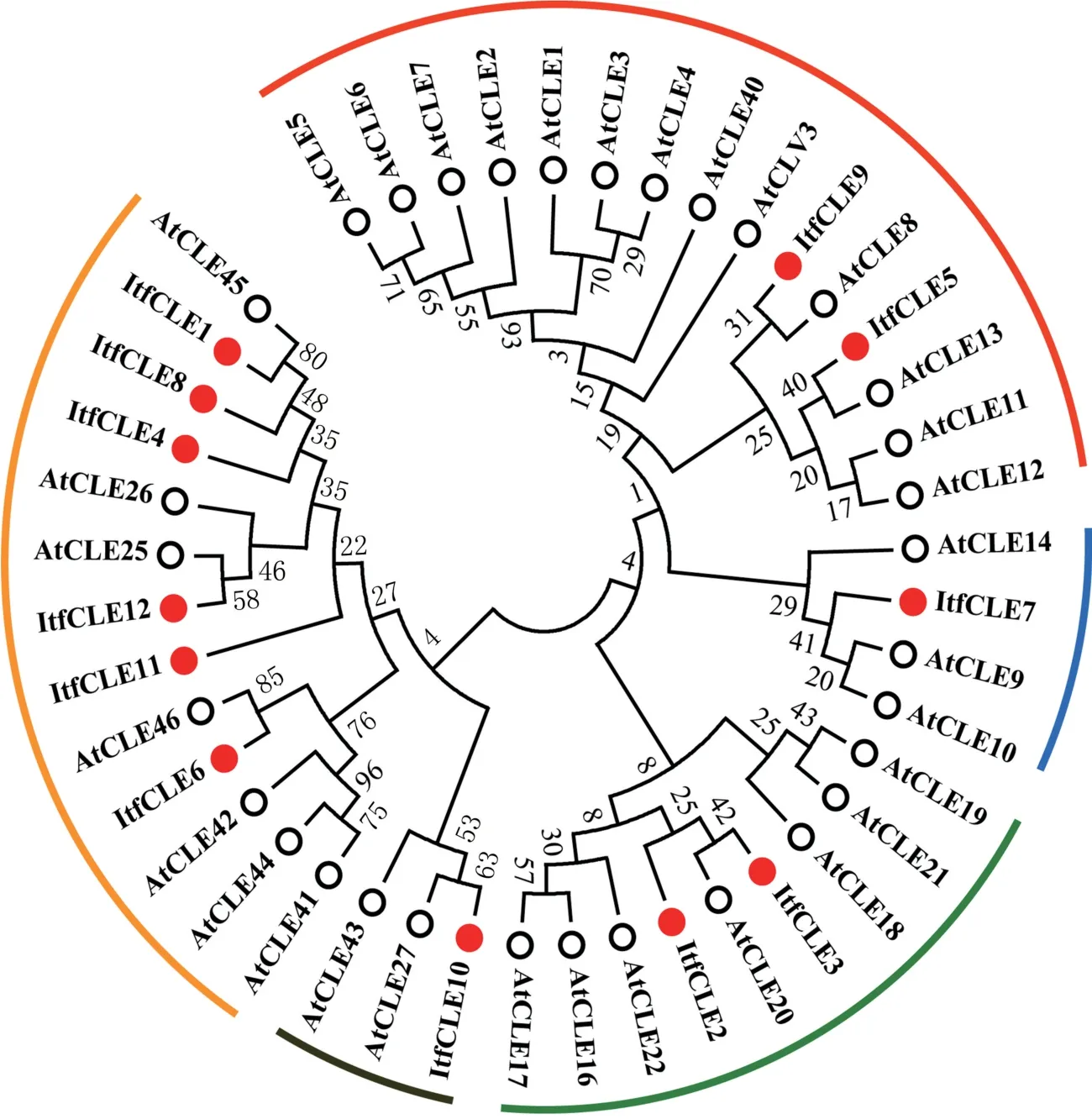
图2 甘薯12 个ItfCLEs 和拟南芥32 个AtCLEs 进化树分析
2.3 ItfCLE 基因的组织特异性表达分析
结果(图3)表明,ItfCLE9、ItfCLE10在花中有较高的表达;ItfCLE4、ItfCLE12在根中有较高的表达;ItfCLE1、ItfCLE3、ItfCLE5、ItfCLE7、ItfCLE11在各组织中表达量较低,差异较小;ItfCLE2、ItfCLE6和ItfCLE8在茎中表达较高。
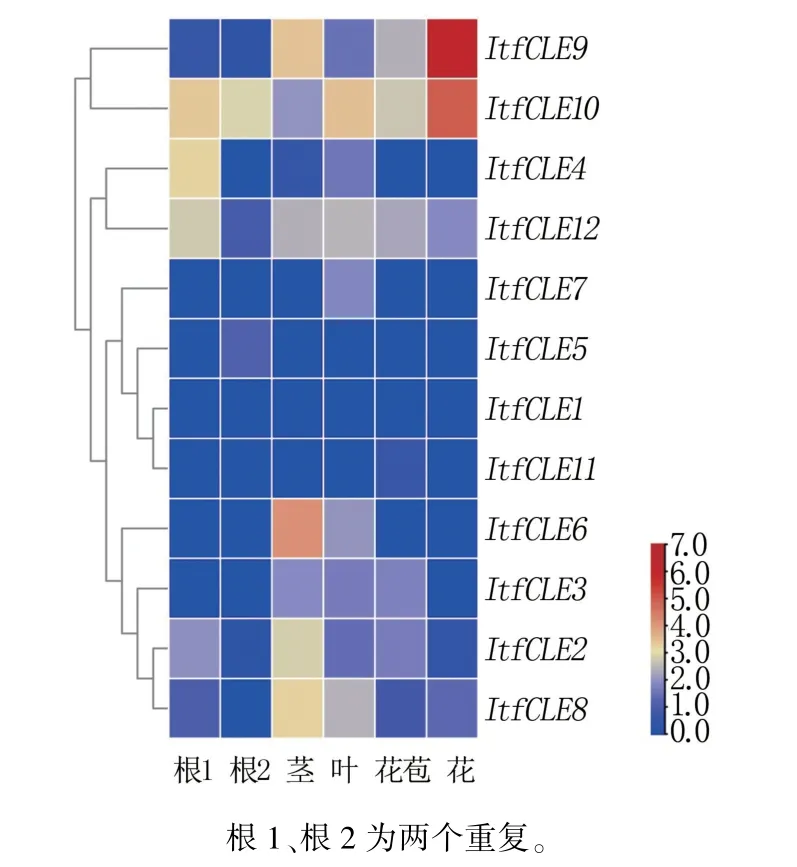
图3 甘薯12 个ItfCLE 基因的组织特异性表达分析
2.4 ItfCLE 基因在不同逆境胁迫下的表达特征分析
结果(图4)表明,与对照相比,ItfCLE10受旱、盐胁迫和两种病害处理后下调表达;ItfCLE8受旱、盐胁迫后上调表达,受冷和热胁迫后下调表达;ItfCLE12受旱胁迫和病2 处理后上调表达,受盐、冷和病1 胁迫后下调表达;ItfCLE3受盐、冷、热胁迫后下调表达。 其余基因受胁迫后表达量变化较小。
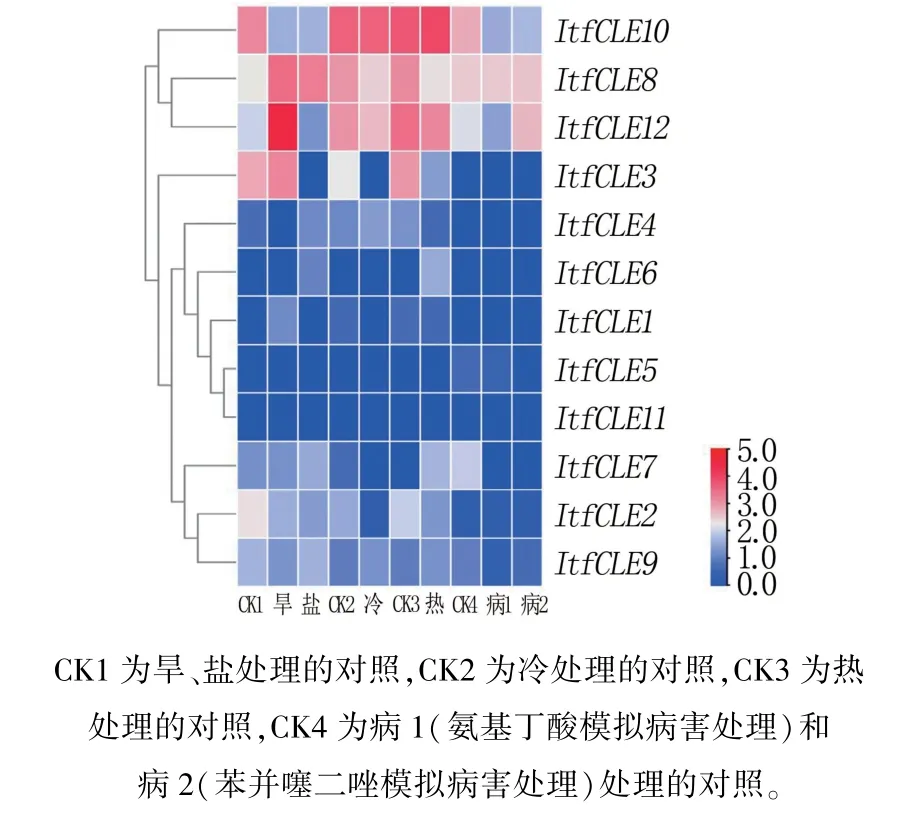
图4 甘薯12 个ItfCLE 基因的表达谱分析
2.5 IbCLE12 基因的克隆和氨基酸序列比对
利用同源克隆方法在济薯25 中克隆得到Itf-CLE12的同源基因,命名为IbCLE12。 氨基酸序列比对结果(图5)表明,IbCLE12 与ItfCLE12 一致性较高,仅在第23、26 位有两个氨基酸的差异;IbCLE12与AtCLE25的一致性达到32.74%,两者C 端保守CLE 基序一致。

图5 IbCLE12 与ItfCLE12、AtCLE25 的序列比对
2.6 IbCLE12 基因受干旱和ABA 诱导表达分析
结果(图6)表明,IbCLE12基因受干旱和ABA 诱导上调表达,分别在处理6 h(5.48 倍)和12 h(5.06 倍)表达量最高。 处理时间延长,表达量显著降低。

图6 干旱(A)和ABA(B)胁迫下IbCLE12 基因的表达情况
3 讨论与结论
CLE 家族基因编码的多肽是21 世纪以来发现的新型多肽激素,在细胞间的信号交流中发挥重要作用[3,5,14]。 CLE 蛋白信号肽使蛋白迁移至细胞膜,并与膜上的受体激酶特异性结合,将信号转导至下游分生组织发育的WUSCHEL(WUS)转录因子,形成信号转导模块,从而调控分生组织的干细胞增殖和分化,最终影响根、茎等器官的分化[15]。 甘薯野生种I.trifida中包含12 个CLE 家族基因,蛋白C 端均包含CLE 保守结构域,家族基因数量少于拟南芥等模式植物,可能与数据库组装过程中小分子基因的覆盖度相关[16]。 甘薯作为块根作物,根中分生组织的干细胞分化和增殖直接影响形成层的发育,并最终与产量密切相关。 有研究表明,通过对人参CLE 家族序列特征和系统进化关系进行分析,发现PgCLE基因对其根型发育起重要的调控作用[17]。 本研究结果表明,ItfCLE在野生型甘薯中存在明显的组织表达差异性,其中,ItfCLE9、ItfCLE10在花中有较高的表达;ItfCLE4、ItfCLE12在根中有较高的表达;Itf-CLE2、ItfCLE6和ItfCLE8在茎中表达较高;Itf-CLE1、ItfCLE3、ItfCLE5、ItfCLE7、ItfCLE11在各组织中表达量较低,差异较小。 可见,甘薯中CLE成员在不同器官发育过程中可能发挥不同的调控作用。
CLE 家族基因在作物响应生物胁迫中也具有重要调节作用。 研究表明,在大豆囊线虫、马铃薯囊线虫和根结线虫中具有和宿主类似的CLE,并通过与宿主中的CLE 受体相互作用,提高线虫对宿主的侵染力[18,19]。 敲除CLE 受体可以显著提高大豆对大豆囊线虫的抵抗能力[20]。 本研究结果表明,与对照相比,在两种病害胁迫下大部分ItfCLEs表达量变化不大,但ItfCLE7、ItfCLE10明显下调表达,ItfCLE12在病1 处理下也表达下调,推测其在生物胁迫中主要起负调控作用。
CLE 家族基因在作物抵御非生物胁迫中也发挥重要作用。 研究表明,拟南芥的AtCLE25 多肽可以实现从根到芽的长距离信号转导,并通过介导ABA 信号途径促进气孔关闭,感应根的缺水信号,提高植物的抗旱性[9];AtCLE45通过下游受体SKM1 和SKM2 调节拟南芥种子形成过程中的耐热性[21]。 在拟南芥缺失突变体cle40-2中,ABA合成通路上的AtCLE25、AtCLE45基因表达下调,说明其在ABA 合成通路上起关键的正调控功能[22]。 在本研究中,受旱胁迫后明显上调表达的有ItfCLE8和ItfCLE12,而ItfCLE12 与拟南芥At-CLE25 亲缘关系较近,推测ItfCLE12在甘薯抗旱性中发挥调控作用。 为此,我们在甘薯栽培品种中克隆了ItfCLE12的同源基因IbCLE12,并进行模拟干旱和ABA 处理,结果表明该基因分别在处理6 h 和12 h 受干旱和ABA 诱导显著上调表达,可为深入研究IbCLE12在甘薯抗旱性中的分子机制奠定基础。
