亚洲栽培稻5 种不同生态类型与籼粳间的杂种F1 育性研究
2023-06-14周家武浦秋红吕永刚邓先能黄望启陶大云
王 杰,周家武,李 静,杨 莺,浦秋红,吕永刚,邓先能,黄望启,王 敏,张 玉**,陶大云**
(1.云南大学 资源植物研究院,云南 昆明 650500;2.云南省稻作遗传改良重点实验室,云南省农业科学院粮食作物研究所,云南 昆明 650200)
亚洲栽培稻(Oryza sativa)是世界上最重要的粮食作物之一,提供了全球近50%人口的主食来源,水稻增产对保障中国乃至世界的粮食安全和人民生活水平具有极其重要的作用[1].传统上,亚洲栽培稻被划分为籼稻和粳稻2 个亚种[2].这2 个亚种在形态特征、生理特性和遗传基础上表现出明显的差异[3].随着生物技术的发展和新的分子标记在亚洲栽培稻遗传多样性检测上的应用,传统的亚洲栽培稻的籼稻和粳稻分类得到了证实[4-11].亚洲栽培稻因适应不同地理、生态、耕作栽培方式而具有丰富的变异,普遍被划分成5 个生态型:Aus稻(Aus)、芳香稻(aromatic)、籼稻(indica)、热带粳稻(tropicaljaponica)和温带粳稻(temperatejaponica)[12-15].亚种间遗传距离较大,具有较强的杂种优势,是最有希望且能在较短时期内实现产量突破的途径[16].由于亚种间杂种F1的花粉育性或(和)小穗育性较低,表现为不育或半不育,严重阻碍了亚种间杂种优势的利用.Kato 等[2,17]研究表明亚洲栽培稻亚种间杂种F1代育性变化范围很大,从完全不育到完全可育,且亚种间杂种育性显著低于亚种内杂种的育性,这一结论也得到了后续研究的支持[3,18-19].以杂种不育为主的生殖隔离是造成亚种间杂种育性低的主要原因.近年来,随着分子标记技术的发展和广泛应用,已有超过50 个水稻杂种不育基因被定位,其中有10 个已经被克隆[20].一系列研究表明,不同杂交组合含有不同的杂种不育位点.例如Sa[21]、S5[22]、Sc[23]、HSA1[24]是在温带粳和籼稻杂交组合中检测到的杂种不育基因座;DPL1和DPL2[25]等是在温带粳和Aus稻杂交组合中检测到的杂种不育基因座;S7位点则为Aus稻和籼稻/粳稻杂交组合中发现的控制杂种雌配子育性的基因座[26].
生殖隔离尤其是杂种不育是生物种/群形成的重要指标,一直以来是生命科学领域的核心问题之一.目前杂种不育研究主要基于亚洲栽培稻籼、粳2 个亚种的少数组合之间,证实了籼、粳稻杂种不育的普遍性.基于分子标记、基因/基因组序列把亚洲栽培稻划分为5 种生态类群/型,但缺乏生殖隔离的研究结果来验证、支撑.本研究基于基因组研究成就,从生产实践出发,既考虑到亚洲栽培稻以籼、粳为主,又考虑到亚洲栽培稻种内丰富的变异,选用国际水稻研究所筛选的19 份代表5 种不同生态类型的亚洲栽培稻作为父本,云南温带粳品种滇粳优1 号及籼稻品种IR64 为母本,对杂种F1育性进行了系统全面的研究,为认识亚洲栽培稻分化分类和利用杂种优势提供参考.
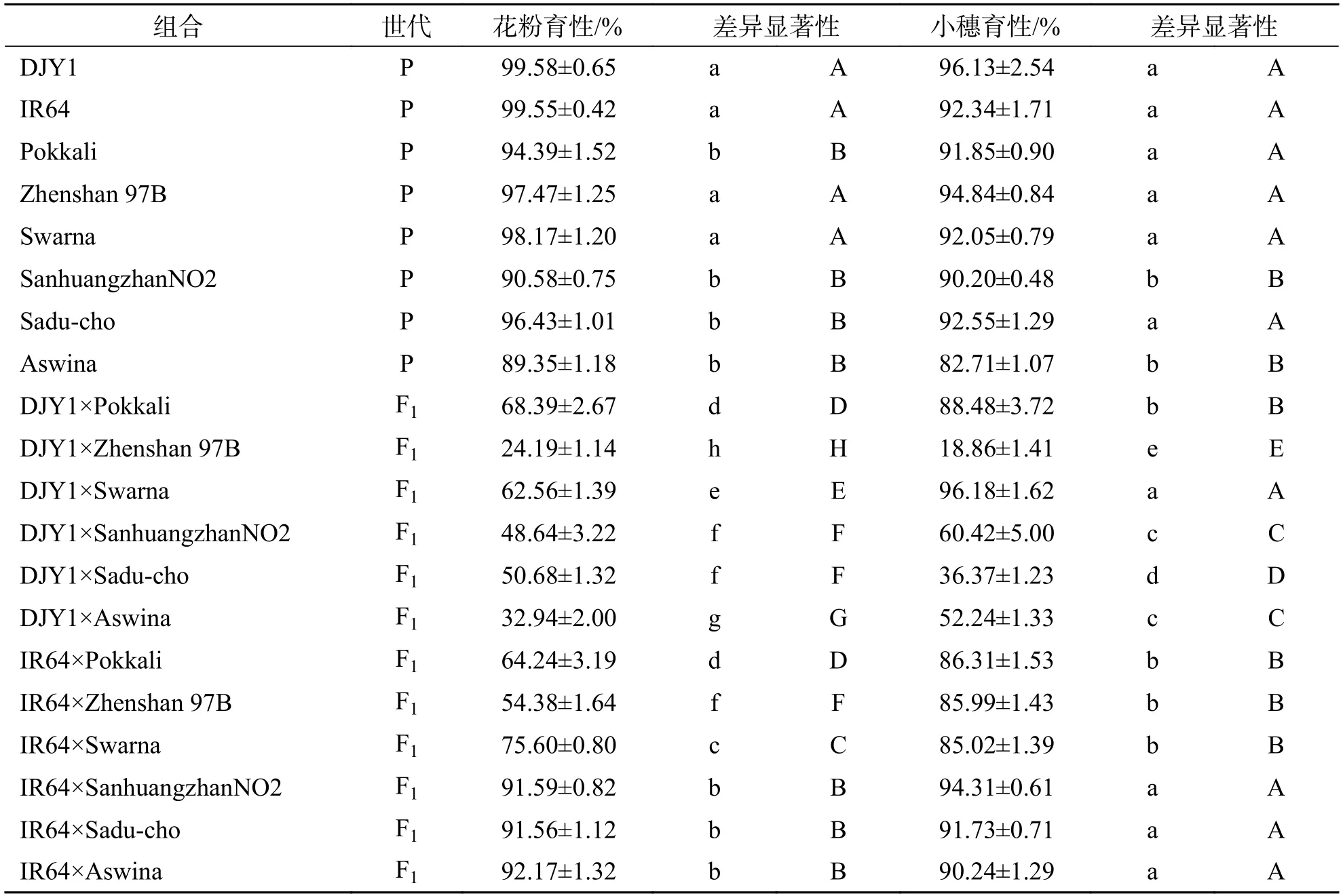
表5 籼稻分别与滇粳优1 号和IR64 杂交F1 花粉育性和小穗育性Tab.5 Pollen and spikelet fertility of F1 hybrids of japonica rice and Dianjingyu 1 and IR64,respectively
1 材料与方法
1.1 材料以云南温带粳稻滇粳优1 号、国际水稻所选育的籼稻品种IR64 为母本,以国际水稻研究所(IRRI)筛选的19 份代表5 种不同生态类型的亚洲栽培稻为父本[13],分别进行杂交(表1),配制F1组合.
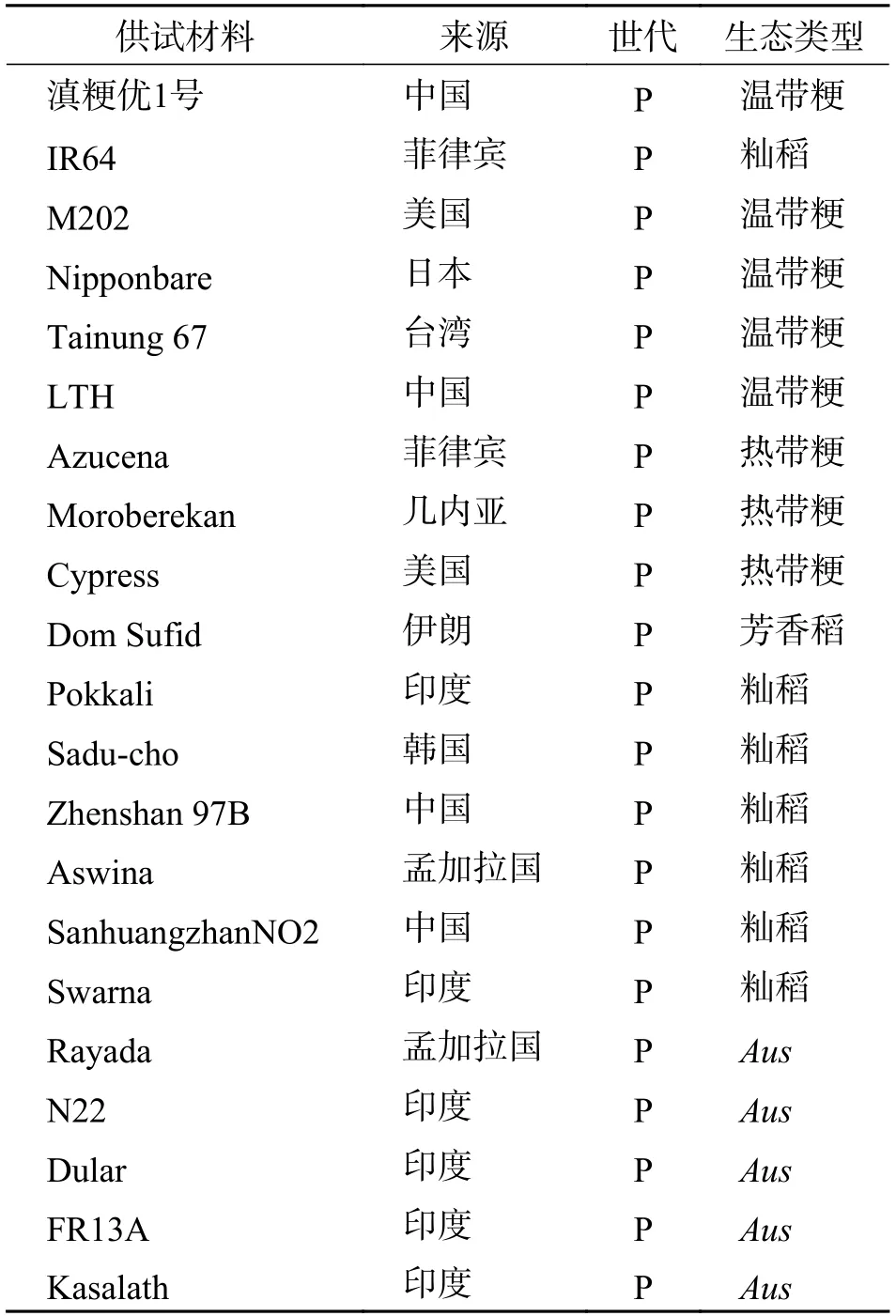
表1 供试材料Tab.1 Test materials
1.2 方法试验于2020 年在云南省农业科学院粮食作物研究所景洪试验基地进行.21 份供试材料于早稻(1-6 月)种植,用于配制F1杂交组合;晚稻(6-11 月)种植亲本及38 个F1杂交组合各10 株,常规栽培管理,调查花粉育性和小穗育性.
花粉育性调查:从单株主穗中上部的枝梗中,采集 5~10 朵发育成熟、于当天或次日即将开花的小花(枝梗末端的颖花),在70%的乙醇中固定.用1% I2-KI 溶液染色,在160×普通光学显微镜下观察.根据花粉染色及形态判断其育性,即大小正常、圆形黑褐色的花粉为可育,形状不规则、黄色及不染色、或者染色但较小的花粉粒为不育.每份材料观察3 个视野,且每个视野不少于300 粒花粉.花粉育性为可育花粉占总花粉粒数的比率.
小穗育性调查:植株成熟后,于每株3 个穗子中上部考察小穗受精粒数.小穗育性为受精粒数占总粒数的百分比.
2 结果与分析
2.1 温带粳分别与滇粳优1 号和IR64 杂交F1 育性分析由表2 可知,以滇粳优1 号为母本,分别与4 个温带粳品种配组的杂交组合育性均表现正常.与父本比较,4 个杂交F1组合花粉育性均显著性提高,小穗育性也有一定程度的升高.值得注意的是,与Nipponbare 相比,DJY1×Nipponbare 组合小穗育性由原来的74.82%升高到90.56%,差异极显著.这种情况可能由于Nipponbare 是典型的长日照品种,适合在中国北方种植,而在云南景洪短日照自然条件下,表现为株高变矮,抽穗期提前且小穗育性下降.当与云南品种滇粳优1 号杂交后,对光周期的敏感性降低,育性正常.以IR64 为母本,4 个温带粳品种为父本配制的组合育性差异较大.小穗育性变幅为19.27%~55.67%,花粉育性变幅为27.68%~70.59%.各F1杂交组合花粉育性和小穗育性情况也不相同.IR64×M202 组合表现为花粉育性可育,小穗育性半不育;IR64×Tainung 67 组合表现为花粉育性高不育,小穗育性半不育;IR64×Nipponbare、IR64×LTH 组合则均表现为花粉育性低不育,小穗育性高不育,但两个组合育性均存在显著性差异.
在父本相同的情况下,即同一温带粳品种在籼、粳不同背景下进行配组,育性存在明显的差异,滇粳优1 号×温带粳组合育性显著高于与IR64×温带粳组合.
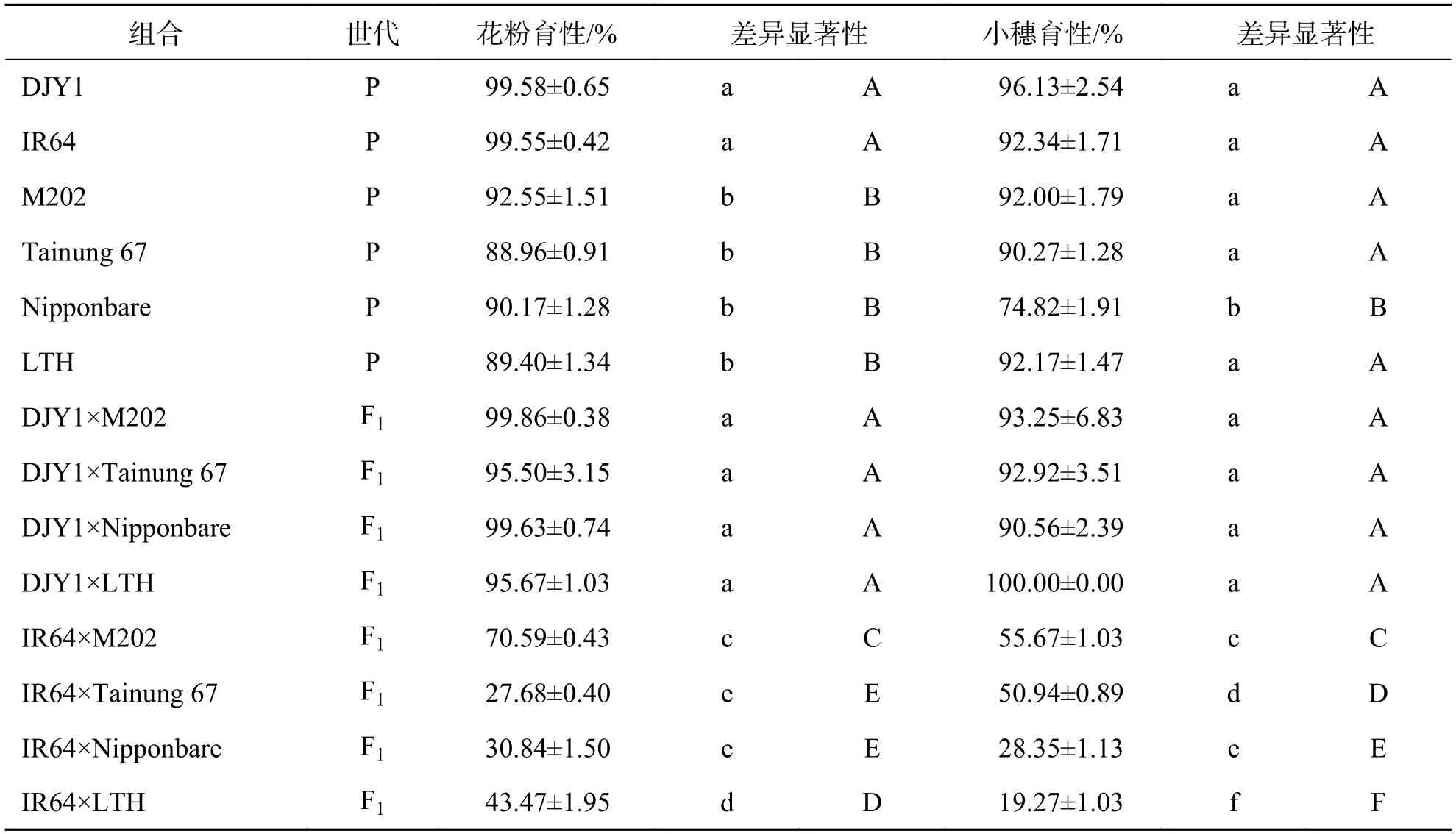
表2 温带粳分别与滇粳优1 号和IR64 杂交F1 花粉育性和小穗育性Tab.2 Fertility of F1 pollen and spikelet of temperate japonica hybridized with Dianjingyou 1 and IR64,respectively
由此看出,温带粳与同属于温带粳的滇粳优1号之间遗传距离较小,杂种育性正常.温带粳与籼稻IR64 之间普遍存在杂种不育,F1代育性变化范围很大,从高不育到正常可育.在母本同为IR64 的情况下,对花粉育性影响依次为:Tainung 67>Nipponbare> LTH> M202,暗示IR64×Tainung 67 组合含有更多控制杂种花粉不育的基因.在4 个不同组合中,对小穗育性的影响依次为:LTH> Nipponbare> Tainung 67>M202,表明IR64×LTH 组合中可能含有更多的控制小穗不育的基因.综上所述,温带粳之间不存在杂种不育现象,而温带粳与籼稻普遍存在杂种不育现象,且杂种不育程度与双亲背景相关.
2.2 热带粳分别与滇粳优1 号和IR64 杂交F1 育性分析由表3 可知,滇粳优1 号与不同热带粳品种杂交的3 个F1组合育性普遍降低,且不育类型和不育程度各不相同.DJY1×Azucena 组合花粉和小穗均为高不育;DJY1×Moroberekan 组合花粉半不育,小穗正常可育;DJY1×Cypress 组合花粉为高不育,小穗为半不育.此外,3 个热带粳品种与IR64 杂交组合育性表现,和与滇粳优1 号杂交组合表现不同.IR64×Azucena 组合育性均在80%以上,表现为花粉育性高于小穗育性.IR64×Cypress组合育性则均在85%以上,表现为花粉育性低于小穗育性.IR64×Moroberekan 组合花粉育性表现为半不育,小穗育性相比于父本Moroberekan(85.16%)升高显著,为90.23%.
从表3 中可以发现,热带粳Azucena、Cypress与籼稻杂交F1育性显著高于与粳稻杂交F1育性,表明这2 个品种与温带粳的育性障碍比与籼稻的育性障碍更为严重.热带粳Moroberekan 无论是与滇粳优1 号还是与IR64 杂交,花粉育性均为半不育,而小穗育性均在90%以上,说明热带粳Moroberekan 与滇粳优1 号和IR64 杂交只影响雄配子育性.
综上所述,温带粳滇粳优1 号与3 个热带粳品种间普遍存在育性障碍,IR64 与所用的热带粳杂交,育性普遍正常.这为克服杂种不育利用强杂种优势提供思路和材料,但原因需要进一步的研究.
2.3 芳香稻分别与滇粳优1 号和IR64 杂交F1 育性分析由表4 可知,芳香稻Dom sufid 分别与滇粳优1 号和IR64 杂交F1育性均不正常,且IR64×Dom Sufid 组合育性显著高于DJY1×Dom Sufid.DJY1×DOM Sufid 组合花粉育性表现为高不育,为10.61%,小穗育性为34.93%.IR64×Dom Sufid 组合花粉育性为43.08%,小穗育性表现为半不育,为53.09%,说明与IR64 相比,Dom Sufid 与DJY1 的育性障碍更为严重,2 个杂交F1组合均为小穗育性高于花粉育性.以上结果表明,芳香稻与滇粳优1号和IR64 均存在育性障碍.
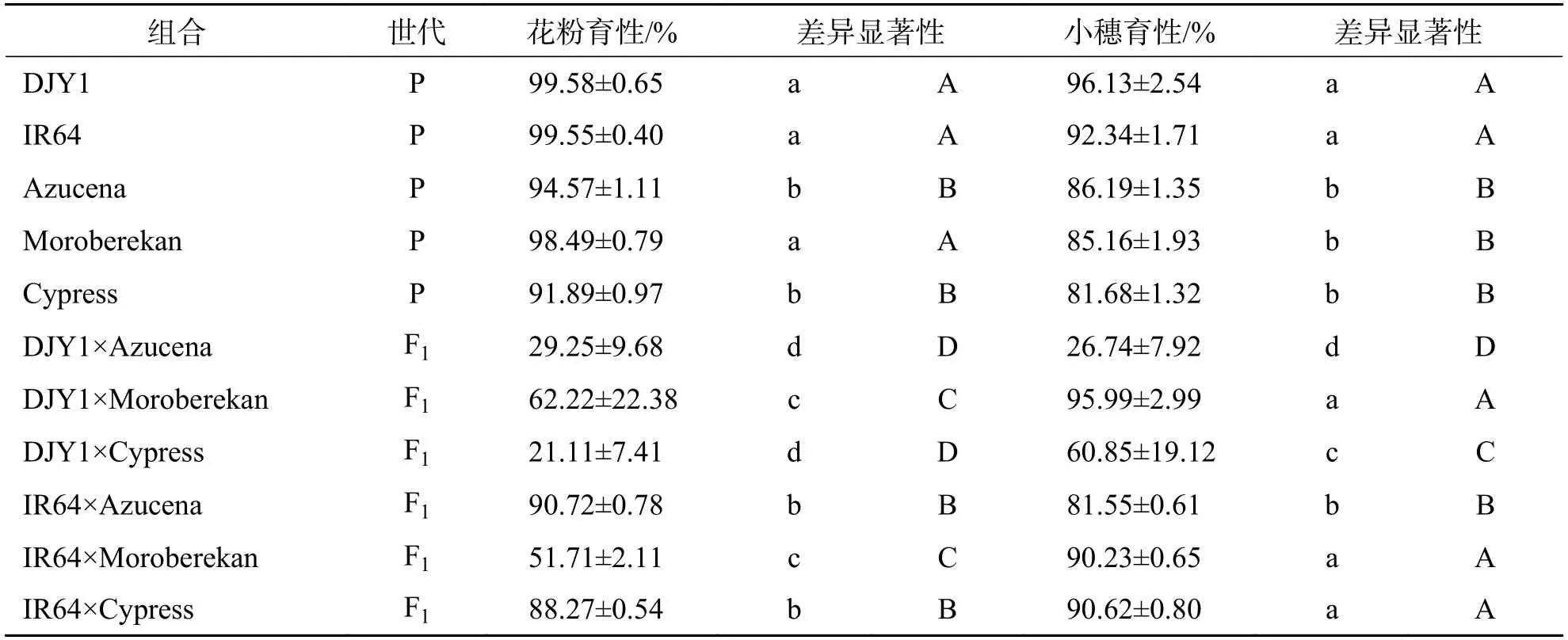
表3 热带粳分别与滇粳优1 号和IR64 杂交F1 花粉育性和小穗育性Tab.3 Pollen fertility and spikelet fertility of hybrid F1 of tropical japonica hybridized with Dianjingou 1 and IR64,respectively
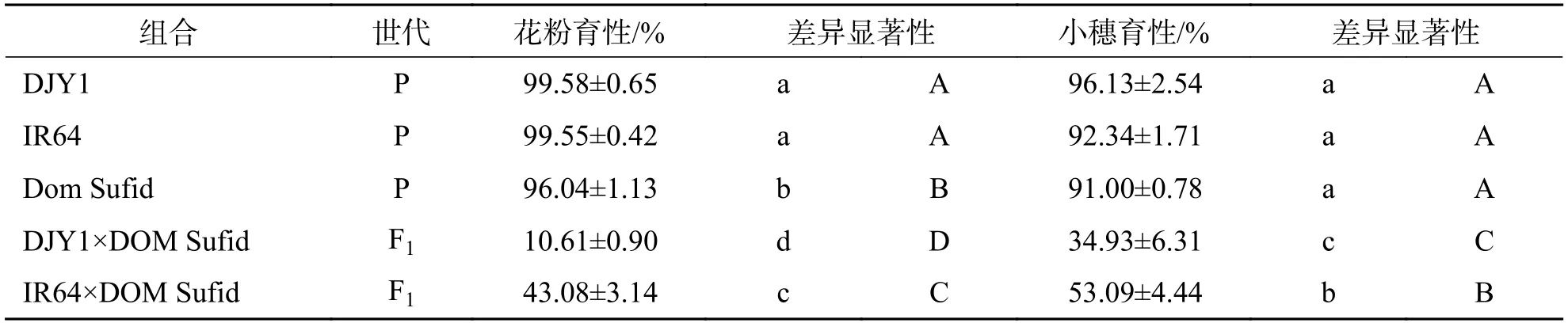
表4 芳香稻分别与滇粳优1 号和IR64 杂交F1 花粉育性和小穗育性Tab.4 Pollen fertility and spikelet fertility of hybrid F1 of aromatic rice and Dianjingyu 1 and IR64,respectively
2.4 籼稻分别与滇粳优1 号和IR64 杂交F1 育性分析由表5 可知,不同籼稻品种与滇粳优1 号普遍存在育性障碍.不同F1组合育性变化范围很大,从完全不育到完全可育,花粉育性变幅在24.10%~68.39%,小穗育性则变幅于18.86%~96.18%.每个组合的小穗育性和花粉育性表现不一致,可以大致分为2 类.①花粉不育,小穗完全可育:DJY1×Pokkali、DJY1×Swarna;②花粉和小穗均不育:DJY1×Zhenshan 97B、DJY1×Sanhuangzhan NO2、DJY1×Sadu-cho、DJY1×Aswina.不同组合花粉和小穗育性不育的程度也各不相同,其中DJY1×Zhenshan 97B 组合育性均为高不育,DJY1×Sanhuangzhan NO2 组合育性均为半不育,DJY1×Sadu-cho 组合花粉育性表现为半不育,小穗育性为低不育,DJY1×Aswina 组合花粉育性为低不育,小穗育性表现为半不育.不同籼稻品种与IR64 杂交F1组合中,大致可以分为3 类.①花粉半不育,小穗完全可育:IR64×Zhenshan 97B;②花粉育性介于60%~80%,小穗完全可育:IR64×Pokkali、IR64×Swarna;③花粉和小穗均完全可育:IR64×Sanhuangzhan NO2、IR64×Sadu-cho、IR64×Aswina.
此外,我们发现,Zhenshan 97B、Sanhuangzhan NO2、Sadu-cho、Aswina 四个品种与籼稻IR64 杂交育性均显著高于与温带粳滇粳优1 号杂交组合育性,说明这4 个籼稻品种与籼稻IR64 的亲和性更强.Pokkali 无论是与IR64 还是滇粳优1 号杂交,育性无显著差异.对于籼稻品种Swarna,IR64×Swarna 组合小穗育性显著低于DJY1×Swarna 组合,IR64×Swarna 组合花粉育性显著高于DJY1×Swarna组合,说明Swarna 与籼稻IR64 的雄配子亲和力较高,而与粳稻滇粳优1 号的雌配子亲和力较高.
由此看出,滇粳优1 号属于温带粳,和籼稻之间遗传距离较大,普遍存在育性障碍.结合表2,表明亚洲栽培稻不同类型中籼粳亚种间普遍存在杂种不育,且亚种间杂种育性显著低于亚种内杂种的育性.此外,不同的籼稻品种之间也存在一定程度上的杂种不育现象.
2.5 Aus 稻分别与滇粳优1 号和IR64 杂交F1 育性分析由表6 可知,在母本同为滇粳优1 号的情况下,与不同Aus品种配组杂交F1的组合中,花粉育性变幅19.12%~82.84%,小穗育性变幅32.12%~90.00%,每个组合小穗育性均高于花粉育性.其中,DJY1×Dular 组合花粉育性和小穗均为完全可育,DJY1×Rayada、DJY1×N22、DJY1×FR13A 三个组合的育性存在显著性差异并依次升高,DJY1×Kasalath 组合小穗育性为90.00%,完全可育,而花粉育性只有45.00%,说明DJY1×Kasalath 杂交组合只影响花粉育性,不影响小穗育性.Aus稻与IR64杂交F1组合中,IR64×Rayada 和IR64×Dular 组合育性均为完全可育;IR64×N22 组合花粉育性为低不育,小穗育性正常可育,为69.99%;IR64×FR13A组合花粉育性为60.48%,小穗育性为46.51%;IR64×Kasalath 组合花粉育性表现为半不育,小穗育性则为高不育,只有20.00%.
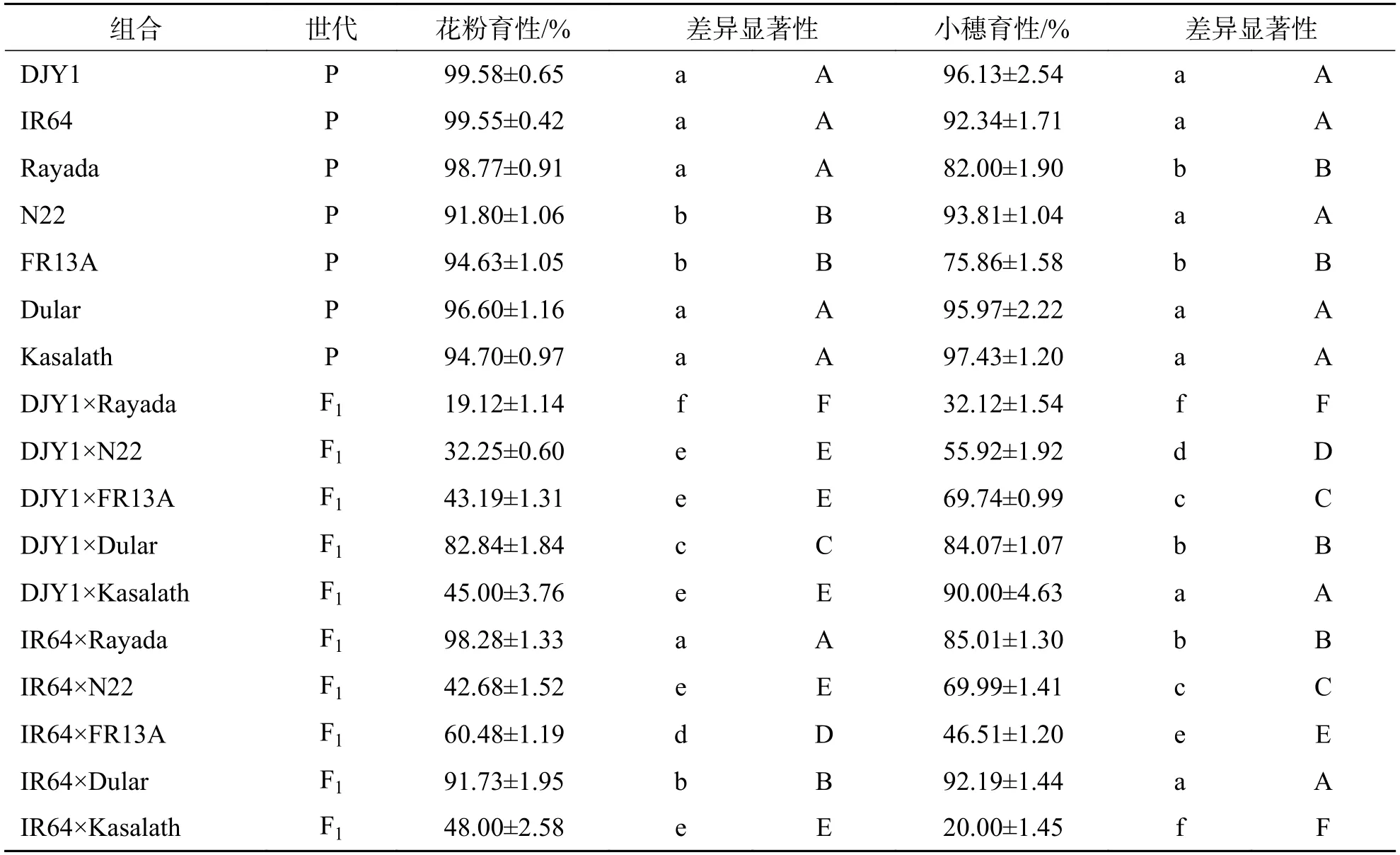
表6 Aus 稻分别与滇粳优1 号和IR64 杂交F1 花粉育性和小穗育性Tab.6 Pollen and spikelet fertility of F1 hybrids of Aus rice and Dianjingyu 1 and IR64,respectively
在研究同一品种分别与粳稻滇粳优1 号和籼稻IR64 杂交F1的表现时发现,Rayada 与IR64 杂交F1花粉育性和小穗育性均完全可育,而与滇粳优1 号杂交则存在明显的育性障碍,说明Rayada与IR64 的亲和性更高.N22 与IR64、滇粳优1 号都存在育性障碍,且DJY1×N22 组合育性显著低于IR64×N22.DJY1×FR13A 组合小穗育性显著低于IR64×FR13A 组合,花粉育性则显著高于IR64×FR13A 组合.Dular 具有广亲和性,其杂种F1均表现为完全可育.Kasalath 为父本分别与滇粳优1 号和IR64 配组的F1组合花粉育性无显著差异,均在45%左右,表现为花粉半不育.DJY1×Kasalath 组合小穗育性为90.00%,IR64×Kasalath 组合小穗育性仅有20.00%,存在极显著性差异,说明Kasalath 与IR64 的杂交组合影响杂种雌配子育性.
综上所述,不同Aus稻品种分别与滇粳优1 号和IR64 杂交,不同组合间育性存在显著差异.除Dular 与籼、粳亚种材料杂交育性正常外,其它Aus品种与籼、粳亚种材料杂交均存在不同程度的育性障碍.
3 讨论
传统上亚洲栽培稻被分为籼稻和粳稻两个亚种[27-29],而新的分类系统普遍把亚洲栽培稻分为籼稻、Aus、芳香稻、温带粳稻和热带粳稻5 种生态类型,大量基于基因组层面的研究也支持这一观点[12-15].生殖隔离尤其是杂种不育是生物种/群形成的重要指标,而对这5 种生态群间杂种不育的研究证据相对缺乏.温带粳品种与同属于温带粳的滇粳优1 号配组F1组合时,育性均表现正常.温带粳品种与籼稻IR64 之间遗传距离较远,存在育性障碍,F1育性普遍不正常.以上现象说明亚种间杂种育性显著低于亚种内杂种的育性.本研究中,IR64×Zhenshan 97B、IR64×Pokkali、IR64×Swarna 组合花粉育性分别为64.24%、54.38%、75.6%,与亲本相比,育性显著降低,表明籼稻品种间也存在一定的育性障碍,这与前人的结果一致[3].有趣的是,热带粳品种与温带粳滇粳优1 号存在育性障碍,支持温带粳与热带粳属于不同亚群的分类.然而本实验所用的热带粳品种与IR64 配组的F1育性普遍正常,这又与5 种生态类型的分类系统不一致.造成这种结果的原因可能是热带粳和籼稻主要分布于东南亚的热带地区[30].一方面,基于基因组研究,相比于温带粳,籼稻与热带粳的遗传距离更近[13].另一方面,籼稻和热带粳主要分布在亚洲热带地区,两者在地理条件、生态环境等方面非常相似,相同的生态适应性和种植区域的重叠可能增加了2 种类群之间的基因交流,导致类群间生殖隔离降低.按照籼粳亚种的分类方法,芳香稻DOM Sufid 属于粳稻,而在亚洲栽培稻5 种类群分类中,芳香稻属于单独一类,且与籼粳平行.本研究中DOM Sufid与籼、粳杂交均存在育性障碍,表明芳香稻可以作为一个独立的群体类型来进行研究,支持亚洲栽培稻5 种类群分类的方法.然而,因为只有1 个品种,代表性有限,后续可选用多个芳香稻品种来进行验证和进一步的研究.此外,Aus稻被认为是一类与籼、粳亚种杂交都表现出一定亲和力的品种,是亚洲栽培稻中除籼粳2 个亚种外还存在的中间类型[31].在本实验中,Aus稻无论是与滇粳优1 号还是IR64杂交,普遍存在育性障碍,进一步表明Aus稻应该作为独立于籼粳亚种外的生态类型来进行研究.由此看来,亚洲栽培稻较难仅从育性上简单地分为籼稻和粳稻亚种,本实验基于不同类群之间的杂种育性程度,更支持把亚洲栽培稻划分为5 种不同生态类型.
亚洲栽培稻亚种间杂种优势的利用是超高产育种的重要途径之一,但杂种不育现象严重阻碍了亚种间杂种优势的利用,因此认识杂种不育的遗传基础是利用杂种优势、培育超高产品种的前提.目前已经定位和克隆了多个控制亚洲栽培稻亚种间的育性基因,但这些研究没有针对亚洲栽培稻丰富的变异来进行,仅仅局限于籼粳亚种间,且使用的材料多为温带粳稻和籼稻,例如S5、S16、S25(t)、S33(t)、HSA1等[20].本研究中,温带粳品种与籼稻IR64、籼稻品种与滇粳优1 号配组的F1代育性变化范围很大,从高不育到正常可育.不同组合间育性存在明显的差异,同一组合内对花粉和小穗育性的影响程度也不同,表明控制育性的基因存在差异,这与前人的研究结果一致.此外,在分析母本相同、父本不同及父本相同、母本不同2 种情况下,发现由于父本品种不同或母本背景不同而导致杂交种的育性差异,进一步表明不同杂交组合含有不同的杂种不育位点,且控制杂种不育的基因在数目、功能、作用方式等方面存在差异[32].热带粳与温带粳滇粳优1 号之间存在育性障碍,目前对这方面的研究很少,也没有相关的育性基因被报道,后续可选用多个热带粳和温带粳品种来进行验证和进一步的研究.芳香稻主要分布于印度和巴基斯坦,与籼稻、粳稻及Aus稻在基因组方面存在差异[33],本研究从育性方面一定程度上证实这一观点.Aus稻作为一个特殊的生态类型,具有较好的亲和力[34].本研究中,Aus稻品种与滇粳优1 号和IR64 之间普遍存在育性障碍,杂交F1组合育性变化范围大,小穗育性变动在20.00%~ 93.19%,花粉育性变动在19.12%~ 98.28%.目前已通过图位克隆法将在第7 染色体调控Aus稻和籼稻/粳稻之间的杂种雌配子育性S7位点克隆[26].推测除S7位点,还有其他主效基因或者微效QTL 控制Aus稻和籼稻/粳稻之间的杂种雌配子育性.除控制小穗育性的基因外,还有影响花粉育性的基因存在,目前对这一方面的研究很少.此外,Aus稻品种含有广亲和基因S5-n[22],而Dular 同时携带有S5-n、S7-n、S8-n、S9-n、S16-n、S17-n、S29-n、S31-n、S32-n等广亲和基因[35],因此与滇粳优1 号以及IR64 之间没有育性障碍.Rayada、N22、FR13A、Kasalath 与滇粳优1 号或(和)IR64 存在一定程度上的育性障碍.上述现象表明这4 个Aus稻品种与籼稻/粳稻的育性除了被S5-n控制外,应该还存在其它位点控制育性.杂种优势难以被充分利用的主要原因之一是由于杂种结实率低[34].本实验中,有一些材料像籼稻Pokkali、籼稻Swarna、Aus稻Kasalath 分别与滇粳优1 号配组,热带粳Moroberekan、Aus稻Rayada 分别与IR64 配组等,花粉育性表现不正常,而小穗育性育性正常,这些材料可以解决由于结实率低导致杂种优势难以得到充分利用的问题.其中,籼稻Pokkali和热带粳Moroberekan 无论是与滇粳优1 号还是IR64 进行配组,均表现为花粉育性半不育,小穗完全可育,推测可能是由一个控制花粉育性的主效基因在起作用,需要进一步去研究.
本实验为系统研究亚洲栽培稻不同生态类型与籼、粳间杂种不育提供理论依据,为打破亚洲栽培稻亚种间育性障碍、充分利用杂种优势培育超高产品种提供材料和技术支撑,为利用亚洲栽培稻种内丰富的遗传基础培育杂交品种及指导不同背景(籼稻或粳稻)下杂交稻的选育工作提供理论指导,同时也为认识亚洲栽培稻的分化提供证据.5种生态类型在籼、粳背景下的育性研究一定程度上对亚洲栽培稻5 种生态类型的分类系统起到支撑作用,也存在一定的局限性.下一步,拟对5 种不同生态型材料分别在籼粳背景下杂种不育遗传和基因定位做系统性研究.研究结果将对全面地阐述亚洲栽培稻的杂种不育,为利用杂种优势提供理论指导和材料.
