吊罗山热带天然林物种多样性的海拔分布格局
2023-06-18李晨笛梁宜文杨小波李东海
李晨笛 梁宜文 杨小波 李东海


摘 要: 海拔梯度是影響物种多样性格局的关键因素。为探究吊罗山热带天然林物种多样性的海拔分布格局,该研究对吊罗山245~1 130 m海拔内的6个热带天然林样地进行植被调查,从群落物种组成、物种多样性、区系组成和叶性质等方面分析沿海拔梯度吊罗山热带天然林物种多样性与群落特征变化规律。结果表明:(1)随海拔升高、温度降低、湿度升高、人为干扰减少,吊罗山热带天然林物种组成以及Margalef、Shannon-Wiener、Simpson、Pielou多样性指数均呈现先升高后降低的格局;中海拔群落水热条件最为适宜、人为干扰适中、α物种多样性最高。(2)随海拔升高,优势种重要值占比呈现先降低后升高的格局,且优势种具有明显的更替现象,中海拔群落呈现低地雨林与山地雨林的过渡性质。(3)群落间Srensen群落相似度和海拔高度差呈显著负相关(P<0.05)。(4)区系组成以热带区分布为绝对优势,并以热带亚洲(印度-马来西亚)分布为主;热带区分布比例和海拔呈极显著负相关(P<0.01),温带区分布比例和海拔呈极显著正相关(P<0.01)。(5)叶性质呈现出以中型叶、单叶、革质叶、全缘叶为主的热带地区显著特点;小型叶、单叶比例和海拔呈极显著正相关(P<0.01),革质叶、非全缘叶比例和海拔相关性不显著(P>0.05)。综上所述,吊罗山低海拔群落水热条件和人为干扰程度都与高海拔群落存在较大差异,从而导致了物种分布和群落特征的差异性,体现出热带植物与其生境条件相适应的特点。
关键词: 海拔梯度, 热带天然林, 物种组成, 物种多样性, 区系, 叶性质
中图分类号: Q948 文献标识码: A 文章编号: 1000-3142(2023)05-0912-11
Abstract: Altitude is the key factor affecting species diversity pattern. In order to explore the altitude distribution pattern of species diversity in tropical natural forest in Diaoluo Mountain, vegetation survey was conducted in six tropical natural forest plots from 245 to 1 130 m. The species diversity and community characteristics of tropical natural forest along the altitude of Diaoluo Mountain were analyzed from the aspects of species composition, species diversity, flora composition and leaf characteristics. The results were as follows: (1)With the increase of altitude, the temperature decreased, the humidity increased and the human disturbance decreased. The species composition of tropical natural forest in Diaoluo Mountain and the diversity indexes of Margalef, Shannon-Wiener, Simpson, and Pielou all showed a pattern of first increase and then decrease. The middle-altitude community had the most suitable hydrothermal conditions and moderate human disturbance, and the α species diversity was the highest. (2)With the increase of altitude, the proportion of important value (IV) of dominant species decreased first and then increased, and the dominant species had obvious replacement phenomenon. The middle altitude community showed the transition nature of lowland rain forest and mountain rain forest. (3) The Srensen community similarity coefficient between communities was significantly negatively correlated with altitude difference (P<0.05). (4)The floristic composition was dominated by tropical elements and mainly distributed in tropical Asia (India-Malaysia). The proportion of tropical distribution was significantly negatively correlated with altitude (P<0.01), and the proportion of temperate distribution was significantly positively correlated with altitude (P<0.01). (5)The leaf characteristics showed the obvious indigenous characteristics in tropical regions mainly with mesophyll leaves, single leaves, leathery leaves and entire leaves. The proportion of microphyll leaves and single leaves was significantly positively correlated with altitude (P<0.01), while the proportion of leathery leaves and non-entire leaves was not significantly correlated with altitude (P>0.05). To sum up, the hydrothermal conditions and human disturbance degree of low altitude community in Diaoluo Mountain are quite different from those of high altitude community, which leads to the difference of species distribution and community characteristics, and reflects the characteristics of tropical plants adapting to their habitat conditions.
Key words: altitude, tropical natural forest, species composition, species diversity, flora, leaf characteristics
物种多样性对反映群落物种的丰富程度、群落结构的复杂程度和生态系统的稳定性具有重要意义(Legendre et al., 2009;Ghaley & Porter, 2014)。不同生境条件下,植物群落物种组成各不相同,海拔梯度不仅改变了群落中光照、温度、湿度等环境因子(Gaston, 2000; Simsim et al., 2015),甚至影响树木受人为干扰的程度,是影响物种多样性格局的关键因素之一(Brown, 2001)。一般而言,溫度随海拔升高而降低,湿度随海拔升高而升高,树木受人为干扰的影响随海拔上升呈下降趋势(龙文兴,2016)。前人将物种多样性在海拔层面的分布格局一般分为5种(贺金生和陈伟烈,1997),分别是与海拔呈负相关的单调降低格局、与海拔呈正相关的单调升高格局、随海拔梯度先升高后降低的格局、随海拔梯度先降低后升高的格局以及与海拔相关性不显著的格局。如在人为干扰小的湿润地区,温度是影响群落物种垂直分布的第一要素,随海拔升高,温度降低,物种多样性也随之下降(唐志尧和方精云,2004)。在干旱地区,随海拔升高,虽温度下降,但雨水增多,有利于物种多样性的提高,然而山顶高寒且风巨的严峻生境又导致物种多样性降低,故呈现出先升高后降低的物种分布格局(龙文兴,2016)。因此,受水热、光照、群落特性、外部干扰等多重因素影响,沿山体海拔,物种分布具有差异性(龙文兴,2016)。而不同的生境条件亦使群落区系组成和外貌特征各不相同。受温度变化影响,随海拔升高,群落区系呈现出热带成分减少、温带成分增加的特点(朱华,2008);较少的人为干扰,使高海拔群落中国特有分布增加(苏文苹,2007),区系地理成分更加复杂(巫翠华等,2021)。随海拔升高,川滇高山栎(Quercus aquifolioides)群落叶性质呈现出细微型叶比例升高、纸质叶比例降低的特点(刘兴良等,2013),体现了植物与其生境紧密适应的特征(吕晓波,2012)。因此,研究物种多样性的海拔分布格局,分析随海拔梯度群落特征变化规律,对揭示树木与海拔的内在联系具有重要意义。
海南省吊罗山国家森林公园是我国重要的热带森林分布区之一,具有我国热带地区发育最盛、最接近赤道热带雨林的热带森林(江海声,2006),沿山体海拔分布着热带低地雨林、热带山地雨林等植被类型。前人对吊罗山热带天然林的物种多样性、区系组成、群落特征进行了大量研究(安树青等,1999;丁坦等,2002;王帅等,2015;韩天宇等,2019),但主要基于单一植被类型,而沿海拔梯度吊罗山热带天然林的物种多样性和群落特征变化规律尚待研究。本研究以吊罗山热带低地雨林至热带山地雨林内的6个固定样地为对象,拟探讨以下问题:(1)吊罗山热带天然林物种多样性随海拔梯度的变化规律;(2)海拔梯度对群落区系组成的影响;(3)海拔梯度对群落叶性质的影响。以期为不同海拔热带森林的物种多样性保护和生态管理提供依据。
1 材料与方法
1.1 研究区域概况
吊罗山国家森林公园位于海南岛东南部的109°41′38″—110°4′46″ E、 18°38′42″—18°50′22″ N之间。年均温度为20.8 ℃,属东亚海洋性热带季风气候,年降雨量充足,降雨量为1 800~2 800 mm,干湿季明显。吊罗山具有鲜明的山地垂直气候变化,海拔每升高100 m,气温递减0.6 ℃,500 m以上的高海拔地区具有明显的低温期(江海声,2006)。随着海拔高度的变化,降雨量、相对湿度递增,云雾增多。沿山体海拔相应出现热带低地雨林、热带山地雨林、山顶常绿矮林及灌丛等(丁坦等,2002)。成土母岩为花岗岩和闪长岩,300 m海拔以下土壤为黄色砖红壤,300 m海拔以上土壤为山地黄壤(王峥峰等,1999)。吊罗山地区山泉众多,水资源非常丰富,光、热和水分等自然环境较优越,孕育了丰富的植物资源。
1.2 样地设置和调查
为探明吊罗山不同海拔热带天然林的物种多样性和群落特征,在吊罗山热带天然林内设置6个永久固定样地(Plot 1-Plot 6),样地沿245~1 130 m海拔分布,具有明显海拔高度差,群落结构完整,发育较好。样地设置见表1,其中地处245~475 m海拔的Plot 1和Plot 2属热带低地雨林,地处555~665 m海拔的Plot 3和Plot 4属热带低地雨林与热带山地雨林过渡性质,地处940~1 130 m海拔的Plot 5和Plot 6属热带山地雨林。每块样地投影面积为2 500 m2(50 m × 50 m),共计1.5 hm2。每块样地以西南角作为原点,采用“相邻格子法”划分为5个50 m × 10 m中样方,再将50 m × 10 m中样方划分为10 m × 10 m小样方,共150个。采用“每木记账调查法”,以胸径(diameter at breast height,DBH)≥3 cm为木本植物的起测径阶,对每木进行编号、挂牌,记录其胸径、树高、冠幅等。通过查阅《海南植物图志》和Flora of China,对样地内木本植物进行物种鉴定(杨小波,2016)。
1.3 重要值计算
重要值(important value,IV)的计算公式如下:
重要值=(相对密度+相对频度+相对显著度)/3。依据物种重要值的大小确定样地优势种(龙文兴,2016)。
1.4 物种多样性指数的计算
1.4.1 群落α多样性 用Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson多样性指数、Pielou均匀度指数来测定(Lakhani & Magurran, 1989)。
Margalef丰富度指数:
DMa=S-1lnN。
Shannon-Wiener多样性指数:
H=-∑Si=1PilnPi。
Simpson多样性指数:
C=∑Si=1Ni(Ni-1)N(N-1)。
Pielou均匀度指数:
E=HlnS。
式中:S表示物种数目; N表示群落中所有种的个体总数; Pi表示第i个物种的相对多度,Pi = Ni/N,Ni表示第i个种的个体数目。
1.4.2 β多样性 用Srensen群落相似度来测定(龙文兴,2016)。
Srensen群落相似度计算公式如下:
Cs=2ca+b。
式中:a、b是兩个样地中各自的物种数;c是两个样地中的共有种数。
1.5 群落区系分析
依据吴征镒等(2006)种子植物属的分布区类型系统,对不同海拔热带天然林样地的植物区系组成进行分析。
1.6 群落外貌特征分析
依据Raunkiaer叶级划分系统和Paijimans叶质分类系统(王伯荪,1987),对不同海拔热带天然林样地群落的叶性质进行分析。按叶级将叶划分为鳞型叶、微型叶、小型叶、中型叶、大型叶和巨型叶,共六级;按叶型将叶划分为单叶和复叶;按叶质将叶划分为革质叶、纸质叶和膜质叶;按叶缘将叶划分为全缘叶和非全缘叶。
1.7 数据处理
数据统计与制图通过Microsoft Excel 2021软件和Origin 2021软件完成。利用Origin 2021软件计算Pearson相关系数和Spearman秩相关系数,对海拔差与Srensen群落相似度、海拔与区系组成、海拔与叶性质的相关性进行分析。
2 结果与分析
2.1 物种组成
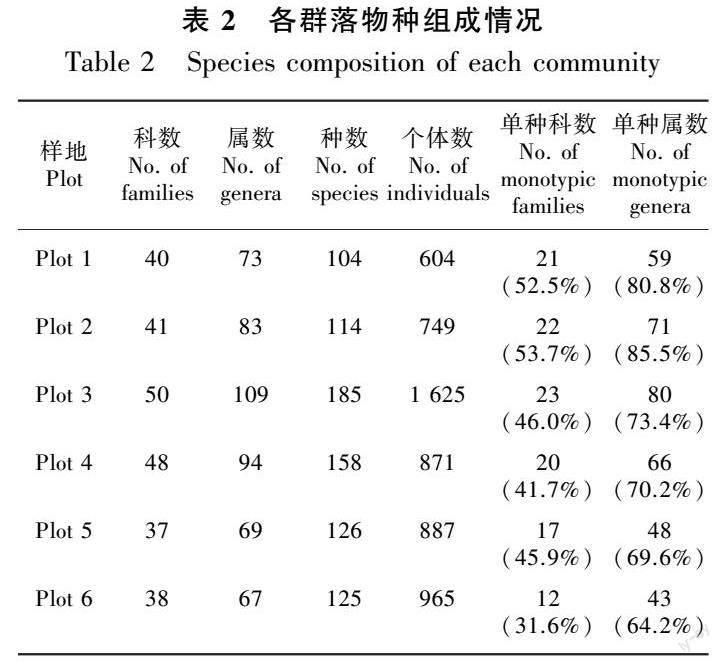
根据调查结果显示(表2),吊罗山热带天然林物种组成丰富,随海拔升高,群落科、属、种组成呈现先升高后降低的格局,地处中海拔地区且属于热带低地雨林与热带山地雨林过渡性质的Plot 3、Plot 4物种组成最丰富。随海拔升高,群落单种科和单种属数量呈现先升高后降低的格局,低地雨林群落单种科和单种属数量高于山地雨林。吊罗山不同海拔热带天然林群落单种属均占有较高比例,为64.2%~85.5%,表明在属的组成上具有较高的分散性。
从优势科来看,Plot 1拥有5个种以上的科有7个,其中樟科(Lauraceae)(11种)、桃金娘科(Myrtaceae)(10种)、大戟科(Euphorbiaceae)(7种)、豆科(Leguminosae)(7种)、柿科(Ebenaceae)(6种)占较大优势。Plot 2拥有5个种以上的科有5个,分别为樟科(21种)、茜草科(Rubiaceae)(10种)、桃金娘科(10种)、山茶科(Theaceae)(5种)、芸香科(Rutaceae)(5种)。Plot 3拥有5个种以上的科有13个,其中樟科(19种)、大戟科(17种)、桃金娘科(12种)、茜草科(11种)、山矾科(Symplocaceae)(11种)占较大优势。Plot 4拥有5个种以上的科有8个,其中樟科(24种)、桃金娘科(11种)、壳斗科(Fagaceae)(10种)、山茶科(10种)、茜草科(8种)占较大优势。Plot 5拥有5个种以上的科有5个,分别为樟科(18种)、壳斗科(14种)、山矾科(11种)、山茶科(9种)、桃金娘科(8种)。Plot 6拥有5个种以上的科有6个,其中樟科(18种)、壳斗科(13种)、山矾科(10种)、山茶科(7种)、桃金娘科(6种)占较大优势。由此可见,吊罗山热带天然林多以樟科、桃金娘科、山茶科、茜草科、山矾科等为优势科,其中广泛分布于热带地区的樟科占主要优势地位,表明群落具有较强的热带性。
2.2 优势种
森林群落优势种对构建群落起着决定性作用,调查结果显示(表3),Plot 1群落以托盘青冈(Cyclobalanopsis patelliformis)、粘木(Ixonanthes reticulata)、白颜(Gironniera subaequalis)、柬埔寨子楝树(Decaspermum montanum)、黄杞(Engelhardia roxburghiana)等为优势种,其中托盘青冈占绝对优势,粘木占相对优势。Plot 2以鹅掌柴(Schefflera heptaphylla)、肉实树(Sarcosperma laurinum)、青藤公(Ficus langkokensis)、狭叶泡花树(Meliosma angustifolia)、大花五桠果(Dillenia turbinata)等为优势种,其中鹅掌柴优势地位明显。Plot 3以贡甲(Maclurodendron oligophlebium)、白颜、柬埔寨子楝树、白肉榕(Ficus vasculosa)、九节(Psychotria asiatica)等为优势种,优势种重要值相近,为多优势种群落。Plot 4以柬埔寨子楝树、陆均松(Dacrydium pectinatum)、鹅掌柴、钝齿木荷(Schima crenata)、海南杨桐(Adinandra hainanensis)等为优势种,优势种重要值相差不大,为多优势种群落。Plot 5以米锥(Castanopsis chinensis)、海南杨桐、陆均松、 尖峰润楠 (Machilus monticola)、 五列木(Pentaphylax euryoides)等为优势种,其中米锥占绝对优势。Plot 6以岭南青冈(Cyclobalanopsis championii)、琼南柿(Diospyros howii)、黄叶树(Xanthophyllum hainanense)、陆均松、岭南柿(Diospyros tutcheri)等为优势种,其中以岭南青冈占绝对优势,琼南柿占相对优势。
由此可见,随海拔升高,群落优势种具有明显的更替现象,中海拔群落呈现出低地雨林与山地雨林过渡性质。中海拔群落与低地雨林和山地雨林共有优势种较多,其中柬埔寨子楝树、白颜、白肉榕、大花五桠果、肉实树为低地雨林与中海拔群落的共有优势种。陆均松、海南杨桐、木荷(Schima superba)、五列木为中海拔群落与山地雨林的共有优势种。而鹅掌柴和红鳞蒲桃(Syzygium hancei)为低地雨林和山地雨林共有优势种,在低地雨林和山地雨林内均有分布,表明鹅掌柴和红鳞蒲桃种群适应能力强且生态幅宽广。Plot 1-Plot 6排名前十的优势种重要值之和占比分别为49.04%、44.77%、33.95%、32.6%、40.44%、39.88%。随海拔升高,各群落排名前十优势种重要值之和占比基本呈现先降低后升高格局。中海拔群落优势种重要值占比最低,且优势种重要值相差不大,为多优势种群落。
2.3 物种多样性
如图1所示,Plot 1-Plot 6,Margalef丰富度指数分别为16.085、17.073、24.888、23.192、18.415、18.044。Shannon-Wiener多样性指数分别为3.898、3.883、4.304、4.471、4.180、4.088。Simpson多样性指数分别为0.966、0.963、0.973、0.982、0.975、0.970。Pielou均匀度指数分别为0.839、0.820、0.824、0.883、0.864、0.847。随海拔升高,Margalef丰富度指数、Shannon-Wiener多样性指数、Simpson多样性指数、Pielou均匀度指数基本都呈现先升高后降低的格局。山地雨林Shannon-Wiener多样性指数、Simpson多样性指数、Pielou均匀度指数均高于低地雨林,说明山地雨林群落物种多样性更高且物种分布更均匀。低地雨林中Plot 1 Simpson多样性指数和Pielou均匀度指数高于Plot 2,可能与Plot 1的演替阶段较Plot 2高有关,物种分布更均匀。
对不同海拔样地间的Srensen群落相似度研究显示(表4),群落间Srensen群落相似度和海拔高度差呈显著负相关(Pearson相关系数=-0.66, P<0.01;Spearman秩相关系数=-0.60, P<0.05),故随海拔高度差增加,群落间Srensen群落相似度有降低的趋势。Plot 3和Plot 4、Plot 4和Plot 5、Plot 5和Plot 6,海拔相近的Srensen群落相似度较高,为0.527~0.571;Plot 2和Plot 6、Plot 1和Plot 6、Plot 1和Plot 5,海拔差较大的Srensen群落相似度较低,为0.218~0.252。低地雨林间Srensen群落相似度(0.349)低于山地雨林(0.527),作为低地雨林与山地雨林过渡性质的Plot 3和Plot 4群落相似度最高,为0.571。
2.4 区系组成特征
对吊罗山不同海拔热带天然林群落木本植物属的分布区类型研究显示(表5),吊罗山不同海拔热带天然林均以热带区分布占绝对优势,占比82.1%~94%,温带区分布占比较少,世界分布和中国特有分布极少,具有强烈的热带性质。热带区分布类型中,以热带亚洲(印度-马来西亚)分布为主,占比20.9%~33.9%,包含木荷属(Schima)、润楠属(Machilus)、五列木属(Pentaphylax)、蕈树属(Altingia)、波罗蜜属(Artocarpus)等。其次为泛热带分布,占比19.3%~25.4%,包含鹅掌柴属(Schefflera)、厚壳桂属(Cryptocarya)、山矾属(Symplocos)、柿属(Diospyros)等。之后依次为热带亚洲至热带大洋洲分布,占比11.6%~14.7%,包含山龙眼属(Helicia)等。旧世界热带分布占比9%~13.8%,包含柴龙树属(Apodytes)、蒲桃属(Syzygium)等。热带亚洲和热带美洲间断分布占比4.1%~9%,包含木姜子属(Litsea)等。热带亚洲至热带非洲分布占比较少,为2.9%~6.8%,包含杨桐属(Adinandra)等。表明吊罗山热带天然林区系具有强烈的热带亚洲性质,并具有热带亚洲或印度-马来西亚亲缘。温带区分布类型中,以东亚和北美洲间断分布为主,包含柯属(Lithocarpus)等。各群落地中海区、西亚至中亚分布仅有1属,均为木犀榄属(Olea)。各群落中国特有分布仅有1属,其中Plot 4为半枫荷属(Semiliquidambar),Plot 6为山铜材属(Chunia),其他群落为多核果屬(Pyrenocarpa)。
吊罗山热带天然林热带区分布比例和海拔呈极显著负相关(Pearson相关系数=-0.79, P<0.01;Spearman秩相关系数=-0.76, P<0.01),温带区分布比例和海拔呈极显著正相关(Pearson相关系数=0.72, P<0.01;Spearman秩相关系数=0.70, P<0.01)。故随海拔升高,吊罗山热带天然林属的分布区类型变化呈现热带区分布占比降低、温带区占比升高的特点。
2.5 叶性质
对群落外貌特征-叶性质研究显示(表6),吊罗山热带天然林呈现出以中型叶、单叶、革质叶、全缘叶为主的热带地区典型外貌特征。从叶级来看,吊罗山热带天然林中型叶占样地总物种数的60.00%~64.56%,处于绝对优势;小型叶占比24.56%~36.00%,处于相对优势;大型叶占比3.20%~14.04%;细型叶、微型叶、巨型叶极少。小型叶比例和海拔呈极显著正相关(Pearson相关系数=0.63, P<0.01;Spearman秩相关系数=0.65, P<0.01),随海拔升高,叶级呈现出中型叶、大型叶比例降低,小型叶比例升高的现象。Plot 1、Plot 2地处气温高、水热条件较好的低海拔地区,叶级偏大,而地处1 130 m高海拔地区的Plot 6,气温低,叶级偏小。
从叶型来看,单叶占比83.33%~95.20%,处于绝对优势;复叶占比4.80%~16.67%。单叶比例和海拔呈极显著正相关(Pearson相关系数=0.56, P<0.01;Spearman秩相关系数=0.59, P<0.01),随海拔升高,叶型呈现出复叶比例降低,单叶比例升高的现象。低地雨林水热条件较好,拥有更多大型羽状复叶植物。
从叶质来看,革质叶占比62.28%~75.96%,处于绝对优势,纸质叶占比24.04%~32.46%,处于相对优势,膜质叶很少。革质叶比例和海拔相关性不显著(Pearson相关系数=0.16, P=0.39>0.05;Spearman秩相关系数=0.16, P=0.40>0.05),随海拔升高,叶质变化不明显。
从叶缘来看,全缘叶占比75.32%~85.09%,处于绝对优势,非全缘叶占比14.91%~24.68%。非全缘叶比例和海拔相关性不显著(Pearson相关系数=0.11, P=0.57>0.05;Spearman秩相关系数=0.21, P=0.26>0.05),随海拔升高,叶缘变化不明显。
3 讨论与结论
3.1 物种多样性变化
随海拔升高,温度降低、湿度升高、树木受人为干扰的影响减少,吊罗山热带天然林物种组成以及α物种多样性均呈现先升高后降低的“中间膨胀”现象。与江海声(2006)对吊罗山蕨类植物、卓书辉等(2017)对五指山热带雨林研究具有一致性。海拔较低的低地雨林受人为活动影响大,干扰了植物种子的定居和生长,严重的人为干扰将使得物种多样性骤降(巫翠华等,2021)。海拔较高的山地雨林温度低,影响了物种的生存和分布。而中海拔群落水热条件最为适宜、中等干扰使群落树种竞争排斥作用减弱,有助于维持较高的物种多样性(龙文兴,2016;贾真真等,2021); 且Plot 3和Plot 4是低地雨林向山地雨林的过渡类型,交汇区生境异质性高, 物种多样性大幅提升(王宇超和王得祥,2013)。吴裕鹏等(2013)研究发现尖峰岭原始林形成特殊的倒“S”型垂直分布格局。尖峰岭地处海南岛西部,高温少雨且土壤贫瘠,低海拔仅适合旱生植物生存,中低海拔又为龙脑香科(Dipterocarpaceae)优势群落,故物种多样性先降低后升高。而吊罗山地处海南岛东部沿海地区,水热条件优良、土壤肥沃,中低海拔群落物种多样性随人为干扰程度降低而升高,体现出树木与其生境相适应的特点。
不同树种生态幅各不相同,随海拔升高,优势种具有明显的更替现象,随海拔高度差增加,群落间Srensen群落相似度降低。低地雨林位于较低海拔,地貌结构和水热条件都与海拔较高的山地雨林存在较大差异,从而造成了物种分布的差异性。低地雨林间共有优势种不明显,Srensen群落相似度较低,且群落单种科和单种属数量丰富(王帅等,2015),可能是20世纪50年代的森林砍伐(李意德,1995)和频繁的人为干扰使具有肥沃土壤的吊罗山低地雨林有更多新物种入驻,形成低地混合雨林,而非演替为耐贫瘠、耐干旱的单优龙脑香科低地雨林(黄瑾等,2013)。而山地雨林均以陆均松为优势,且Srensen群落相似度较高,表明吊罗山山地雨林保护较好,已演替至后期阶段。
3.2 区系组成变化
吊罗山不同海拔热带天然林属的分布区类型均以热带区分布占绝对优势,且以热带亚洲(印度-马来西亚)分布为主。表明区系组成含有强烈的热带亚洲性质,并具有热带亚洲或印度-马来西亚亲缘。其温带区分布以东亚和北美洲间断分布为主,而霸王岭以壳斗科为优势的山地雨林以北温带分布为主(吕晓波,2012),反映了不同地区物种起源和迁移历史的差异性。随海拔升高,温度降低,植物区系呈现热带性减弱、温带性加强的特点(苏文苹,2007;朱华,2008;巫翠华等,2021),与区系组成随纬度变化规律相同(吴征镒等,2006)。除纬度因素和海拔因素外,过度的人为干扰会导致狭域分布种、特征种减少,入侵种、广布种增多(程占红等, 2014),使植物区系成分趋于单一。而复杂、特殊的生境有利于特有种、珍稀树种和起源古老的孑遗植物存活,中国特有分布丰富(魏新增等,2009)。一般来说,随海拔升高,人为干扰减少,中国特有分布将增加(苏文苹,2007),植物区系地理成分更加复杂(巫翠华等,2021)。而吊罗山不同海拔热带天然林中国特有属分布均很少,表明物种特有性较弱,可能是20世纪50年代的森林砍伐(李意德,1995),影响了生态位狭窄的特有种存活。
3.3 叶性质变化
叶性质是植物对自然生境长久适应的结果,具有深刻的生境指示作用。随纬度降低,温度與湿度增加,落叶树种减少而喜温暖湿润的常绿树种增多,植物叶性质呈现小型叶、复叶、草质叶、非全缘叶比例降低,中型叶、单叶、革质叶、全缘叶比例升高的现象(孔祥海等,2009)。吊罗山热带天然林水热条件优良,外貌结构呈现以中型叶、单叶、革质叶、全缘叶为主的热带地区显著特点(王伯荪,1987)。热带森林植株茂密,林冠遮挡了阳光和部分降雨,较大叶片有助于获取更多阳光和蒸发水分降低叶片温度。革质叶通过反射剧烈阳光而降低蒸腾作用(苏文苹,2007),以适应热带地区的高温气候。与以壳斗科为优势的霸王岭山地雨林相比(吕晓波,2012),吊罗山热带天然林叶级偏小,复叶比例、纸质叶比例偏低。这与霸王岭样地地处沟谷附近气候高温且潮湿有关,有利于巨型叶植物生长;且霸王岭样地以壳斗科植物为优势并增加了对草本植物及蕨类的调查,故复叶比例和纸质叶比例相对较高。从温暖潮湿地区到干燥或寒冷地区叶级呈现由大变小的趋势(王伯荪,1987),随海拔升高,温度降低湿度增加,叶级变化与低纬度地区至高纬度地区叶性质变化相似(刘兴良等,2013),而叶型变化相反。可能的原因是低地雨林地处沟谷附近,气候同赤道雨林相似,温暖且潮湿,拥有更多大型羽状复叶树种。而高海拔地区山风强烈且气温低的特殊生境,使叶面积减小。因此,受水热、光照、历史起源等因素的综合影响,植物叶性质体现出植物与生境相适应的外貌特征。
参考文献:
AN SQ, WANG ZF, ZENG FJ, et al., 1999. Biodiversity of tropical mountane rain forest on Diaoluo Mountain, Hainan [J]. Acta Sci Nat Univ Sunyatseni, 38(6): 79-84. [安树青, 王峥峰, 曾繁敬, 等, 1999. 海南吊罗山热带山地雨林植物种类多样性研究 [J]. 中山大学学报(自然科学版), 38(6): 79-84.]
BROWN JH, 2001. Mammals on mountainsides: elevational patterns of diversity [J]. Glob Ecol Biogeogr, 10(1): 101-109.
CHENG ZH, NIU LQ, HU YQ, 2014. Ecological change of species of wetland plant with human disturbance in scenic area of Wutai mountains [J]. Wetland Sci, 12(1): 89-96. [程占红, 牛莉芹, 胡亚晴, 2014. 五台山风景区人为干扰下湿地植物物种的生态变化 [J]. 湿地科学, 12(1): 89-96.]
DING T, LIAO WB, JIN JH, et al., 2002. Floristic analysis on the seed plants of Mt. Diaoluo in Hainan Island [J]. Guihaia, 22(4): 311-326. [丁坦, 廖文波, 金建华, 等, 2002. 海南岛吊罗山种子植物区系分析 [J]. 广西植物, 22(4): 311-326.]
GASTON KJ, 2000. Global patterns in biodiversity [J]. Nature, 405(6783): 220-227.
GHALEY BB, PORTER JR, 2014. Ecosystem function and service quantification and valuation in a conventional winter wheat production system with DAISY model in Denmark [J]. Ecosyst Serv, 10: 79-83.
HAN TY, SHEN Y, WANG X, et al., 2019. Community characteristics of a lowland rainforest in Diaoluoshan in Hainan Province [J]. For Environ Sci, 35(3): 43-49. [韩天宇, 沈燕, 王旭, 等, 2019. 海南吊罗山低地雨林群落特征分析 [J]. 林业与环境科学, 35(3): 43-49.]
HE JS, CHEN WL, 1997. A review of gradient changes in species diversity of land plant communities [J]. Acta Ecol Sin, 17(1): 93-101. [贺金生, 陈伟烈, 1997. 陆地植物群落物种多样性的梯度变化特征 [J]. 生态学报, 17(1): 93-101.]
HUANG J, YANG XB, LONG WX, et al., 2013. Characteristics of forest communities solely dominated by Dipterocarpaceae plants in Hainan Island [J]. Chin J Trop Crops, 34(3): 578-583. [黄瑾, 杨小波, 龙文兴, 等, 2013. 海南单优龙脑香科植物群落特征 [J]. 热带作物学报, 34(3): 578-583.]
JIA ZZ, LI WJ, TIAN A, et al., 2021. Ecological influence of tourism disturbance on the characteristics of different communities in Baili Rhododendron forest [J]. Acta Ecol Sin, 41(11): 4641-4649. [贾真真, 李苇洁, 田奥, 等, 2021. 贵州百里杜鹃风景区旅游干扰对杜鹃群落特征的影响 [J]. 生态学报, 41(11): 4641-4649.]
JIANG HS, 2006. The biodiversity and its conservation of Diaoluo mountain, Hainan, China [M]. Guangzhou: Guangdong Science & Technology Press: 1-71. [江海声, 2006. 海南吊罗山生物多样性及其保护 [M]. 广州: 广东科学技术出版社: 1-71.]
KONG XH, HUANG SH, CHEN XH, et al., 2009. Characteristics analysis of the evergreen broad-leaved forest in Minxi, Fujian Province [J]. J Fujian Coll For, 29(4): 351-356. [孔祥海, 黃素华, 陈小红, 等, 2009. 闽西常绿阔叶林群落特征分析 [J]. 福建林学院学报, 29(4): 351-356.]
LAKHANI KH, MAGURRAN AE, 1989. Ecological diversity and its measurement [J]. J Appl Ecol, 26(3): 1101.
LEGENDRE P, MI XC, REN HB, et al., 2009. Partitioning beta diversity in a subtropical broad-leaved forest of China [J]. Ecology, 90(3): 663-674.
LI YD, 1995. Changes of tropical forest and protection countermeasures of biodiversity in Hainan Island [J]. For Res, 8(4): 455-461. [李意德, 1995. 海南岛热带森林的变迁及生物多样性的保护对策 [J]. 林业科学研究, 8(4): 455-461.]
LIU XL, HE F, FAN H, et al., 2013. Leaf-form characteristics of plants in Quercus aquifolioides community along an elevational gradient on the Balang Mountain in Wolong Nature Reserve, Sichuan, China [J]. Acta Ecol Sin, 33(22): 7148-7156. [刘兴良, 何飞, 樊华, 等, 2013. 卧龙巴郎山川滇高山栎群落植物叶特性海拔梯度特征 [J]. 生态学报, 33(22): 7148-7156.]
LONG WX, 2016. Plant ecology [M]. Beijing: Science Press: 84-185. [龙文兴, 2016. 植物生态学 [M]. 北京: 科学出版社: 84-185.]
L XB, 2012. Study on the community structure and species diversity of Fagaceae dominance in montane rain forest in Bawangling, Hainan Island [D]. Haikou: Hainan University: 34-38. [吕晓波, 2012. 海南霸王岭以壳斗科植物为优势的山地雨林群落结构及物种多样性研究 [D]. 海口: 海南大学: 34-38.]
SIMSIMM, LOPES T, RUAS S, et al., 2015. Does altitude shape molecular diversity and richness of bryophytes in Madeiras natural forest? A case study with four bryophyte species at two altitudinal levels [J]. Plant Ecol Evol, 148(2): 171-180.
SU WP, 2007. Comparison of mid-montane humit evergreen broadleaved forest community between eastern and western slope of southern Gaoligong mountains [D]. Kunming: Southwest Forestry University: 77-78. [苏文苹, 2007. 高黎贡山南段东西坡中山湿性常绿阔叶林群落学比较研究 [D]. 昆明: 西南林学院: 77-78.]
TANG ZR, FANG JY, 2004. A review on the elevational patterns of plant species diversity [J]. Biodivers Sci, 12(1): 20-28. [唐志尧, 方精云, 2004. 植物物种多样性的垂直分布格局 [J]. 生物多样性, 12(1): 20-28.]
WANG BS, 1987. Phytocoenology [M]. Beijing: Higher Education Press: 56-60. [王伯荪, 1987. 植物群落学 [M]. 北京: 高等教育出版社: 56-60.]
WANG S, LI JL, WANG X, et al., 2015. Tree species diversity of secondary tropical lowland rainforest in Diaoluoshan in Hainan Province [J]. Chin J Trop Crops, 36(5): 998-1005. [王帅, 李佳灵, 王旭, 等, 2015. 海南吊罗山热带低地雨林次生林乔木物种多样性研究 [J]. 热带作物学报, 36(5): 998-1005.]
WANG YC, WANG DX, 2013. Study on elevational patterns of plant species diversity and community complexity in Foping Natural Reserve [J]. Acta Bot Boreal-Occident Sin, 33(1): 169-176. [王宇超, 王得祥, 2013. 佛坪自然保護区植物群落物种多样性和复杂性的海拔格局研究 [J]. 西北植物学报, 33(1): 169-176.]
WANG ZF, AN SQ, DAVID G, et al., 1999. Biodiversity of the montane rain forest in Diaoluo mountain, Hainan [J]. Acta Ecol Sin, 19(1): 63-69. [王峥峰, 安树青, DAVID G, 等, 1999. 海南岛吊罗山山地雨林物种多样性 [J]. 生态学报, 19(1): 63-69.]
WEI XZ, HE D, JIANG MX, et al., 2009. Characteristics of riparian rare plant communities on the Shennongjia mountains, Central China [J]. J Wuhan Bot Res, 27(6): 607-616. [魏新增, 何东, 江明喜, 等, 2009. 神农架山地河岸带中珍稀植物群落特征 [J]. 武汉植物学研究, 27(6): 607-616.]
WU CH, ZHANG LL, QIAO WG, et al., 2021. Vertical distribution pattern and impact factor of species diversity in Zibai Mountain Nature Reserve [J]. J Fujian Agric For Univ(Nat Sci Ed), 50(5): 630-635. [巫翠华, 张利利, 乔卫国, 等, 2021. 紫柏山自然保护区植物多样性垂直分布格局及其影响因素 [J]. 福建农林大学学报(自然科学版), 50(5): 630-635.]
WU YP, XU H, LI YD, et al., 2013. Elevation patterns of tree and shrub species diversity of tropical forests in Jianfengling, Hainan Island [J]. Sci Silv Sin, 49(4): 16-23. [吴裕鹏, 许涵, 李意德, 等, 2013. 海南尖峰岭热带林乔灌木层物种多样性沿海拔梯度分布格局 [J]. 林业科学, 49(4): 16-23.]
WU ZY, ZHOU ZK, SUN H, et al., 2006. The areal-types of seed plants and their origin and differentiation [M]. Kunming: Yunnan Science and Technology Press: 1-451. [吳征镒, 周浙昆, 孙航, 等, 2006. 种子植物分布区类型及其起源和分化 [M]. 昆明: 云南科技出版社: 1-451.]
YANG XB, 2016. Checklist of plant species in Hainan (Vol. 1-14) [M]. Beijing: Science Press. [杨小波, 2016. 海南植物图志(1-14卷) [M]. 北京: 科学出版社.]
ZHU H, 2008. Distribution patterns of genera of Yunnan seed plants with references to their biogeographical significances [J]. Adv Earth Sci, 8(8): 830-839. [朱华, 2008. 云南种子植物区系地理成分分布格局及其意义 [J]. 地球科学进展, 8(8): 830-839.]
ZHUO SH, HU N, CHEN K, et al., 2017. Species distribution pattern of tree plant communities on different slopes and altitudes in the Wuzhishan Nature Reserve [J]. J Trop Biol, 8(4): 436-443. [卓书辉, 胡能, 陈康, 等, 2017. 五指山自然保护区不同坡向和海拔的乔木群落物种分布格局 [J]. 热带生物学报, 8(4): 436-443.]
(责任编辑 周翠鸣)
